Isolation, Production and Optimization of Cellulase from a Combination of Aspergillus Niger and Trichoderma Viride Isolated from Decaying Woods
Cellulase is an enzyme that is capable of hydrolyzing the complex organic form of cellulose into simple sugar (glucose). Cellulase production by a combination of Aspergillus niger and Trichoderma viride isolated from decaying woods were investigated and optimized in this study. Standard methods were used to assay for the cellulase activity. During the time course production of cellulase, it was observed that cellulase production was maximum (9.51μmol/min) at 120 hours of incubation. Increase in substrate concentration increased enzyme activity. Maximum cellulase activity (10.61μmol/min) produced by Aspergillus niger and Trichoderma viride was observed at pH 7 while the optimum temperature was at 35 °C. In conclusion, it might not be suitable to use cellulase from this study above pH 7 and temperature above 35 °C for industrial purposes.
Introduction
Cellulose is a carbohydrate polymer consisting of tens to hundreds to several thousand monosaccharide units (glucose). Cellulose is the main building material out of which plants are made and plants are the primary or first link in what is known as the food chain. It is the most abundant naturally occurring biopolymer [1]. Various natural fibers such as cotton and higher plants have cellulose as their main constituent [2]. It consists of long chains of anhydro-D-glucopyranose units with each cellulose molecule having three hydroxyl groups peranhydro-D-glucopyranose, with the exception of the terminal ends. Cellulose is insoluble in water and most common solvents [1]. The poor solubility is attributed primarily to the strong intramolecular and intermolecular hydrogen bonding between the individual chains [3].
Cellulose can be derived from a number of sources using a number of techniques that are considered synthetic and natural fibers. Natural fibers are made from plant, animal and mineral sources while synthetic fibers generally come from synthetic materials such as petrochemicals but some types of synthetic fibers are manufactured from natural cellulose, including rayon, modal and the more recently developed lyocell [4].
Cellulases are a group of hydrolytic enzymes which are capable of depolymerizing cellulose to smaller molecules. These enzymes are produced mainly by fungi though some bacterial strains have also been found to produce cellulases. The production of cellulase has been reported from a wide variety of bacteria [5] and fungi [6, 7]. However, filamentous fungi are preferred for commercial enzyme production, because the level of the enzymes produced by these cultures is higher than those obtained from yeast and bacteria [8]. Almost all fungi of genus Aspergillus synthesize cellulase, therefore this genus has the potential to dominate the enzyme industry. Aspergillus and Trichoderma species are well known for efficient production of cellulases [9]. Industrially important enzymes have traditionally been obtained from submerged fermentation (SmF) because of the ease of handling and greater control of environmental factors such as temperature and pH.
Cellulases are increasingly being used for a large variety of industrial purposes in the textile industry, pulp and paper industry and food industry, as well as an additive in detergents and improving digestibility of animal feeds. Now cellulases account for a significant share of the world’s industrial enzyme market. The growing concerns about depletion of crude oil and the emissions of greenhouse gases have motivated the production of bioethanol from lignocellulose, especially through enzymatic hydrolysis of lignocellulosic materials sugar platform [10, 11]. However, costs of cellulase for hydrolysis of pretreated lignocellulosic materials need to be reduced and their catalytic efficiency should be further increased in order to make the process economically feasible [12]. It is against this background that this study is carried out to isolate and optimize cellulase from a combination of Aspergillus niger and Trichoderma viride isolated from decaying wood.
Materials and Methods
Decaying wood samples were collected around Adobayero hall, University of Maiduguri, Borno state, Nigeria in a tightly closed container. One (1) g of the sample (decaying wood) was ground and dissolved in 100 ml of Ringers solution. Serial dilution of the sample was made 5 times and cultured on solid media. Each distinct colony was subcultured and incubated for 48 hours. Fungal species Aspergillus niger and Trichoderma viride were identified based on morphological appearance as described by Devanathan, et al. [13] and Barnett, et al. [14] respectively.
Preparation of Inoculum
The fungal species (Aspergillus niger and Trichoderma viride) were inoculated into 100 mL fermentation media containing 0.004 g ZnSO4, 10.5 g KH2PO4, 0.5 g MgSO4·7H2O, 0.5 g CaCl2, 0.13 g FeSO4·7H2O, 0.005 g MnCl2·4H2O, 0.5 g K2HPO4, 0.5 g NaMoO4 and 0.5 g (NH4)2SO4. Incubation was carried out at 37 ºC for 72 hours. All the experiments were done in triplicate and average values were recorded. After 72 hours of incubation the cells were harvested by centrifugation at 10,000 rpm for 15 minutes and the supernatant was used as an enzyme source for estimation of cellulase activity [15].
Assay for Cellulase Activity
The method of Miller, [16] was employed in determining the enzyme activity. 1 ml of crude enzyme filtrate was added to 1 ml of 0.1% (w/v) carboxymethyl cellulose (CMC) in a test tube. Sodium acetate buffer (0.1 mL, pH 6.5) was pipetted into the mixture and then incubated in a Water Bath at 35 °C for 30 min. After incubation, 3 ml of the dinitrosalicylic acid (DNS) reagent was added to terminate the reaction. The tube was boiled for 5 min and monitored for colour development. Optical density readings were taken at 546 nm and the sugar released extrapolated from the standard glucose curve. The result obtained was then expressed as cellulase activity (µmol/min) defined as the enzyme amount that produced 1 µmol of glucose per minute under the assay conditions.
Optimization of Cellulase Production
Effect of incubation time on cellulase production: Effect of incubation time on cellulase production by a combination of A. niger and T. viride was carried out for a period of six (6) days. Enzyme activity was evaluated at an interval of 24 hours. Effect of temperature on cellulase production: Effect of different incubation temperatures (30, 35, 40, 45 and 50°C) on the production of cellulase by a combination of A. niger and T. viride was studied. Cellulase activity was assayed after 120 hours of incubation. Effect of pH on cellulase production: The effect of pH on cellulase activity was determined by varying the pH values (4.0, 5.0, 6.0, 7.0 and 8.0), their influence on cellulase production was determined after 120 hours of incubation. Effect of substrate concentration on cellulase production: The effect of substrate (CMC) concentration on cellulase production was determined by varying the substrate concentration (0.5, 1.0, 1.5, 2.0 and 2.5 mg/ml) enzyme activity was determined after 120 hours of incubation.
Results and Discussion
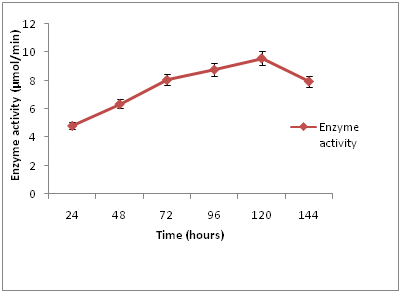
The time course production of cellulase (Figure 1), revealed that maximum enzyme activity (9.51 µmol/min) was at 120 hours of incubation. The lowest cellulase activity (4.82 µmol/min) was observed at 24 hours of incubation. The activity of the enzyme gradually decreased after 120 hours of incubation. Fermentation time has an important impact on product formation and enzyme production, the enzyme activity gradually increased as incubation time increased. Incubation period of 120 hours was optimum for cellulase enzyme production by the isolates. This is comparable with the result of Karthikeyan, et al., [17] who reported incubation time of 120 hours as optimum for the production of cellulase by A. niger. This result revealed increasing trend of enzyme activity up to 120 hours of incubation and then a decline. This may be due to increase in concentration of toxic substances and depletion of nutrients in the fermentation media which led to decreased fungal growth and enzymes production. Another reason may be the high viscosity of the medium, which decreases the oxygen supply to the microorganisms. High viscosity leads to retard cell division, resulting in low production and excretion of cellulase [18].
The effect of temperature on cellulase activity (Figure 2) revealed that maximum cellulase enzyme activity (2.41 µmol/min) was at 35°C after 120 hours of incubation. The lowest cellulase activity (0.35 µmol/min) was observed at 50°C after 120 hours of incubation. Cellulase activity gradually decreases as temperature increases. Temperature is another critical factor that influences enzyme activity and it has to be controlled in any bioprocess. This result is in contrast with the results of [19] who reported 30°C as optimum temperature for cellulase enzyme production. Cellulase is highly sensitive towards temperature and high temperature decreases cellulase activity due to instability [18].
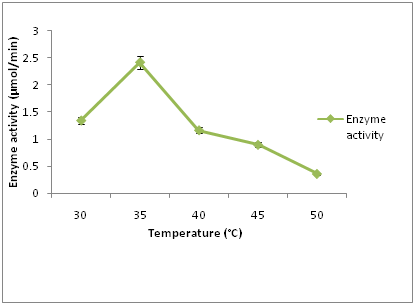
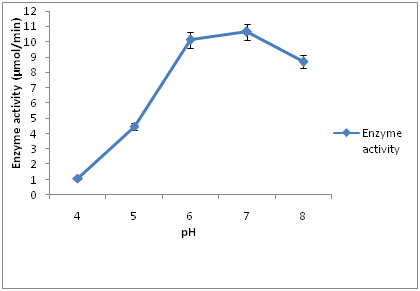
Figure 3 The effect of pH on cellulase activity, maximum cellulase enzyme activity (10.61 µmol/min) was observed at pH 7 after 120 hours of incubation. As the pH increases, decrease in enzyme activity was observed. The minimum cellulase enzyme activity (1.01 µmol/min) was observed at pH 4 after 120 hours of incubation. Different enzymes have different optimum pH values. The pH of the growth medium plays an important role by inducing microbes to secret enzymes and other metabolites [4]. The study conducted showed that cellulase production was greatly influenced by pH of the medium. Cellulase production was significantly inhibited when the pH level of the medium was lower than 5.0. Similar result was reported by Sazci, et al., [20] optimal pH was at 7.0 when cellulase was produced under submerged fermentation.
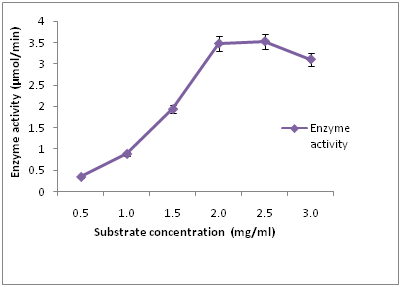
The effect of different concentration of substrate on cellulase activity (Figure 4) showed that the maximum enzyme activity (3.52 µmol/min) was observed at 120 hours when 2.5 mg/ml of the substrate was used. Lowest cellulase activity (0.35 µmol/min) was observed when 0.5 mg/ml of the substrate was used after 120 hours of incubation. All fungi must utilize the energy source in their environment in order to produce ATP. ATP is required by fungi for all of the biosynthetic processes that fungi use for maintenance and reproduction. Carbon is a major requirement of the cell and the rate at which a carbon source is metabolized can often influence the production of metabolites [4]. Among different volumes of substrate studied, 2.5 mg/ml of the substrate showed highest enzyme activity after 120 hours of incubation. As the substrate concentration increases, increase in cellulase enzyme activity was observed. The result was in conformity with the work carried out by Ahmed, et al., [21] who reported that CMC as the best substrate for cellulase production due to its soluble nature compared to other substrates.
Conclusion
This study demonstrates that Aspergillus niger and Trichoderma viride effectively produced cellulase and may be used for industrial production of cellulases. The combined effect of all the growth factors was observed that there was a trend of increase in the cellulase activity at every step of the optimization process which confirms the optimization of the process parameters. Further studies would focus on isolation and purification of cellulase for industrial purposes.
References
-
Bochek AM (2003) Effect of Hydrogen Bonding on Cellulose Solubility in Aqueous and Non-aqueous Solvents. Russian Journal of Applied Chemistry 76(11): 1711-1719.
-
Myasoedova VV (2000) Physical Chemistry of Non- aqueous Solutions of Cellulose and its Derivative. John Wiley and Sons, Chirchester 22: 111-118.
-
Hinterstoisser B, Salmen L (2000) Application of Dynamic 2D FTI to Cellulose. Vibrational Spectroscopy 22: 111-118.
-
Percival Zhang YH, Himmel ME, Mielenz JR (2006) Outlook for Cellulase Improvement: Screening and Selection Strategies. Biotechnol 24(5): 452-481.
-
Immanual G, Dhanusha R, Prema P, Palavesan A (2006) Effect of Different Growth Parameters on Endoglucanase Enzyme Activity by Bacteria Isolated from Coir Retting Effluents of Estuarine Environment. Int J Environ Sci Tech 3(1): 25-34.
-
Anita S, Namita S, Bishnoi NR (2009) Production of Cellulases by Aspergillus heteromorphus from Wheat Straw under Submerged Fermentation. International Journal of Agricultural and Biosystems Engineering 3(3): 124-127.
-
Sarao LK, Arora M, Sehgal VK (2010) Use of scopulariopsis acremonium for the Production of Cellulose and Xylanase through Submerged Fermentation. African Journal of Microbiology 4(14): 1506-1510.
-
Bakri Y, Jacques P, Thonart P (2003) Xylanase Production by Penicillum canescens 10-10c in Solid State Fermentation. Appl Biochem Biotechnol 108: 737-748.
-
van Peij NN, Gielkens MM, de Vries RP, Visser J, de Graaff LH (1998) The Transcriptional Activator X in R regulates Both Xylanolytic Endoglucanase Gene Expression in Aspergillus niger. Appl Environ Microbiol 64(10): 3615-3619.
-
Bayer EA, Belaich JP, Shoham Y, Lamed R (2004) The Cellulosomes: Multienzyme Machines for Degradation of Plant Cell Wall Polysaccharides Annu Rev Microbiol 58: 521-554.
-
Zaldivar J, Nielsen J, Olsson L (2001) Fuel Ethanol Production from Lignocellulose: A Challenge for Metabolic Engineering and Process Integration. Appl Microbiol Biotechnol 56(1-2): 17-34.
-
Sheehan J, Himmel M (1999) Enzymes, Energy, and the Environment: A Strategic Perspective on the U.S. Department of Energy’s Research and Development Activities for Bioethanol. Biotechnol Prog 15(5): 817- 827.
-
Devanathan G, Shanmugan A, Balasubramanian T, Manivannan S (2007) Cellulase Production by Aspergillus niger Isolated from Coastal Mangrove Debris. Trends Applied Science Research 2(1): 23-27.
-
Barnett HL, Hunter B, Prema J (1972) Illustrated Genera of Imperfect Fungi. Burgess publishing company, Minnesota.
-
Ali SA, Sayed RT, Sarker, Alam R (1991) Factors Affecting Cellulase Production by Aspergillus terreus and Aspergillus niger. Enzyme Microbial Technology 11: 606-616.
-
Miller GL (1972) Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Annual Chemistry 31(3): 426-428.
-
Karthikeyan N, Sakthivel M, Palani P (2010) Screening, Identifying of Penicillium K-P Strain and its Cellulase Producing Conditions. Journal of Ecobiotechnology 2: 4-7.
-
Haque UI (1992) Optimization of Cellulase Synthesis by Locally Isolated Trichoderma species using Agricultural By-products as Substrates (Dissertation). Punjab University, Lahore, Pakistan.
-
Maurya DP, Singh D, Partap D, Maurya JP (2012) Optimization of Solid State Fermentation Sonditions for the Production of Cellulase by Trichoderma reesei. J Environ Biol 33(1): 5-8.
-
Sazci A, Radford A, Erenle K (1986) Detection of Cellulolytic Fungi by using Congo red as an Indicator: a Comparative Study with the Dinitrosalicyclic Acid Reagent Method. Journal of Applied Bacteriology 61(6): 559-562.
-
Ahmed S, Aslam N, Naeem S, Jamil A (2003) Induction of Xylanase and Cellulase Genes from Trichoderma harzianum with Different Carbon Sources. Pakistan Journal of Biological Sciences (Pakistan) 6(22): 1912- 1916.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis