Role of Endogenous Hormones in Germination and Dormancy and Gene Action on Hormones: A Comprehensive Review
Germination and dormancy are pivotal transitions in the seed life cycle, profoundly influenced by endogenous hormones. This review delves into the roles of major phytohormones, including abscisic acid (ABA), gibberellins (GAs), ethylene, Cytokinins, auxins, Jasmonates, and brassinosteroids (BRs) which elucidate their specific functions in promoting germination or maintaining dormancy, emphasizing the underlying molecular mechanisms. The study underscores the genetic underpinning of hormonal action, spotlighting key regulatory genes and their associated pathways. The intricate crosstalk between these hormones is explored, highlighting their collective impact on seed behaviours. Emerging perspectives encompass the role of hormones in priming treatments, stress responses, and seed longevity. Recent advances in genomic and epigenomic studies provide insights into complex feedback loops and the integration of environmental signals. Leveraging technologies like CRISPR/Cas9 has deepened our understanding of genetic influences on hormonal pathways. Overall, this comprehensive review illuminates the interplay between endogenous hormones and seed processes, offering potential strategies for improving germination efficiency and manipulating dormancy.
Introduction
Germination and dormancy are crucial fundamental stages in the life cycle of seeds, playing pivotal roles in plant propagation, survival, and population dynamics allowing them to colonize new habitats and ensure their survival under adverse conditions. The precise regulation of these processes is essential for successful seedling establishment and optimal plant growth. Endogenous hormones have emerged as key players in controlling germination and dormancy transitions, orchestrating a wide range of physiological and molecular events. Understanding the intricate interplay between endogenous hormones and seed behavior is of utmost importance in advancing our knowledge of plant development and optimizing agricultural practices.
Germination is the process by which a dormant seed resumes growth and develops into a seedling. It involves a series of coordinated physiological and biochemical changes, including water uptake, activation of metabolic pathways, cell expansion, and organ development. In contrast, seed dormancy refers to a state in which seeds fail to germinate even under favourable conditions.
Dormancy is a protective mechanism that enables seeds to withstand adverse environmental conditions, such as drought, cold, or darkness, until suitable conditions for growth and survival are encountered. Seed dormancy is an adaptive trait that evolved to ensure the germination of seeds occurs under optimal conditions for survival and growth. Dormancy can be influenced by various factors, including genetic, environmental, and hormonal cues. In particular, endogenous hormones play a significant role in mediating the balance between dormancy induction and dormancy release.
Phytohormones, also known as plant hormones, are small organic molecules that act as signalling molecules in plant growth and development. They exert profound effects on various aspects of plant physiology, including germination and dormancy regulation. Among the major classes of endogenous hormones involved in these processes are abscisic acid (ABA), gibberellins (GAs), ethylene, Cytokinins, auxins, Jasmonates, and brassinosteroids (BRs). These hormones exhibit distinct functions and interact with each other to finely modulate the balance between germination promotion and dormancy maintenance.
In recent years, advancements in molecular biology and genomics have provided novel insights into the complex hormonal networks and signalling pathways underlying germination and dormancy regulation. The identification of key genes involved in hormone biosynthesis, perception, and response has shed light on the genetic control of hormonal pathways in seeds. Additionally, emerging research has explored the role of hormones in seed priming, stress responses, seed longevity, and the manipulation of dormancy states for agricultural purposes.
This review aims to comprehensively examine the role of endogenous hormones in germination and dormancy, elucidating the molecular mechanisms and hormonal crosstalk involved. By synthesizing existing literature and incorporating emerging perspectives, we seek to provide a comprehensive overview of this vital area of research. Ultimately, a deeper understanding of the role of endogenous hormones in germination and dormancy will not only expand our fundamental knowledge of plant physiology but also hold great potential for improving agricultural practices and seed-based industries.
Background
Abscisic acid (ABA), a key hormone involved in stress responses, has long been recognized as a major player in inducing and maintaining seed dormancy. ABA accumulates in dormant seeds and inhibits germination by blocking cell growth and suppressing the expression of genes required for germination processes. Conversely, gibberellins (GAs) act as antagonists of ABA, promoting germination by stimulating cell expansion and activating gene expression [1]. The interplay between ABA and GAs, known as the ABA- GA balance, has been widely studied as a central regulator of dormancy and germination transitions [2]. In addition to ABA and GAs, other endogenous hormones such as ethylene, Cytokinins, auxins, Jasmonates, and brassinosteroids (BRs) have emerged as important modulators of germination and dormancy. Ethylene, for instance, can promote or inhibit germination depending on the concentration and environmental context [3]. Cytokinins and auxins play critical roles in cell division and elongation, contributing to seed germination and early seedling growth [4]. Jasmonates have been implicated in stress responses and defence mechanisms, affecting seed dormancy and germination [5]. The hormonal pathway directly regulates the abscisic acid which provides signals during germination and during the elongation and development of the plants [6]. BRs, on the other hand, are involved in cell elongation and division hypocotyl elongation, and their application can overcome dormancy and enhance germination efficiency [7].
Understanding the intricate interplay between these endogenous hormones is crucial for unravelling the regulatory networks that govern germination and dormancy. Recent advancements in molecular biology and omics technologies have provided new insights into hormone biosynthesis, signalling pathways, and gene regulation during seed development and maturation. Furthermore, the identification of hormone-related mutants and the exploration of hormone-responsive genes have deepened our understanding of the genetic control underlying germination and dormancy. Given the importance of germination and dormancy in plant propagation, agriculture, and ecological processes, a comprehensive review of the role of endogenous hormones in regulating these processes is highly valuable. By synthesizing the current knowledge and exploring emerging perspectives, this review aims to shed light on the multifaceted interactions between endogenous hormones and seed behaviour. Such insights have the potential to contribute to the development of innovative strategies for enhancing seed germination, overcoming dormancy limitations, and optimizing crop production in various agricultural and ecological contexts.
Novelty
This review article on the role of endogenous hormones in germination and dormancy presents a comprehensive synthesis of the latest research and emerging perspectives in the field. While previous reviews have explored the individual functions of specific hormones in germination and dormancy, this review goes beyond by highlighting the intricate interplay between multiple endogenous hormones and their collective impact on seed behaviour.
Furthermore, this review delves into novel aspects of hormone regulation in germination and dormancy. It explores the influence of endogenous hormones on seed responses to environmental cues, such as temperature, light, and moisture, shedding light on how hormonal signalling pathways integrate external signals to fine-tune germination and dormancy transitions. Additionally, this review investigates the role of endogenous hormones in seed priming and pre-sowing treatments, stress-induced germination and dormancy breakage, seed longevity and viability, and the genetic control of hormonal pathways.
By addressing these novel aspects, this review extends the current understanding of the role of endogenous hormones in germination and dormancy. It offers fresh insights into the complex hormonal networks that govern seed behaviour, providing a comprehensive and up-to-date overview of this critical field of research. The integration of recent advancements and emerging perspectives makes this review a valuable resource for researchers, agronomists, and plant biologists seeking to deepen their understanding of the multifaceted interactions between endogenous hormones and seed physiology.
Seeds are remarkable structures that contain the potential for plant growth and development. The induction and release of seed dormancy are influenced by a variety of factors, including genetic traits, environmental signals, and hormonal regulation. Endogenous hormones play a crucial role in mediating the balance between dormancy and germination. Hormones such as abscisic acid (ABA) are known to promote dormancy, while others like gibberellins (GAs) are involved in stimulating germination [8]. Understanding the processes of germination and dormancy is essential for comprehending the intricate role of endogenous hormones in regulating these stages. This section provides a concise introduction to the mechanisms underlying germination and dormancy, setting the stage for a deeper exploration of the specific role of endogenous hormones in these processes.
Key Factors Influencing the Transition Between Germination and Dormancy States
The transition between germination and dormancy in seeds is regulated by a complex interplay of various factors. Understanding these key factors is essential for unraveling the mechanisms that control the switch between these states. This section explores the factors that influence the transition between germination and dormancy in seeds, shedding light on the intricate nature of seed behavior and development.
Environmental Cues
Environmental factors such as temperature, light, moisture, and oxygen availability play crucial roles in determining the transition between germination and dormancy. Optimal conditions, including specific temperature ranges and light qualities, can trigger germination, while unfavourable conditions can maintain or induce dormancy. Temperature play vital role in regulation of abscisic aid and Gibberellins. Seeds like Arodiopsis require a period of cold to break dormancy. Cold stratification period decreases ABA levels and/or sensitivity, thus promoting germination [9]. Cold can induce the expression of genes that degrade ABA, like the CYP707A family [10]. Two genes PpCYP707As and PpNCEDs found in peach which regulates seed and bud dormancy is influenced by exposure of cold temperature [11]. Similarly extreme temperature might create condition like Thermoinhibition and Thermodormancy. High temperatures can inhibit germination in some seeds, in part through the suppression of GA biosynthesis or signalling [12]. In certain seeds, high or low temperatures can induce a secondary dormancy, often modulated by Gas [13]. Similarly, light also plays pivotal role in phytochromes and ABA. Seeds exposed to two forms of phytochromes Pr (inactive, red light-absorbing) and Pfr (active, far-red light absorbing) can promote or inhibit the plant growth regulators. Seeds in the soil are exposed to a high red to far-red ratio, converting Pr to Pfr. The active Pfr form can promote GA synthesis and inhibit ABA synthesis, promoting germination [14]. Light can reduce ABA concentrations, promoting the germination. The study shows that exposure to light can induce CYP707A expression, leading to ABA degradation [15]. Moisture play an indispensable role for seed imbibition which is the initial step for germination. Dry conditions or water stress can elevate ABA synthesis, inhibiting germination. When seeds sense adequate moisture, ABA levels drop, and the germination process can begin [16]. In waterlogged conditions, ethylene synthesis is promoted. Ethylene acclimate by promoting lysigenous aerenchyma formation [17]. Ethylene can override ABA’s inhibitory effect and promote germination, especially in species adapted to such conditions [3]. Hormonal Balance Endogenous hormones, particularly abscisic acid (ABA) and gibberellins (GAs), are central regulators of the germination-dormancy transition. The balance between these hormones, known as the ABA-GA balance, determines the dominance of dormancy or germination. A high ABA level suppresses germination, while an increase in GA promotes germination [2].
Genetic Factors
Genetic traits and variations among seed populations influence the transition between germination and dormancy. Different plant species or even different accessions within a species may exhibit varying degrees of seed dormancy due to genetic differences.
Endogenous Hormones and Their Role in Plant Development
Endogenous hormones, also known as plant hormones or phytohormones, are natural signalling molecules that regulate various aspects of plant growth, development, and physiological responses. These hormones are produced within the plant, acting as chemical messengers, coordinating and modulating a wide range of cellular processes. Endogenous hormones play pivotal roles in plant development from seed germination to flowering, fruiting, and senescence. They regulate processes such as cell division, elongation, differentiation, and organ formation, ensuring the proper growth and morphogenesis of plants. The major classes of endogenous hormones include abscisic acid (ABA), auxins, Cytokinins, gibberellins (GAs), ethylene, Jasmonates, and brassinosteroids (BRs). Each hormone has specific functions and interacts with others to form complex signalling networks.
ABA
ABA is primarily involved in stress responses, regulating seed dormancy, stomatal closure, and the adaptation of plants to environmental challenges such as drought and salinity [18]. Auxins Auxins control cell elongation, tissue differentiation, and apical dominance. They are responsible for tropic responses such as phototropism and gravitropism [19].
Cytokinins
Cytokinins promote cell division and affect shoot and root development. They interact with auxins to maintain the balance between shoot and root growth [20]. GAs GAs are essential for stem elongation, seed germination, and the transition from the vegetative to the reproductive phase. They counteract the inhibitory effects of ABA on seed germination [21]. Ethylene Ethylene regulates fruit ripening, senescence, and the response to biotic and abiotic stresses. It also plays a role in seed germination, root development, and organ senescence [22]. Jasmonates Jasmonates are involved in defence responses against herbivores and pathogens, as well as in regulating plant growth and development, including root growth and flower formation [23]. BRs BRs promote cell elongation, stimulate vascular tissue differentiation, and affect leaf and flower development. They also play a role in seed germination and responses to environmental stresses [24].
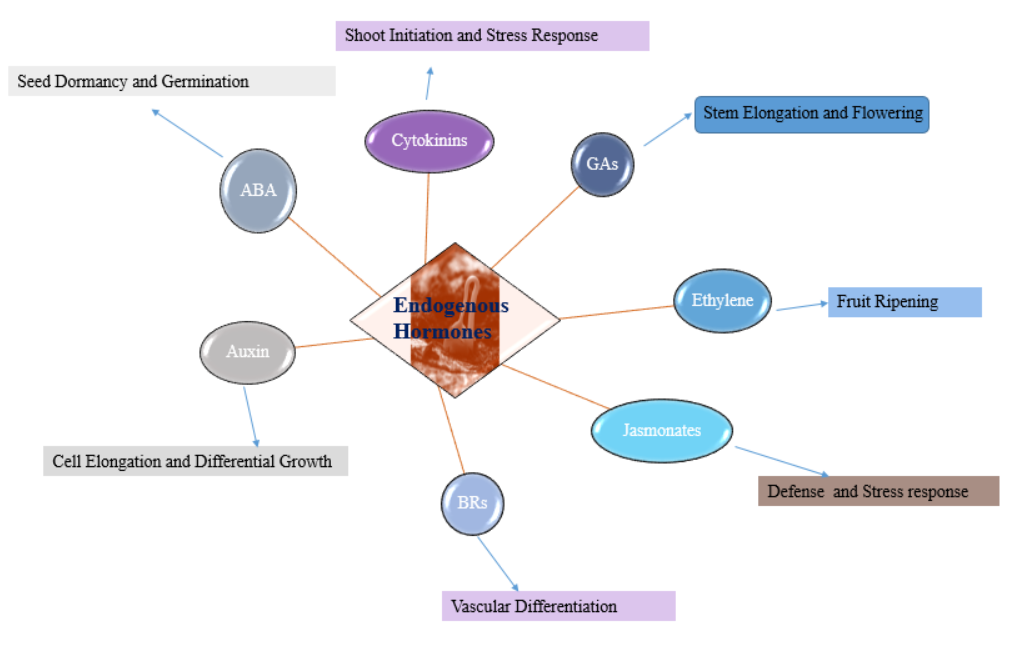
The intricate interactions and crosstalk among these endogenous hormones allow plants to integrate internal and external cues, fine-tuning their growth and development to optimize survival and reproductive success.
Understanding the roles and mechanisms of action of endogenous hormones is crucial for unravelling the complexities of plant development. This knowledge not only enhances our understanding of fundamental plant biology but also has practical applications in agriculture, horticulture, and plant biotechnology, where manipulating hormone levels or responses can lead to improvements in crop yield, stress tolerance, and overall plant performance.
Gene Action of hormones
| Gene | Typical function | |
|---|---|---|
| NCED (9-cis- epoxycarotenoid dioxygenase) | Increased ABA levels. | ABA biosynthesis |
| CYP707A | ABA levels in the seed decrease | Promoting germination. |
| PYR/PYL/RCAR | When ABA is present, leading to dormancy. | Initiate signalling pathways |
Table 2: Gene associated with Abscisic Acid (ABA).
| Gene | Typical function |
|---|---|
| GA3ox, GA20ox | When they are active, they lead to the production of biologically active GAs that promote seed germination |
| GA2ox | Deactivation of GAs that promote dormancy |
Table 3: Gene associated with Gibberellins (GAs).
| Gene | Typical function |
|---|---|
| ACS (1-aminocyclopropane- 1-carboxylate synthase) | Expression results in increased ethylene production |
| ACO (1-aminocyclopropane- 1-carboxylate oxidase): | Increased ethylene production. |
Table 4: Gene associated with Ethylene.
| Gene | Typical function |
|---|---|
| DWF4 | BR biosynthesis |
| IPT (Isopentenyl transferase): | When expressed, it leads to increased cytokinin levels. |
Table 5: Gene associated with Brassinosteroids (BRs) and Cytokinins.
| Gene | Typical function |
|---|---|
| YUCCA | When they are active, they lead to increased auxin production. |
| TIR1/ AFB | Auxin signalling. |
Table 1: Gene associated with Auxins.
Integration of Hormonal Signalling Networks in Germination and Dormancy
BRs and Gibberellins (GAs)
BRs and GAs exhibit both synergistic and antagonistic interactions in seed physiology. While GAs are generally associated with promoting germination, BRs can enhance GA responses and synergistically regulate germination processes. Conversely, BRs can also act antagonistically to GAs, inhibiting GA-induced processes and maintaining dormancy.
BRs and Abscisic Acid (ABA)
The interaction between BRs and ABA is crucial for balancing seed dormancy and germination. ABA promotes seed dormancy, while BRs counteract the inhibitory effects of ABA and promote germination. BRs can enhance the degradation of ABA or interfere with ABA signalling pathways, resulting in the inhibition of dormancy and the promotion of germination.
BRs and Cytokinins
BRs and Cytokinins often act synergistically in promoting seed germination. Cytokinins, known for their role in cell division and growth, can enhance BR-induced processes such as cell elongation and differentiation, leading to seed germination.
BRs and Ethylene
BRs and ethylene exhibit complex interactions in seed physiology. While ethylene can induce seed dormancy and inhibit germination, BRs can counteract the inhibitory effects of ethylene and promote germination processes.
BRs and Auxins
BRs and auxins interact synergistically in regulating seed germination and growth. Auxins are involved in various aspects of seed development, including embryo growth and root elongation, while BRs enhance auxin responses and promote these processes.
Understanding the dynamic interactions between BRs and other hormones in seed physiology provides insights into the complex regulatory networks that govern seed germination, dormancy, and growth. Unravelling the molecular mechanisms and cross-regulation among these hormones opens up possibilities for manipulating seed behaviour, optimizing seedling establishment, and improving crop productivity in agriculture.
Unravelling the Genetic Control of Hormonal Pathways in Germination and Dormancy
| SN | Gene/Protein | Signalling and Pathways | Function | References |
|---|---|---|---|---|
| 1 | CORONITINE INSENSITIVE 1 (COI1) | Leucine-rich repeat containing F-box protein that recognizes and interacts with JAZ proteins Along with S-PHASE KINASE-ASSOCIATED PROTEIN1, CULLIN, and RING-BOX PROTEIN1, COI1 forms a complex | Acts as a receptor for plant hormones named as auxin Polyubiquitinates JAZ proteins, marking them for degradation via the 26S proteasome | (Thines, et al. [25] and Chini, et al. [26]) |
| 2 | JASMONATE ZIM- DOMAIN (JAZ) | AZ represses activity of the bHLH factors belonging to sub-group IIIb (ICE1 and ICE2), IIIe (MYC2, MYC3, and MYC4) and IIIf (GL3, EGL3 and TT8), the AP2 TF TARGET OF EAT1 and 2 (TOE1 and TOE2), as well as the MYB factors PAP1, GL1 MYB21, and MYB2 JAZ interacts with NINJA via the TIFY domain Recruits the corepressor TOPLESS (TPL In the presence of JA, COI1 and Degradation of JAZ frees MYC2 and other TFs to activate downstream JA responsive genes | Control plant defence against the attack of herbivore, regulate various aspect of development of plants including tendril coiling, ripening of the fruits and senescence | (Niu, et al. [27] Zhai, et al. [28] and Pauwels, et al. [29]) |
| 3 | MYC2 (bHLH transcription factor (TF)) | Master regulators activating JA responsive genes | Plays important roles in stress tolerance of plants, biosynthesis of secondary metabolites such as anthocyanin, flower and fruit development. | (Dombrecht, et al. [30]) |
| 4 | TOPLESS | TPL recruits histone deacetylases to modify chromatin architecture to repress target gene transcription | Helps in seed maturation and flower development | (Krogan, et al. [31]) |
| 5 | MEDIATOR SUBUNIT 2 | Functions as both a coactivator and corepressor and recruits RNA polymerase II to the promoter of activated genes | Transcriptional Regulation | (Flanagan, et al. [32] and Kelleher, et al. [33]) |
| 6 | JA-isoleucine (Conjugated signalling molecule) | Activates the MYC transcription factors by directly Binding to JAZ and COI1 | Pollen maturation | (Ruan, et al. [34]) |
| 7 | Arabidopsis JASMONATE TRANSPORTER (AtJAT1) | Facilitate the export of JA across the plasma membrane | Increase plant resilience Promote adventitious root regeneration | (Li Q, et al. [35]) |
| 8 | Systemin | Transported to other cells via the apoplast and combined with the cell surface receptor SR160 (a protein rich in leucine repeat units)to finally activate the JA signalling pathway | Modify root growth | (Li C, et al. [36]) |
| 9 | 26S proteasome | JAZ proteins are degraded after being transferred to the 26S proteasome | Defense against herbivores and pathogens, as well as in growth and development. | (Ruan, et al. [34]) |
| 10 | NAC (ATAF1 and ATAF2) | NAC family are both induced by JA signalling | Signal transduction of ABA hormones Involved in plant resistance to drought, salt stress, Botrytis cinerea, and other pathogens | (Delessert, et al. [37]) |
| 11 | WRKY | In addition , Arabidopsis WRKY57 interacts with the inhibitor JAZ4/ JAZ8 in the JA signalling pathway and the inhibitor IAA29 in the auxin signalling pathway, thereby regulating the interaction between JA and auxin- mediated signalling pathways | Regulate plant hormone signal transduction pathway. Effects on plant leaf senescence Increase the levels of JA and JA-Ile. In Nicotiana attenuata, regulate the expression of JAs biosynthesis-related genes (LOX,AOS,AOC, and OPR) | (Skibbe, et al. [38] Jiang, et al. [39]) |
| 12 | ERF | Regulates the biosynthesis of hydroxycinnamic acid amides | Regulate Hormone ABA | (Sasaki-Sekimoto, et al. [40]) |
| 13 | Jam Proteins - JASMONATE- ASSOCIATED MYC2- LIKE1 (JAM1, JAM2, JAM3) | Transcriptional repressors via forming protein–protein interactions with JAZs to regulate JAs responses | Pollen Maturation | (Li Q, et al. [35]) |
| 14 | mitogen-activated protein kinase (WIPK) | Expression of WIPK | Wound-induced JA biosynthesis. | (Seo S, et al. [41]) |
| 15 | OPR gene | Encodes OPR enzyme Catalyze conversion from OPDA to 12-oxo- phylodienoic acid | Growth and development of plants and hormonal regulation | (Turner, et al. [42] and He, et al. [43]) |
| 16 | JAR (auxin induced gene) | Catalyze formation of biologically active jasmonyl-isoleucine (JA-lle) conjugate Promotes the interaction between JAZ1 and COI1 in jasmonate signalling pathways | Root growth | (Ruan, et al. [34]) |
| 17 | JIN1 | encodes AtMYC2, whose expression is rapidly upregulated by JA repressed by AtMYC2 Systemic response to wounding activated by AtMYC2 | Pollen maturation Defense response against pathogen | (Ruan, et al. [34]) |
| 18 | JMT | Transgenic Arabidopsis overexpressing JMT accumulates MeJA without altering Jasmonic acid content, expresses the JA-responsive genes for VSP and PDF1.2 Expression of JMT alone is sufficient to induce some JA-dependent responses | Vegetative growth and development of plants Developing flowers Displays enhanced resistance to infection by Botrytis cinerea MeJA can function as an endogenous signal molecule in plant defences. | (Seo HS, et al. [44] and Li C, et al. [23]) |
| 19 | JAR | Rather than acting as a signalling intermediate, it biochemically modify JA itself. | Flower maturation Affect jasmonate signal transduction Reduce sensitivity to JA | (Ruan, et al. [34]) |
| 20 | PR genes | Act as OPDA Reductase | Plant defence against pathogens | (Wang G, et al. [45]) |
| 21 | VSP2 genes | Work as a vegetative storage protein 2 | Control leaf and flower morphology | (Wang G, et al. [45]) |
| 22 | DAD1 | DAD1 possesses a chloroplast transit peptide at its N-terminus and can accumulate in chloroplasts. | Pollen maturation Filament development, pollen grain formation, and anther dehiscence. JA biosynthesis in stamen formation, | (He, et al. [43]) |
| 23 | LOX2 | LOX2 is required for wound-induced JA formation, LOX2 genes is required for JA formation in pollen and stamen development. LOX2 gene transcripts accumulate in response to JA | JA-dependent pollen and stamen development. | (Turner, et al. [42]) |
| 24 | AOC | AOC transcripts are transiently induced in wounded tomato leaves, where it is expressed primarily in the vascular bundle tissues | Biosynthesis of Jasmonic acid (JA), The AOC mRNA is expressed at low levels in stems, young leaves, and young flowers, contrasting with a high accumulation in flower buds, flower stalks, and roots. | (Turner, et al. [42]) |
| 25 | FAD Fatty acids desturase | Modifying fatty acids bound to coenzyme A or embedded in membrane lipids, such as phosphatidylcholine. | Biosynthesis of polyunsaturated fatty acids | (Wang G, et al. [45]) |
| 26 | AOS (Allene oxide synthase) | Conversion of α-linolenic acid from membrane lipids into JA. | Biosynthesis of Jasmonic acid (JA | (Wang G, et al. [45]) |
| 27 | LEC | Lectine family protein LEC genes, primarily LEC1, LEC2, and FUSCA3 (FUS3), are transcription factors regulating developmental processes | Helps in development of embryo and maturation | (Wang G, et al. [45]) |
| 28 | PLD (in Arabidopsis) | Phospholipase than can be induced by systemin and AtPEP1 | Root hair patterning, root growth | (Ruan, et al. [34]) |
| 29 | DGL (in Arabidopsis) | Induced by systemin and AtPEP1 | Control flowering time, morphology of flowers | (Ruan, et al. [34]) |
Table 6: Analysis of the genetic factors governing hormone biosynthesis, signalling, and response pathways in seeds.
Conclusion
In conclusion, this comprehensive review has highlighted the crucial role of endogenous hormones in the regulation of germination and dormancy in seeds. The interplay between hormones such as abscisic acid (ABA), gibberellins (GAs), ethylene, Cytokinins, auxins, Jasmonates, and brassinosteroids (BRs) orchestrates the balance between dormancy induction and germination promotion. Through their intricate signalling pathways, these hormones integrate genetic, environmental, and physiological cues to fine-tune seed behaviour.
The review has provided an overview of the individual functions of these hormones, elucidating their roles in promoting germination or maintaining dormancy. The molecular mechanisms underlying hormone actions, including gene regulation, cellular processes, and hormone crosstalk, have been explored. Furthermore, the review has highlighted novel perspectives, such as the influence of hormones in seed responses to environmental cues, seed priming and pre-sowing treatments, stress-induced responses, seed longevity, and the genetic control of hormonal pathways.
The findings presented in this review contribute to a deeper understanding of the complex hormonal networks that regulate germination and dormancy. Such knowledge holds great potential for practical applications in agriculture, horticulture, and ecological restoration. Manipulating endogenous hormone levels or their signalling pathways could enhance seed germination rates, overcome dormancy limitations, and optimize crop production in various environmental conditions.
Despite the significant progress made in unravelling the role of endogenous hormones, there are still gaps in our understanding. Further research is needed to explore the cross-talk and synergy among different hormones, as well as their interactions with other regulatory factors. Additionally, the genetic control of hormonal pathways and the identification of key genes involved in hormone biosynthesis, perception, and response should be further investigated.
In conclusion, the role of endogenous hormones in germination and dormancy represents a fascinating and complex area of study. This review provides a comprehensive overview of the current knowledge, highlights emerging perspectives, and identifies avenues for future research. By advancing our understanding of the intricate hormonal regulation in seeds, we can unlock new possibilities for improving seed-based industries, ecological restoration efforts, and sustainable agriculture practices.
Conflict of Interest
The author affirmed no conflict of interest.
Acknowledgement
The authors deliver gigantic gratitude towards the helping hands including friends, family members, and participants throughout the period of study.
References
-
Liu X, Hou X (2018) Antagonistic Regulation of ABA and GA in Metabolism and Signaling Pathways. Front Plant Sci 9: 251.
-
Shu K, Liu X, Xie Q, He Z (2016) Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol Plant 9(1): 34-45.
-
Corbineau F, Xia Q, Bailly C, El-Maarouf-Bouteau H (2014) Ethylene, a key factor in the regulation of seed dormancy. Front Plant Sci 5: 539.
-
Sosnowski J, Truba M, Vasileva V (2023) The Impact of Auxin and Cytokinin on the Growth and Development of Selected Crops. Agriculture 13(3): 724.
-
Xu Q, Truong TT, Barrero JM, Jacobsen JV, Hocart CH, et al. (2016) A role for jasmonates in the release of dormancy by cold stratification in wheat. J Exp Bot 67(11): 3497- 3508.
-
Pan J, Hu Y, Wang H, Guo Q, Chen Y, et al. (2020) Molecular Mechanism Underlying the Synergetic Effect of Jasmonate on Abscisic Acid Signaling during Seed Germination in Arabidopsis. The Plant Cell 32(12): 3846-3865.
-
Castro-Camba R, Sánchez C, Vidal N, Vielba JM (2022) Plant Development and Crop Yield: The Role of Gibberellins. Plants 11(19): 2650.
-
Liu X, Zhang H, Zhao Y, Feng Z, Li Q, et al. (2013) Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proceedings of the National Academy of Sciences 110(38): 15485-15490.
-
Rodríguez-Gacio MDC, Matilla-Vázquez MA, Matilla AJ (2009) Seed dormancy and ABA signaling. Plant Signal Behav 4(11): 1035-1048.
-
Dejonghe W, Okamoto M, Cutler SR (2018) Small Molecule Probes of ABA Biosynthesis and Signaling. Plant Cell Physiol 59(8): 1490-1499.
-
Wang D, Gao Z, Du P, Xiao W, Tan Q, et al. (2016) Expression of ABA Metabolism-Related Genes Suggests Similarities and Differences Between Seed Dormancy and Bud Dormancy of Peach (Prunus persica). Front Plant Sci 6: 1248.
-
Toh S, Imamura A, Watanabe A, Nakabayashi K, Okamoto M, et al. (2008) High Temperature-Induced Abscisic Acid Biosynthesis and Its Role in the Inhibition of Gibberellin Action in Arabidopsis Seeds. Plant Physiol 146(3): 1368- 1385.
-
Martel C, Blair LK, Donohue K (2018) PHYD prevents secondary dormancy establishment of seeds exposed to high temperature and is associated with lower PIL5 accumulation. J Exp Bot 69(12): 3157-3169.
-
Farooq MA, Ma W, Shen S, Gu A (2022) Underlying Biochemical and Molecular Mechanisms for Seed Germination. Int J Mol Sci 23(15): 8502.
-
Okamoto M, Kuwahara A, Seo M, Kushiro T, Asami T, et al. (2006) CYP707A1 and CYP707A2, which Encode Abscisic Acid 8′-Hydroxylases, Are Indispensable for Proper Control of Seed Dormancy and Germination in Arabidopsis. Plant Physiology 141(1): 97-107.
-
Muhammad Aslam M, Waseem M, Jakada BH, Okal EJ, Lei Z, et al. (2022) Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. International Journal of Molecular Sciences 23(3): 1084.
-
Ali S, Kim WC (2018) Plant Growth Promotion Under Water: Decrease of Waterlogging-Induced ACC and Ethylene Levels by ACC Deaminase-Producing Bacteria. Front Microbiol 9: 1096.
-
Xiong L, Zhu JK (2003) Regulation of Abscisic Acid Biosynthesis. Plant Physiol 133(1): 29-36.
-
Liscum E, Askinosie SK, Leuchtman DL, Morrow J, Willenburg KT, et al. (2014) Phototropism: Growing towards an Understanding of Plant Movement. Plant Cell 26(1): 38-55.
-
Su YH, Liu YB, Zhang XS (2011) Auxin–Cytokinin Interaction Regulates Meristem Development. Molecular Plant 4(4): 616-625.
-
Finkelstein RR, Gampala SSL, Rock CD (2002) Abscisic Acid Signaling in Seeds and Seedlings. Plant Cell 14(Suppl): s15-45.
-
Lin Z, Zhong S, Grierson D (2009) Recent advances in ethylene research. J Exp Bot 60(12): 3311–3336.
-
Li C, Xu M, Cai X, Han Z, Si J, et al. (2022) Jasmonate Signaling Pathway Modulates Plant Defense, Growth, and Their Trade-Offs. Int J Mol Sci 23(7): 3945.
-
Vriet C, Russinova E, Reuzeau C (2012) Boosting Crop Yields with Plant Steroids. Plant Cell 24(3), 842-857.
-
Thines B, Katsir L, Melotto M, Niu Y, Mandaokar A, et al. (2007). JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 448(7154): 661-665.
-
Chini A, Fonseca S, Fernández G, Adie B, Chico JM, et al. (2007) The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448(7154): 666-671.
-
Niu Y, Figueroa P, Browse J (2011) Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis. J Exp Bot 62(6): 2143-2154.
-
Zhai Q, Zhang X, Wu F, Feng H, Deng L, et al. (2015) Transcriptional Mechanism of Jasmonate Receptor COI1-Mediated Delay of Flowering Time in Arabidopsis. Plant Cell 27(10): 2814-2828.
-
Pauwels L, Barbero GF, Geerinck J, Tilleman S, Grunewald W, et al. (2010) NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature, 464: 788-791.
-
Dombrecht B, Xue GP, Sprague SJ, Kirkegaard JA, Ross JJ, et al. (2007) MYC2 Differentially Modulates Diverse Jasmonate-Dependent Functions in Arabidopsis. Plant Cell 19(7): 2225-2245.
-
Krogan NT, Hogan K, Long JA (2012) APETALA2 negatively regulates multiple floral organ identity genes in Arabidopsis by recruiting the co-repressor TOPLESS and the histone deacetylase HDA19. Development 139(22): 4180-4190.
-
Flanagan PM, Kelleher RJ, Sayre MH, Tschochner H, Kornberg RD (1991) A mediator required for activation of RNA polymerase II transcription in vitro. Nature 350(6317): 436-438.
-
Kelleher RJ, Flanagan PM, Kornberg RD (1990) A novel mediator between activator proteins and the RNA polymerase II transcription apparatus. Cell 61(7): 1209- 1215.
-
Ruan J, Zhou Y, Zhou, M, Yan J, Khurshid M, et al. (2019) Jasmonic Acid Signaling Pathway in Plants. Int J Mol Sci 20(10): 2479.
-
Li Q, Zheng J, Li S, Huang G, Skilling SJ, et al. (2017) Transporter-Mediated Nuclear Entry of Jasmonoyl- Isoleucine Is Essential for Jasmonate Signaling. Mol Plant 10(5): 695-708.
-
Li C, Liu G, Xu C, Lee GI, Bauer P, et al. (2003). The Tomato Suppressor of prosystemin-mediated responses2 Gene Encodes a Fatty Acid Desaturase required for the Biosynthesis of Jasmonic Acid and the Production of a Systemic Wound Signal for Defense Gene Expression. Plant Cell 15(7): 1646-1661.
-
Delessert C, Kazan K, Wilson IW, Straeten DVD, Manners J, et al. (2005) The transcription factor ATAF2 represses the expression of pathogenesis-related genes in Arabidopsis. Plant J 43(5): 745-757.
-
Skibbe M, Qu N, Galis I, Baldwin IT (2008) Induced Plant Defenses in the Natural Environment: Nicotiana attenuata WRKY3 and WRKY6 Coordinate Responses to Herbivory. Plant Cell 20(7): 1984-2000.
-
Jiang, Y, Liang G, Yang S, Yu D (2014) Arabidopsis WRKY57 Functions as a Node of Convergence for Jasmonic Acid– and Auxin-Mediated Signaling in Jasmonic Acid–Induced Leaf Senescence. Plant Cell 26(1): 230-245.
-
Sasaki-Sekimoto Y, Jikumaru Y, Obayashi T, Saito H, Masuda S, et al, (2013) Basic Helix-Loop-Helix Transcription Factors JASMONATE-ASSOCIATED MYC2- LIKE1 (JAM1), JAM2, and JAM3 Are Negative Regulators of Jasmonate Responses in Arabidopsis. Plant Physiology 163(1): 291-304.
-
Seo S, Okamoto M, Seto H, Ishizuka K, Sano H, et al. (1995) Tobacco MAP Kinase: A Possible Mediator in Wound Signal Transduction Pathways. Science 270(5244): 1988-1992.
-
Turner JG, Ellis C, Devoto A (2002) The Jasmonate Signal Pathway. Plant Cell, 14(Suppl): s153-164.
-
He L, Chen X, Xu M, Liu T, Zhang T, et al. (2021) Genome- Wide Identification and Characterization of the Cystatin Gene Family in Bread Wheat (Triticum aestivum L.). Int J Mol Sci 22(19): 10264.
-
Seo HS, Song JT, Cheong JJ, Lee YH, Lee YW, et al. (2001) Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses. Proceedings of the National Academy of Sciences, 98(8): 4788-4793.
-
Wang G, Huang W, Li M, Xu Z, Wang F, et al. (2016) Expression profiles of genes involved in jasmonic acid biosynthesis and signaling during growth and development of carrot. Acta Biochim Biophys Sin 48(9): 795-803.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends