The Impact of Mitochondrial KATP Channel Opening on Energy Metabolism under Physical Stress
<p>The modulation of mKATP channel activity is known to have a great impact on energy metabolism in a living organism. The aim of this work was to study the impact of mKATP channels opening on the physical endurance in the rats subject to compulsory swimming with a load. Our purpose was to find a mechanistic basis to explain the modulation of the energy metabolism under exercise training by studying the direct effect of mKATP channel opening on mitochondrial functions. Male Wistar rats exhibiting high and low resistance to physical stress were separated in two groups, and subjected to compulsory swimming with a load. Swimming time (ST) was monitored from the start till the time when the rats began to drawn. ST was reliably higher in high resistance group, which coincided with higher endogenous mKATP channels activity. mKATP channels blockers, glibenclamide and 5-hydroxydecanoate shown to completely block mKATP channel in vivo, dramatically reduced ST in both groups, which indicated its dependence on mKATP channel activity. To find a mechanistic basis for the observed dependence, we studied the effect of mKATP channels opening on mitochondrial functions in vitro. mKATP channels opener diazoxide stimulated state 4 respiration and decreased RCR, but increased P/O ratio. On the contrary, mKATP channels blockers, dramatically reduced P/O. Based on the experiments, we came to the conclusion of the correlation between the physical endurance and P/O ratio, both dependent on mKATP channels activity. Thus, mKATP channels opening inhibited phosphorylation, but increased its efficiency. In vivo this resulted in the improvement of the endurance in the animals with elevated mKATP channel activity. Physiological relevance of the observed phenomena is discussed.</p>
Introduction
In agreement with the contemporary views, an mKATP channel functioning has a profound impact on the bioenergetics of a living organism. Under metabolic stress conditions, such as ischemia/reperfusion of myocardium mKATP channels were shown to be a promising molecular target of cardioprotective therapy [1, 2, 3]. Multiple experimental works and clinical studies established that administration of pharmacological mKATP channels openers was helpful for the effective recovery of cardiac tissues from ischemia and the risks of oxidative damadge after reperfusion. Similarly, it was shown the ability of mKATP channels openers to attenuate oxidative damage caused by the oxygen deprivation under ischemia and hypoxia [2, 3, 4, 5]. Potassium transport is often considered to be an all- round modulator of mitochondrial functions. It takes part in the regulation of mitochondrial volume, respiration, membrane potential, calcium transport, ROS production and ATP synthesis [6, 7, 8, 9]. Generally, it was supposed that the background of cardio- and neuroprotective effects of mKATP channels opening was the triggering of cellular ROS-dependent signaling and/or attenuation of ROS overproduction in mitochondria [1, 10, 11]. Mitochondrial permeability transition pore was one acknowledged molecular target of cardioprotective therapy using pharmacological mKATP channels openers. The inhibition of the pore activity following mKATP channels opening was shown to prevent the development of cellular apoptosis and necrosis under pathophysiological conditions [1, 2, 11]. Meanwhile, several published data, including the results of our work have shown that oxidative phosphorylation system is another important target of the pharmacological modulators of mKATP channels [12]. However, physiological meaning of the impact of mKATP channels opening on mitochondrial ATP synthesis is not quite well understood. Earlier it was observed that the animals’ resistance to the exposure to physical stress (swimming with a load) correlated with endogenous activity of mKATP channel [13]. Highly resistant animals (HR group) exhibited high mKATP channel activity, while the channel activity in low resistant group (LR group) was low. In our previous work we also tried to assess physiological relevance of the effect of mKATP channels opening on the oxidative phosphorylation by studying the impact of short-term acute hypoxia on the physical endurance of the animals subject to compulsory swimming with a load. By our observations, increase of mKATP channel activity after the short-term acute hypobaric hypoxia resulted in the improvement of the rats’ endurance to physical stress, which prompted further extension of our research [12]. So, the aim of this work was to find the role of mKATP channel in the regulation of energy metabolism in vivo by examining the correlation between animals’ endurance to exercise training and the endogenous mKATP channel activity.
Materials and Methods
The Animals
Adult Wistar-Kyoto male rats with body weight 180- 200 g were chosen for the experiments. Animals were processed according to the European Community Council Directive 86/609/EEC and approved by the Ethics Commission on Animal Experiments of A.A. Bogomoletz Institute of Physiology, NAS Ukraine. The rats were subjected to compulsory swimming with the load. The load weight was 20% of the body mass. The swimming bath was constructed in a way that animal could not reach the bottom of the vessel. Swimming continued until the animals were physically exhausted and began to drawn. Swimming time (ST) as the measure of physical endurance was registered. Using approach developed previously, the rats exhibiting high and low resistance to physical loading based on their ST (HR and LR groups) were separated from those showing intermediate results, and mKATP channels activity was assessed ex vivo on isolated liver mitochondria with K+-selective electrode as we described earlier [12, 13].
Mitochondria Isolation
Immediately after the experiment mitochondrial preparations were obtained according to following procedures. Liver was carefully washed by chilled 0.9% KCl and homogenized in a medium: 250 mM sucrose, 1 mM EDTA, 20 mM Tris-HCl buffer, 4°C (pH 7.2). Isolated mitochondria were obtained by differential centrifugation of homogenates at 700 x g 10 min, and then centrifugation of supernatant at 11000 x g 15 min (4°C). The residues were suspended in a small volume of isolation medium without EDTA. Protein concentration was determined by the method of Lowry. All preparations were stored on ice.
The Study of Oxygen Consumption
Oxygen consumption was studied polarographically in 1 cm3 closed termostated cell at 26°C with platinum electrode at constant stirring in standard incubation medium: 120 mM KCl, 0.5 mM EDTA, 5 mM sodium glutamate, 1 mM KH2PO4, 20 mM Tris-HCl buffer (pH 7.4). ADP was added at 0.2 mM. Respiratory control ratio RCR was found as the ratio of state 3 to state 4 rates of respiration _J_3/_J_4. Phosphorylation efficiency P/O was found as the ratio of ADP to the oxygen consumed in the course of phosphorylation. The rate of ATP synthesis was found from the time of ADP consumption. For studying mKATP channel activity, Mg2+ was added at 1 mM, ATP at 0.3 mM, and oligomycin at 1 μg/mg protein; final protein concentration was 1.5-2.0 mg/ml.
Assessment of mKATP Channel Activity with K+-Selective Electrode
mKATP channel activity was assessed by the method of Baranova, et al. under conditions of membrane depolarization [14]. Potassium efflux from mitochondria (1.5-2.0 mg protein) was stimulated by membrane depolarization using protonophore CCCP, and the increase in potassium concentration was registered at room temperature with K+-selective microelectrode in 1 cm3 cell in the incubation medium: 250 mM sucrose, 0.5 mM EGTA, 1 mM Mg2+, 1 mM NaH2PO4, 5 mM sodium glutamate, 20 mM Tris-HCl buffer (pH 7.4). Oligomycin was added to the medium at 1 μg/mg protein, CCCP was added at 1 μM. mKATP channel activity was estimated as ATP-dependent component of potassium flux, found as the difference between the initial transport rates in the absence and the presence of 1 mM ATP [14]. The rates of potassium transport were derived from the calibration curves obtained by the titration of depolarized mitochondria by the aliquots of KCl solution in the presence of 10−7 M valinomycin.
Chemicals
All reagents were from Sigma-Aldrich, USA. Deionized water was used for medium preparations.
Statistical Analysis
The data were expressed as mean ± S.E. of 4-6 independent experiments. Statistical analysis was performed using paired Student’s t-test; P<0.05 was taken as the level of significance.
Results
High resistant (HR) and low resistant (LR) animals were separated as described in Methods section based on their endurance to physical loading, which reliably differed between these groups, as indicated their different ST, 7.4 vs. 2.1 min (Figure 1A). mKATP channel activity was assessed ex vivo on isolated liver mitochondria using both K+-selective electrode and oxygen consumption assay as we described previously [12]. In agreement with what was shown earlier, the resistance of the rats to physical stress was strongly dependent on their endogenous mKATP channel activity in such a way that HR group of the animals exhibited elevated activity of mKATP channel relative to LR ones (Figures 1A & 1B) [13]. HR and LR groups, in turn, were divided into three subgroups treated with pharmacological mKATP channels blockers, glibenclamide and 5-HD, and with no treatment except physiological solution, and subjected to compulsory swimming with the load. As earlier, mKATP channels activity was estimated after the experiment. Using K+- selective electrode, we have shown that the rate of ATP- sensitive K+ transport constituted 40.0 ± 4.0 nmol K+·min−1·mg-1 in HR group, while LR animals exhibited only 28.0 ± 3.0 nmol K+·min−1·mg−1.
![Figure 1: The impact of mKATP channel activity on the endurance to physical stress. Swimming time (A) and mKATP channel activity (B) were monitored in high and low resistant animals (HR and LR) with no treatment (control) and after the application of glibenclamide and 5-HD in vivo; C – the effect of mKATP channels opening in vitro on the P/O ratio in rat liver mitochondria. M±m, n=9; A, B: * - P<0.05 relative to control (HR, no treatment), # - P<0.05 as compared to control (LR, no treatment); C: * - P<0.05 as compared to control (native liver mitochondria, incubation medium). HR group without pharmacological treatment exhibited higher mKATP channel activity and better endurance to physical stress (Figures 1A & 1B). Unlike this, glibenclamide and 5-HD dramatically reduced physical endurance of the animals and were equally effective in the blockage of mKATP channel _in vivo_ (Figures 1A & 1B). mKATP channel activity in animals treated with mKATP channel blockers was close to standard experimental error (Figure 1B). In parallel experiments, the effects of mKATP channel opening and blockage on mitochondrial functions were studied _in vitro_. mKATP channel activity was assessed as the component of state 4 respiration sensitive to the addition of ATP in the presence of Mg2+ so that the addition of ATP reduced the rate of respiration. mKATP channel in control animals contributed 4.0±1.0 ng- аt.О∙min−1∙mg-1 to state 4 respiration on complex I substrate glutamate. Considering the stoichiometric ratio between the rate of K+ uptake and the rate of respiration, K+/O=10 for complex I substrates, native mKATP channel activity could be estimated as 40 nmol K+·min−1·mg−1 [15]. This well coincided with mKATP channel activity assessed with K+ electrode. The same as _in vivo_, complete blockage of mKATP channel by its blockers was observed _in vitro_ in the presence of mKATP channels blockers in the incubation medium (Figure 1C). Mg∙ATP-dependent component of the respiration in the presence of glibenclamide and 5- HD was reduced to the level of standard experimental error, which proved the blockage of mKATP channel activity _in vitro_. The impact of mKATP channel opening on mitochondrial functions was assessed polarographically. Representative polarographic curves showing the effects of mKATP channels opener diazoxide and the blocker, glibenclamide, are shown on Figure 2. The data showing the modulation of mitochondrial functions by mKATP channel opening were summarized in the Table. In the presence of diazoxide the rate of state 4 respiration in mitochondria oxidizing glutamate was increased because of the activation of K+ cycle, which we have shown earlier, which in rat liver mitochondria was not accompanied by depolarization [16]. The activation of K+ cycle eventually resulted in mitochondrial uncoupling (reduction of respiratory control ratio, RCR), and also in slowing down of the ADP-stimulated respiration (state 3) accompanied by the inhibition of ATP synthesis, which was indicated by the decrease of _J_ADP (Table). The return to state 4 after ADP consumption again was characterized by elevated respiration rate (_J_4ATP) relative to control, and was explained by the activation of K+ cycling. Nevertheless, mKATP channel opening by diazoxide increased the efficiency of ATP synthesis (P/O), which was rather unexpected considering uncoupling effect of diazoxide.](/fulltextimages/1775/fig_1.jpeg)
Figure 1: The impact of mKATP channel activity on the endurance to physical stress. Swimming time (A) and mKATP channel activity (B) were monitored in high and low resistant animals (HR and LR) with no treatment (control) and after the application of glibenclamide and 5-HD in vivo; C – the effect of mKATP channels opening in vitro on the P/O ratio in rat liver mitochondria. M±m, n=9; A, B: * - P<0.05 relative to control (HR, no treatment), # - P<0.05 as compared to control (LR, no treatment); C: * - P<0.05 as compared to control (native liver mitochondria, incubation medium). HR group without pharmacological treatment exhibited higher mKATP channel activity and better endurance to physical stress (Figures 1A & 1B). Unlike this, glibenclamide and 5-HD dramatically reduced physical endurance of the animals and were equally effective in the blockage of mKATP channel in vivo (Figures 1A & 1B). mKATP channel activity in animals treated with mKATP channel blockers was close to standard experimental error (Figure 1B). In parallel experiments, the effects of mKATP channel opening and blockage on mitochondrial functions were studied in vitro. mKATP channel activity was assessed as the component of state 4 respiration sensitive to the addition of ATP in the presence of Mg2+ so that the addition of ATP reduced the rate of respiration. mKATP channel in control animals contributed 4.0±1.0 ng- аt.О∙min−1∙mg-1 to state 4 respiration on complex I substrate glutamate. Considering the stoichiometric ratio between the rate of K+ uptake and the rate of respiration, K+/O=10 for complex I substrates, native mKATP channel activity could be estimated as 40 nmol K+·min−1·mg−1 [15]. This well coincided with mKATP channel activity assessed with K+ electrode. The same as in vivo, complete blockage of mKATP channel by its blockers was observed in vitro in the presence of mKATP channels blockers in the incubation medium (Figure 1C). Mg∙ATP-dependent component of the respiration in the presence of glibenclamide and 5- HD was reduced to the level of standard experimental error, which proved the blockage of mKATP channel activity in vitro. The impact of mKATP channel opening on mitochondrial functions was assessed polarographically. Representative polarographic curves showing the effects of mKATP channels opener diazoxide and the blocker, glibenclamide, are shown on Figure 2. The data showing the modulation of mitochondrial functions by mKATP channel opening were summarized in the Table. In the presence of diazoxide the rate of state 4 respiration in mitochondria oxidizing glutamate was increased because of the activation of K+ cycle, which we have shown earlier, which in rat liver mitochondria was not accompanied by depolarization [16]. The activation of K+ cycle eventually resulted in mitochondrial uncoupling (reduction of respiratory control ratio, RCR), and also in slowing down of the ADP-stimulated respiration (state 3) accompanied by the inhibition of ATP synthesis, which was indicated by the decrease of _J_ADP (Table). The return to state 4 after ADP consumption again was characterized by elevated respiration rate (_J_4ATP) relative to control, and was explained by the activation of K+ cycling. Nevertheless, mKATP channel opening by diazoxide increased the efficiency of ATP synthesis (P/O), which was rather unexpected considering uncoupling effect of diazoxide.
| Respiration rate, ng-at. 0·min-1·mg-1 | RCR | P/O | JADP, nmol·min-1·mg-1 | |||
| RCR | P/O | JADP, nmol·min-1·mg-1 | J4S | J3 | J4ATP | |
| Control | 11.5±1.0 | 92.0±3.0 | 16.7±1.2 | 5.5±0.7 | 1.23±0.08 | 154.0±6.0 |
| Diazoxide | 26.7±1.1* | 65.0±2.0* | 18.8±1.2* | 3.6±0.4* | 1.58±0.12* | 107.0±5.5* |
Table 1: The effect of mKATP channel opening on mitochondrial functions in vitro.
Table 1: The effect of mKATP channel opening on mitochondrial functions in vitro. The incubation medium: 120 mM Table 1: The effect of mKATP channel opening on mitochondrial functions in vitro. The incubation medium: 120 mM KCl, 2 mM Tris-HCl (pH 7.4), 5 mM sodium glutamate, 1 mM KH2PO4, 1 mM EDTA. Diazoxide was added at 0.5 µM. M±m, n = 6. * − p<0.05 as compared to control
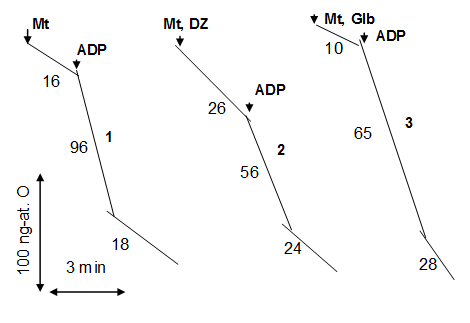
On the contrary, mKATP channels blockers dramatically reduced P/O (Fig. 1C). The reduction of P/O under the action of glibenclamide was accompanied by the inhibition of ATP synthesis and considerable increase of the oxygen consumption under phosphorylation condition (Figure 2). Thus, the experiments have shown that glibenclamide and 5-HD were equally effective in the blocking of mKATP channel both in vivo and in vitro, and dramatic fall of the endurance after their application strongly coincided with the block of mKATP channel and the reduction of phosphorylation efficiency (Figure 1A, C).
Discussion
Direct Impact of mKATP Channel Opening on Mitochondrial Bioenergetics
In the literature no consensus was yet achieved regarding the effect of ATP-sensitive K+ transport on the oxidative phosphorylation, and both the inhibition and the activation thereof by K+ transport [17, 18, 19, 20] were reported. It would seem reasonable to suppose that acceleration of K+ cycle under the action of mKATP channels opener shown in our previous work and simultaneous increase in state 4 respiration should increase the rate of state 3 respiration as well [16]. However, on the contrary, the stimulation of state 4 respiration, was accompanied by the notable suppression of the oxygen consumption in state 3. This was coincident with the decrease in the rate of ATP synthesis (J_ADP) and agreed with our previous data showing slowing down of proton transport measured in the course of phosphorylation [12]. Considering simultaneous inhibition of ATP hydrolysis [12], most likely explanation for the alteration of mitochondrial functions under the action of mKATP channels opener was the inhibition of ATP synthase caused by the activation of K+ cycling. Surprising was only that the energetic efficiency of ATP synthesis (P/O) was increased in parallel with its inhibition (Table). So far, we could propose some considerations aimed to explain observed effects − the inhibition of oxidative phosphorylation caused by the activation of mKATP channel, enhancement of ATP sensitive K+ uptake, the activation of K+ cycle, and the improvement of phosphorylation efficiency. It is well known that the enzymatic activity of F0F1 ATP synthase involves a translocation of protons across mitochondrial membrane [21]. So, it is reasonable to suppose that both K+ uptake and K+/H+-exchange could disturb the coupling between proton transport and ADP/ATP transformations. It is likely that similar uncoupling of the functioning of F0F1 ATP synthase at molecular level was described earlier under the action of K+/H+-ionophore gramicidin [22]. Such uncoupling of the enzymatic activity of F0F1 ATP synthase (called decoupling) was shown to take place even without apparent changes in ΔμН [22]. Thus, potassium uptake and K+ cycling, in addition to classical uncoupling of the respiratory chain caused by mKATP channels opening (by our observations, not capable of so dramatic suppression of phosphorylation under similar uncoupling – _unpublished data), possibly, could result in the decoupling of ATP synthase and the inhibition of ATP synthesis. To explain an increase in P/O caused by mKATP channels opening, it worth mention that “mild” respiratory uncoupling was shown to increase phosphorylation efficiency [23]. Thus, tightly coupled mitochondria exhibit low P/O ratio caused by higher proton leak as compared to partially uncoupled organelles [23]. So, mild uncoupling by diazoxide, possibly, creates conditions optimal to reach higher phosphorylation efficiency relative to control.
Recently, we have shown that the increase of endogenous mKATP channel activity under short-term exposure to acute hypoxia resulted in similar improvement of animals’ endurance, which was blocked by the blockers of mKATP channel [12]. As the purpose of our work was to outline the mechanism whereby the modulation of mitochondrial mKATP channel activity could affect energy metabolism under exercise training, we also studied the effects of mKATP channels blocking on mitochondrial bioenergetics. So, to find the correlation between mKATP channel activity and the animals’ endurance, mKATP channel was blocked by 5- HD and glibenclamide both in vivo and in vitro. As showed the experiments, the blockage of mKATP channel dramatically reduced both the endurance and the efficiency of ATP synthesis, which indicated the dependence of the endurance on P/O ratio and mKATP channels activity (Figures 1A & 1C). This coincided with the ability of mKATP channels blockers to severely reduce the animals’ endurance and completely suppress mKATP channels activity in vivo, which we observed already in our previous work in the rats subject to short-term acute hypoxia [12]. Thus the experiments show a strong correlation between mKATP channels activity, the efficiency of ATP synthesis P/O, and animals’ endurance under exercise training. However, physiological meaning thereof is not clear and needs to be discussed separately.
Physiological Relevance of the Impact of mKATP Channels Opening On the Oxidative Phosphorylation
Adaptive responses to physical stress, ischemic and hypoxic conditions trigger complex network of pathways, such as cytoprotective signaling involving the activation of protein kinases (PI3K, MAPK, PKG and PKC), NO synthase (eNOS), the modulation of glutathione and ATP levels, and the regulation of ROS production [1, 2, 3, 4, 24]. mKATP channels were shown to be a part of the adaptation mechanisms, but physiological relevance thereof remains unclear [4, 24]. In agreement with the literature, we assumed that in energized mitochondria studied in our work the mKATP channel was in the open state [6, 7]. Assuming this, we considered mKATP channels in both high- and low- resistant animals as being in the open state and fully active. However, at present we have not yet data sufficient to explain the differences in the endogenous mKATP channel activity. One of the plausible explanations could be e.g. the different expression of the channels in both groups, but this issue needs more extensive studies. In our work it was shown that mKATP channels opening could largely contribute to the adaptation of a living organism to physical stress by acting on mitochondrial functions, primarily, ATP synthesis. As
showed the experiments in vitro, direct effects of mKATP channels’ opening on the oxidative phosphorylation was the elevation of P/O ratio together with the inhibition of both ATP synthesis and hydrolysis, which was shown in our previous work and the works of other authors [9, 17]. But what could be physiological relevance of such a modulation of ATP synthesis, and why the inhibition thereof could improve the endurance in a living organism? To answer this question, it needs to be considered that ATP synthesis is an oxygen consuming process. Thus, by our hypothesis, suppression of ATP synthesis is aimed at the prevention of the excess oxygen consumption by the tissues under conditions of oxygen deficit, together with the reduction of the energy costs for the phosphorylation. Considering representative polarographic curves showing the effects of mKATP channels opener (diazoxide) and blocker (glibenclamide) on mitochondrial functions (Figure 2), one can see that glibenclamide similarly inhibited ATP synthesis, but, contrary to diazoxide that increased P/O and reduced oxygen consumption, mKATP channel blocker severely reduced P/O and increased oxygen consumption in the course of phosphorylation. Thus, mKATP channels blocker makes ATP synthesis much more energy expensive and highly oxygen consuming process. These considerations, possibly, could be directly applied to the fast exhaustion and dramatic reduction of ST in the animals treated with glibenclamide and 5-HD (Figure 1A). Suppression of ATP hydrolysis because of the direct inhibition of F0F1 ATP synthase by mKATP channels opening was already discussed in the literature, and in agreement with published data, we suppose that it is directed at the preservation of ATP levels needed for muscle tissues under exercise training [9, 17]. Indeed, as it was shown in the literature, mKATP channels openers helped to preserve cellular ATP levels under ischemic conditions [17]. Besides, low ATP levels should keep mKATP channel in functionally active state in order to maintain other physiological functions of the channel, such as the attenuation of ROS overproduction under hypoxic and ischemic conditions, and triggering network of cellular signaling dependent on mKATP channels opening [1]. In addition to what was said above, we would like to hypothesize about the possible compartmentalization of ATP production, and different availability of ATP to different cellular compartments, instead of the “uniform” distribution of ATP in a “bulk” cytosol milieu. Thus, suppression of ATP synthesis and the depletion of ATP nearby ATP binding site of mKATP channel should keep it in highly active state, while the suppression of ATP hydrolysis would preserve cellular ATP needed for the work of actomyosine complex, the ATPases of plasma membrane, and other ATP-consuming enzymes. Under hypoxic conditions suppression of ATP hydrolysis would be helpful also in prevention from the digestion of ATP available from the glycolytic pathway.
Conclusion
Based on the experiments, we came to the conclusion that oxidative phosphorylation system is one of the primary targets of mKATP channel opening under physiological conditions. We propose that suppression of ATP synthesis caused by the elevation of mKATP channel activity could serve a feedback mechanism aimed to keep mKATP channel in highly active state and thereby exert cytoprotective effects dependent on its opening under metabolic stress conditions. The correlation found between mKATP channel activity, increase in phosphorylation efficiency and the improvement of physical endurance of the animals was the evidence for the important role of mKATP channel in the regulation of the energy metabolism in a living organism.
References
-
Garlid KD, Costa AD, Quinlan CL, Pierre SV, Dos Santos P (2009) Cardioprotective signaling to mitochondria. J Mol Cell Cardiol 46(6): 858-866.
-
Yang X, Cohen MV, Downey JM (2010) Mechanism of cardioprotection by early ischemic preconditioning. Cardiovasc Drug Ther 24(3): 225- 234.
-
Correia SC, Cardoso S, Santos RX, Carvalho C, Santos MS, et al. (2011) New insights into the mechanisms of mitochondrial preconditioning-triggered neuroprotection. Curr Pharm Des. 17(31): 3381- 3389.
-
Mironova GD, Shigaeva MI, Gritsenko EN, Murzaeva SV, Gorbacheva OS, et al. (2010) Functioning of the mitochondrial ATP-dependent potassium channel in rats varying in their resistance to hypoxia. Involvement of the channel in the process of animal’s adaptation to hypoxia. J Bioenerg Biomembr 42(6): 473-481.
-
Lucas AM, Caldas FR, da Silva AP, Ventura MM, Leite IM, et al. (2016) Diazoxide prevents reactive oxygen species and mitochondrial damage, leading to anti- hypertrophic effects. Chem Biol Interact 261: 50-55.
-
Costa AD, Quinlan CL, Andrukhiv A, West IC, Jaburek M, et al. (2006) The direct physiological effects of mitoKATP opening on heart mitochondria. Am J Physiol Heart Circ Physiol 290(1): H406-H415.
-
Garlid KD, Paucek P (2003) Mitochondrial potassium transport: the K(+) cycle. Biochim Biophys Acta 1606(1-3): 23-41.
-
Andrukhiv A, Costa AD, West IC, Garlid KD (2006) Opening mitoKATP increases superoxide generation from complex I of the electron transport chain. Am J Physiol Heart Circ Physiol 291(5): H2067-H2074.
-
Belisle E, Kowaltowski AJ (2002) Opening of mitochondrial K+ channels increases ischemic ATP levels by preventing hydrolysis. J Bioenerg Biomembr 34(4): 285-298.
-
Facundo HTF, de Paula JG, Kowaltowski AJ (2007) Mitochondrial ATP-sensitive K+ channels are redox-sensitive pathways that control reactive oxygen species production. Free Radical Biol Med 42(7): 1039-1048.
-
Busija DW, Gaspar T, Domoki F, Katakam PV, Bari F (2008) Mitochondrial-mediated suppression of ROS production upon exposure of neurons to lethal stress: mitochondrial targeted preconditioning. Adv Drug Deliv Rev 60(13-14): 1471-1477.
-
Akopova O, Nosar V, Gavenauskas B, Bratus L, Kolchinskaya L, et al. (2016) The effect of ATP- dependent potassium uptake on mitochondrial functions under acute hypoxia. J Bioenerg Biomembr 48(1): 67-75.
-
Mankovska IN, Nosar VI, Gorbacheva OS, Gonchar OA, Gavenauskas BL, et al. (2014) The effect of uridine on the endurance of animals with different resistance to physical stress: the role of mitochondrial ATP-dependent potassium channel. Biophysics 59(5): 764-767.
-
Baranova OV, Skarga YY, Negoda AE, Mironova GD (2000) Inhibition of DNP-induced potassium efflux by adenine nucleotides in mitochondria. Biokhimia (Moscow) 65: 262-267.
-
Beavis AD (1987) Upper and lower limits of the charge translocation stoichiometry of mitochondrial electron transport. J Biol Chem 262(13): 6165-6173.
-
Akopova OV, Nosar VI, Bouryi VA, Mankovska IN, Sagach VF (2010) Influence of ATP-dependent K+- channel opener on K+-cycle and oxygen consumption in rat liver mitochondria. Biochemistry (Moscow) 75(9): 1139-1147.
-
Dzeja PP, Bast P, Ozcan C, Valverde A, Holmuhamedov EL, et al. (2003) Targeting nucleotide-requiring enzymes: implications for diazoxide-induced cardioprotection. Am J Physiol Heart Circ Physiol 284(4): H1048-H1056.
-
Kopustinskiene DM, Liobikas J, Skemiene K, Malinauskas F, Toleikis A (2010) Direct effects of KATP channel openers pinacidil and diazoxide on oxidative phosphorylation of mitochondria in situ. Cell Physiol Biochem 25(2-3): 181-186.
-
Aon MA, Cortassa S, Wei A-Ch, Grunnet M, O’Rourke B (2010) Energetic performance is improved by specific activation of K+ fluxes through KCa channels in heart mitochondria. Biochim Biophys Acta 1797(1): 71-80.
-
Manon S, Roucou X, Rigoulet M, Guerin M (1995) Stimulation of oxidative phosphorylation by electrophoretic K+ entry associated to electroneutral K+/H+ exchange in yeast mitochondria. Biochim Biophys Acta 1231(3): 282- 288.
-
Seelert H, Poetsch A, Dencher NA, Engel A, Stahlberg H, et al. (2000) Proton powered turbine of a plant motor. Nature 405: 418-419.
-
Koeppe RE, Rottenberg H (1989) Mechanism of uncoupling of oxidative phosphorylation by gramicidin. Biochemistry 28(10): 4355-4360.
-
Stucki JW (1976) Efficiency of oxidative phosphorylation and energy dissipation by H+ ion recycling in rat liver mitochondria metabolizing pyruvate. Eur J Biochem 68(2): 551-562.
-
Murphy E (2004) Primary and secondary signaling pathways in early preconditioning that converge on the mitochondria to produce cardioprotection. Circ Res 94(1): 7-16.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis