Effect of KCl on Biofilm Formation
<p>Aims: Bacteria in biofilms are generally more resistant to environmental stress than their free-living counterparts. In this study, we demonstrated that while bacterial strains Enterococcus faecalis, Escherichia coli and P. aeruginosa displays biofilm formation.</p> <p>Methods and Results: Cells attached to the wells were then fixed with ethanol (%99) for 15 min room temperature and stained with 1% crystal violet. In general, the presence of KCl affects biofilm formation negatively. As temperature increases, biofilm kinetics increases in the presence of KCl. As temperature increases, biofilm kinetics increases in the presence of KCl.</p> <p>Conclusions: We should first mention that studies similar to those we were less common in the literature. We did not find any similar studies with E. coli, E. faecalis and P. aeruginosa in our studies. The work done differs from what we do.</p> <p>Significance and Impact of the Study: The presence of KCl has been shown to influence biofilm formation.</p>
Introduction
Biofilms are increasingly recognized as the predominant form of growth in the environmental life cycle of bacteria. Most of bacteria usually live in a biofilm in their natural environment. Van Leeuwenhoek, using his simple microscopes, first observed microorganisms on tooth surfaces and can be credited with the discovery of microbial biofilms. Most bacteria in natural environments are organized in biofilms. Biofilms can be defined as a community of microbes embedded in an organic polymer matrix, adhering to a surface. Bacteria in biofilms are generally more resistant to environmental stress than their free-living counterparts [1, 2, 3, 4, 5, 6].
A biofilm is an assemblage of a single or multiple species that are encapsulated in self-produced extracellular polymeric substances (EPS). Biofilms in the industrial settings (e.g. ship hulls, water pipes, and membrane filters) cause loss of performance and increased cost for maintenance and quality control. Biofilm cells are known to be 10-1,000 fold more resistant to antimicrobial agents than planktonic cells [7].
Biofilm-forming microorganisms, that P. aeruginosa; B. megaterium, Burkholderia cenocepacia, Citrobacter werkmanii, E. coli, E. faecalis, K. pnemoniae, Listeria monocytogenes, Mycobacterium colombiense, Salmonella typhinurium, Shwanella putrefaciens, S. aureus, S. epidermidis, S. mutans and V. cholera [8, 9, 10, 11].
When bacteria attach to a surface and grow as a biofilm they are protected from killing by antibiotics, biocides, and other chemical or physical challenges. Biofilms are the most common mode of bacterial growth in nature. P. aeruginosa is a model organism for the study of quorum-sensing extracellular virulence factors and biofilm formation [12, 13].
Pseudomonas aeruginosa is a gram-negative bacterium found in almost every ecological niche, including soil, water, and plants. Pseudomonas aeruginosa is the preferred microorganism for the production of rhamnolipid type of biosurfactant using glycerol, mannitol, fructose, glucose, and vegetable oils. While biofilms of the common opportunistic pathogens Staphylococcus epidermidis and Pseudomonas aeruginosa are well known for their antibiotic resistance, diverse other microorganisms form biofilms which confer protection against many antibiotics. In many bacteria protective responses may be deployed in biofilms. Stress responses may be induced in biofilm bacteria by environmental challenge, just as they are in suspended bacteria [12, 14, 15].
Escherichia coli strains (UPEC) can create a specific biofilm a complex intracellular bacterial community (IBC) within the superficial umbrella cells of the bladder, unlike intestinal strains. Biofilm formation on solid surfaces is a very common phenomenon among bacteria with important economic and medical consequences. A strain of Escherichia coli K-12 able to form a thick biofilm on inert surfaces such as glass or polystyrene has previously been isolated from a continuous culture in minimal media [16, 17].
Enterococcus faecalis is a saprophytic commensally that inhabits the oral cavity and gastrointestinal flora of humans and animals, although it can behave as an opportunistic pathogen causing severe urinary tract infections, surgical wound infections, bacteremia, and bacterial endocarditis. Biofilm formation in E. faecalis is presumed to play an important role in a number of enterococcal infections [18, 19].
In this study, we demonstrated that while bacterial strains Enterococcus faecalis, Escherichia coli and P. aeruginosa displays biofilm formation using LB medium or LB medium with 100 mM KCl and temperature and shaker, respectively.
Materials and Methods
Chemicals
KCl, KH2PO4 was purchased from Carlo-Erba. NaCl, Na2HPO4, crystal violet were purchased from Merck. Yeast and peptone were purchased from Mast Diagnostics. Acetic acid was purchased from Acros. Ethanol was purchased from Riedel de Haen. All chemicals used were of analytical grade.
Bacterial Strains
Bacterial strains Enterococcus faecalis (ATCC 25539), Escherichia coli (ATCC 20715) and Pseudomonas aeruginosa (ATCC 27853) were used in this study. Cells were routinely grown in Luria-Bertani broth (LB) at 37°C.
Growth Conditions
Cells were maintained on agar plates at 4 ºC with transfers at monthly intervals. The liquid media used throughout the study was Luria- Bertani (LB) broth medium (g l-1); peptone (10), NaCl (10), and yeast extract (5). The final pH values of broth media was adjusted to 7.0. 100 μl of overnight cultures grown in 20 ml LB in 125 ml Erlenmeyer flasks was inoculated into 50 ml of the same medium in 150 ml volume flasks and incubated for different periods of time. Shake flasks were incubated at 37°C in a 200 rpm gyratory water-bath, drawing the samples at certain intervals. The value (OD600) was increased from 0,5 and diluted to obtain an average of 0,3 - 0,4. A 100 µl culture were inoculums LB medium 10 ml/100 ml Erlenmeyer flasks. 30 or 37°C, 0, 100 rpm or 200 rpm shaker and 100 mM KCl, respectively.
Biofilm Formation
To determine the biomass of static biofilms grown under aerobic conditions, cells were grown on in LB medium 24 h 0, 100 and 200 rpm at 37°C (Figure 1). After the incubation period, the culture supernatant was removed, and both kinds of biofilm tubes were washed four times with 1x phosphate buffered saline (PBS) to eliminate any remaining planktonic cells. Cells attached to the wells were then fixed with ethanol (%99) for 15 min room temperature and stained with 1% crystal violet. After staining, excess crystal violet was eliminated with water, and 33% acetic acid was used to dissolve the remaining dye (Figure 2). Biofilm mass was finally determined as a function of the concentration of this dye based on the absorbance at 570 nm [17, 20, 21, 22].


Results
All of the following results are OD570 values.
Biofilm Formation of Bacterial at Static 30°C
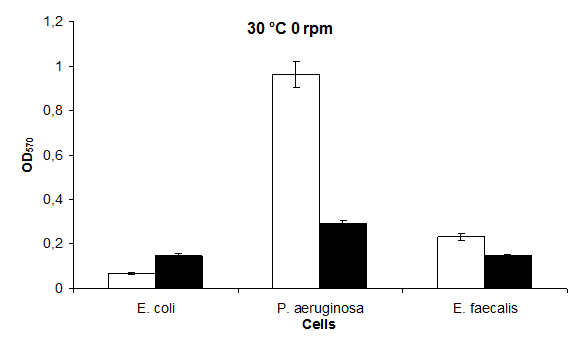
The highest biofilm formation value in the 30°C static environment was in P. aeruginosa; 0,96 and the lowest value in E. coli; it was found to be 0,07. The highest biofilm formation in LB medium containing 30°C KCl was found to be in P. aeruginosa 0,29 and in E. coli and E. faecalis 0,15 (Figure 3).
Biofilm Formation of Bacterial at Static 37 °C
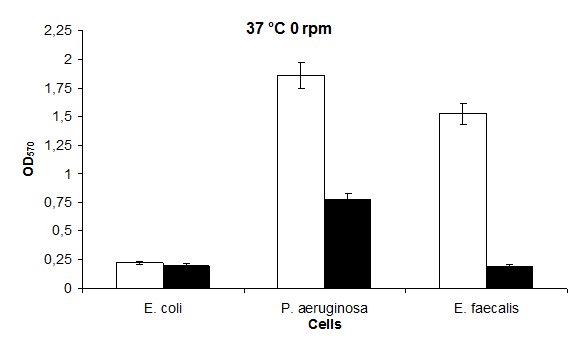
The highest biofilm formation value in 37°C static medium was found to be in P. aeruginosa 1,86 and in E. coli 0,23 (Figures 3 & 4), respectively. The highest biofilm formation in LB medium containing 37°C KCl was found to be in P. aeruginosa 0,78 and in E. coli and E. faecalis 0,20, respectively (Figure 4).
Biofilm Formation of Bacterial at 30°C at 100 rpm
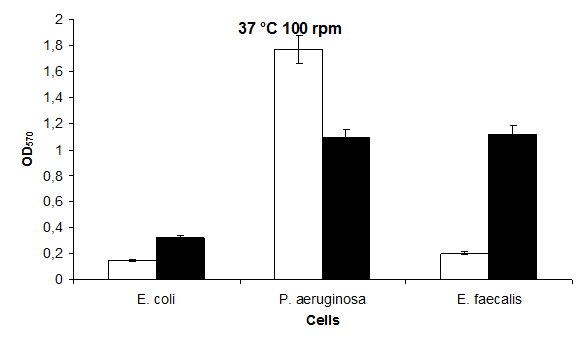
In this study; the highest value at 30°C 100 rpm is in P. aeruginosa 1,21 and the lowest value is in E. coli 0,12 was found. The highest value for biofilm formation at 30°C KCl 100 rpm was in P. aeruginosa 0,77 and in E. coli 0,27 (Figure 5).
Biofilm Formation of Bacterial at 37°C at 100 rpm
At 37°C 100 rpm, the highest biofilm formation value was in P. aeruginosa 1,77 and the lowest value in E. coli;
0,15 was found (Figure 6), respectively. The highest biofilm formation in LB medium containing 37°C 100 rpm KCl was found to be in E. faecalis 1,12 and in E. coli 0,32 (Figure 6), respectively.
Biofilm Formation of Bacterial at 30°C at 200 rpm
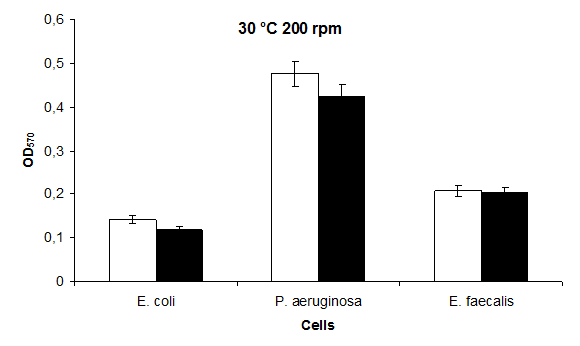
The highest value at 30°C 200 rpm was found to be in P. aeruginosa 0,45 and the lowest value were found to be in E. coli 0,14. The highest biofilm formation at 30°C 200 rpm KCl in LB containing KCl was found to be in P. aeruginosa 0,43 and in E. coli 0,12 (Figure 7).
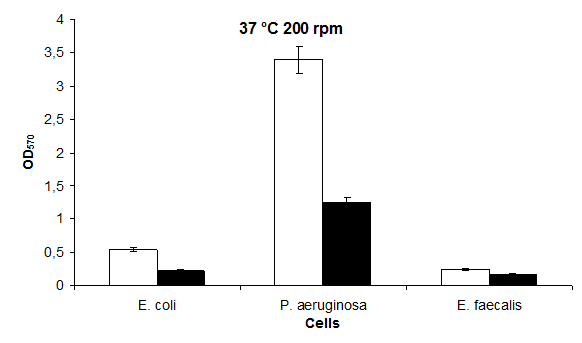
Biofilm Formation of Bacterial at 37°C at 200 rpm
At 37°C 200 rpm, biofilm formation was the highest at P. aeruginosa 3,40 and in E. faecalis 0,25, respectively. The highest biofilm formation in LB medium containing 37 °C 200 rpm KCl was found to be in P. aeruginosa 1,25 and in E. faecalis 0,17 (Figure 8), respectively.
Discussion
In general, the presence of KCl affects biofilm formation negatively. As temperature increases, biofilm kinetics increases in the presence of KCl. The highest biofilm production occurred in E. coli (0.07 A) (Figure 1) at the lowest temperatures of 30°C and 0 rpm at 37°C and 200 rpm in P. aeruginosa (3.40 A) (Figure 6) under normal conditions. Likewise, the highest biofilm production in the presence of 100 mM KCl was found in E. coli (0.12 A (Figure 5) at the lowest 30°C and 200 rpm while the highest biofilm production was at 37°C and 200 rpm in P. aeruginosa (1.25 A) (Figure 8). There is a 49- fold difference between the highest and lowest values (Figures 3 & 8) in the absence of KCl; In the presence of KCl, this difference is 10- fold (Figures 5 & 6).
A similar study to our study was performed with Citrobacter werkmanii and 0.3 Abs values in the presence of 0,6 Abs and 100 mM KCl in 100 mM glucose were obtained [23]. In general, the presence of KCl affects biofilm formation negatively. As temperature increases, biofilm kinetics increases in the presence of KCl. We should first mention that studies similar to those we were less common in the literature. We did not find any similar studies with E. coli, E. faecalis and P. aeruginosa in our studies. The work done differs from what we do.
Conclusion
In this study E. coli and E. faecalis were used to control purposes. In other studies, NaCl was used as stress source. In our study, we used KCl, an unnatural salt, as a source of stress. As a result, biofilm formation increases with increasing temperature and ventilation. However, the biofilm formation, especially in P. aeruginosa, has led decline in the way we expect KCl to exist. P. aeruginosa, which has the ability to form a blend, has the highest biofilm formation under all experimental conditions.
Acknowledgements and Funding
This work was supported by a Grant (APYB 2015/23) from Research Fund Unit of Inonu University. Conflict of interest: Nothing to declare.
References
-
Donlan RM (2002) Microbial Life on Surfaces. Emerging Infectious Emer Infect Dis 8(9): 881-890.
-
Kanamarua S, Kurazono H, Terai A, Mondend K, Kumond H, et al. (2006) Increased biofilm formation in Escherichia coli isolated from acute prostatitis. Int J Antimicrob Agents 28: 21-25.
-
Yang L, Barken KB, Skindersoe ME, Christensen AB, Givskov M, et al. (2007) Effects of iron on DNA release and biofilm development by Pseudomonas aeruginosa. Microbiology 153(5): 1318-1328.
-
Rice SA, Koh KS, Queck SY, Labbate M, Lam KW, et al. (2005) Biofilm formation and sloughing in Serratia marcescens are controlled by quorum sensing and nutrient cues. J Bacteriol 187(10): 3477-3485.
-
Fuente-Núñez C, Korolik V, Bains M, Nguyen U, Breidenstein EBM, et al. (2012) Inhibition of bacterial biofilm formation and swarming motility by a small synthetic cationic peptide. Antimicrob Agents Chemother 56(5): 2696-2704.
-
Kafil HS, Mobarez AM (2015) Assessment of biofilm formation by enterococci isolates from urinary tract infections with different virulence profiles. J King Saud University-Science 27: 312-317.
-
Kim H, Cha E, Kim Y, Jeon Y, Olson BH, et al. (2016) Raffinose, a plant galactoside, inhibits Pseudomonas aeruginosa biofilm formation via binding to LecA and decreasing cellular cyclic diguanylate levels. Nature Sci Rep 6: 25318.
-
Donlan RM (2001) Biofilms and Device-Associated Infections. Emer Infect Diseas 7(2): 277-281.
-
Fux CA, Costerton JW, Stewart PS, Stoodley P (2005) Survival strategies of infectious biofilms. Trends Microbiol 13(1): 34-40.
-
Guerrieri E, Bondi M, Sabia C, Niederhausern S, Borella P, et al. (2008) Effect of Bacterial Interference on Biofilm Development by Legionella pneumophila. Curr Microbiol 57(6): 532-536.
-
Lynch SA, Robertson TG (2008) Bacterial and fungal biofilm infections. Annu Rev Med 59: 415-428.
-
Stewart PS (2002) Mechanism of antibiotic resistance in bacterial biofilms. Int J Med Microbiol 292(2): 107- 113.
-
Qin Z, Yang L, Qu D, Molin S, Tolker-Nielsen T (2009) Pseudomonas aeruginosa extracellular products inhibit staphylococcal growth, and disrupt established biofilms produced by Staphylococcus epidermidis. Microbiology 155(7): 2148-2156.
-
Jackson KD, Starkey M, Kremer S, Parsek MR, Wozniak DJ (2004) Identification of psl, a locus encoding a potential exopolysaccharide that is essential for Pseudomonas aeruginosa PAO1 biofilm formation. J Bacteriol 186(14): 4466-4475.
-
Khezri M, Jouzani GS, Ahmadzadeh M (2016) Fusarium culmorum affects expression of biofilm formation key genes in Bacillus subtilis. Braz J Microbiol 47(1): 47-54.
-
Dorel C, Vidal O, Prigent-Combaret C, Vallet I, Lejeune P (1999) Involvement of the Cpx signal transduction pathway of E. coli in biofilm formation. FEMS Microbiol Lett 178(1): 169-175.
-
Adamus-Bialek W, Kubiak A, Czerwonka G (2015) Analysis of uropathogenic Escherichia coli biofilm formation under different growth conditions. ACTA Biochim Pol 62(4): 765-771.
-
Toledo-Arana A, Valle J, Solano C, Arrizubieta MJ, Cucarella C (2001) The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl Environ Microbiol 67(10): 4538- 4545.
-
Creti R, Koch S, Fabretti F, Baldassarri L, Huebner J (2006) Enterococcal colonization of the gastro- intestinal tract: role of biofilm and environmental oligosaccharides. BMC Microbiol 6: 60.
-
Kim HS, Park HD (2013) Ginger Extract Inhibits Biofilm Formation by Pseudomonas aeruginosa PA14. Plos One 8(9): e76106.
-
Wu C, Cheng Y, Yin H, Song X, Li W, et al. (2013) Oxygen promotes biofilm formation of Shewanella putrefaciens CN32 through a diguanylate cyclase and an adhesin. Nature Sci Rep 3: 1945.
-
Crespo A, Pedraz L, Astola J, Torrents E (2016) Pseudomonas aeruginosa exhibits deficient biofilm formation in the absence of class II and III ribonucleotide reductases due to hindered anaerobic growth. Frontiers Microbiol 7: 688.
-
Zhou G, Li LJ, Shi Q, Ouyang Y, Chen Y, et al. (2013) Effects of nutritional and environmental conditions on planktonic growth and biofilm formation of Citrobacter werkmanii BF-6. J Microbiol Biotechnol 23(12): 1673-1682.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis