High Yield Purification of Mytilidae PP2A: An Enzyme for the Sensitive Detection of Phycotoxins
Diarrheic Shellfish Poison (DSP) toxins such as Okadaic Acid (OA), Dinophisis toxin 1 and Dinophisis toxin 2, and Microcystins such as Microcystin-LR (MC-LR) are widely spread biotoxins whose molecular mechanism of action is based on the specific inhibition of protein phosphatase 2A (PP2A) with costly negative effects on public health, even at lower quantities than those established by international safe limits worldwide, due to their tumor-promoting action and hepatotoxicity. Considering these, a zero tolerance on the human consumption of these phycotoxins should be advocated. The high affinity of these phycotoxins for PP2A allows using the same enzyme as a very sensitive biosensor. Until now, PP2A has been purified following the classic protocol to produce this enzyme that is based on mammalian tissue as the source material. However, it is characterized by its low PP2A content, resulting in low yields per gram of tissue. In this study PP2A was purified from an invertebrate, a native Chilean shellfish classified as Aulacomia atra. This Mytilidae is a common endemic shellfish that evolved together with the dinoflagellate DSP phycotoxin producers in the Chilean littoral. When the tissue of these shellfish is tested for PP2A, the data show an unusually high amount of this enzyme per gram of shellfish, using these shellfish, the yield post-purification was forty times greater than those reported for other mammalian PP2A purifications. This Mytilidae PP2A is characterized by a Vmax of 7.60 pmol min-1 μg protein-1 and a Km of 32.09 mM for p-NPP. The inhibition assay resulted in an IC50 for OA and MC-LR of 0.86 ng mL-1 and 0.437 ng mL-1 , respectively. The enzymatic stability over time was evaluated, showing that the enzyme is best kept at 4°C suspended in 10% of glycerol and as such retained 80% of its enzymatic activity after 2 weeks and 60% after more than 4 weeks. Taken together, these results indicate that PP2A purified from this bivalve filter-feeder mollusk is a good candidate biosensor for the detection and quantification of DSP toxins and MC-LR-like toxins, especially considering the sensibility, this method is at least 360 times lower than international detection limits.
Introduction
Ever since Diarrheic Shellfish Poison was first described [1], a new set of protein phosphatase inhibitors have been isolated from the marine environment. Of all the DSP toxins, Okadaic acid (OA) is the most common biotoxin related to the DSP syndrome [2]. This polyether is a hydrophobic marine phycotoxin produced by the dinoflagellate genus Dinophysis that is common in marine waters. On the other hand, freshwater environments also have microorganisms, such as cyanobacteria from the genus Microcystis, which are capable to produce Microcystins, the most well-known of these is Microcystin LR (MC-LR) [3] and even though these are water soluble, they also specifically inhibit PP2A. In both environments, marine, and freshwater, these microorganisms cause no harm when present in low biomass, but as soon as these microalgae bloom, their biomass multiplies and so do the phycotoxins.
OA and MC-LR are well-known inhibitors of Protein Phosphatase 1 (PP1) and Protein Phosphatase 2A (PP2A) [4, 5], which play central roles in protein dephosphorylation in living cells. OA produces protein hyperphosphorylation, which deregulates sodium excretion by intestinal cells, causing sodium release that produces passive losses of fluids, which are responsible for the diarrheic symptoms of DSP [6]. In the case of MC- LR dephosphorylation is inhibited in the cytoplasm of liver cells, causing associated hepatotoxicity [7].
Humans are intoxicated with DSP toxins by eating shellfish contaminated with these phycotoxins, which accumulate these in the midgut gland after filter feeding on DSP toxin producing microalgae [8]. A safe limit of 160 μg OA equivalents/kg shellfish meat has been established by the Food and Agriculture Organization [9], but during recent years studies have reported that OA concentrations below this safe limit stimulate the secretion of inflammatory factors by innate immune cells [10], which are linked to chronic inflammatory processes and cancer [11]. PP2A has a higher affinity for MC-LR than for OA and MC-LR thus is the stronger PP2A inhibitor (2- to 10-fold more than OA) [12]. This remarkable observation is the reason that the authors proposed zero tolerance to DSP toxins in shellfish for human consumption [11]. Hence it is imperative to have a highly sensitive tool in order to monitor and quantify these phycotoxins to prevent contact with and deleterious effects on the human population [13, 14].
For the detection of these toxins, the mouse bioassay was the first method of screening [1], but it has been questioned because of ethical issues, the high cost of animal maintenance, and due to specificity and sensitivity problems. There are alternatives, easy-to-perform bioassays available, but these are time consuming and subjective, making it difficult to unambiguously interpret the results [15]. At the other end of the spectrum there are chromatographic methods, such as for instance high- performance liquid chromatography (HPLC), which are satisfactory, sensitive and accurate, but also require expensive equipment, as well standards and a trained and dedicated workforce that has to invest time and resources in the analysis of each sample [3, 16].
On the other hand, recent years saw the development of an enzymatic assay based on the inhibition by these phycotoxins of their specific targets, the protein phosphatases [12, 17] like, for example, PP2A that undergoes a powerful enzyme-ligand interaction which totally inhibits the enzyme [4, 18]. Because PP2A has a high affinity for toxins as OA and MC-LR, with a range of inhibition between 0.02-0.2 nM [6, 19], the development of an enzymatic assay using PP2A allows a very sensitive and quantitative analysis [15, 20]. Some technological challenges currently impede the widespread use of this enzymatic assay such as cost of the enzyme, and characteristics such as its instability at room temperature that requires a strict cold chain, making transport to places far from the manufacturers difficult. Therefore, it is imperative to develop a method for the low-cost, massive production of active PP2A enzyme to be used as a tool in basic scientific research and biotechnology applications. To fulfill this need, we here describe a purification method to produce active PP2A at unprecedented yields.
Materials and Methods
Collection of Samples
Samples of filtering Mytilidae (Aulacomia atra) were collected in the “Seno del Reloncaví”, Puerto Montt, Chile (41°42′ S, 72°49′ W), and kept cold at 4°C during their transfer to Santiago where these were stored at -20°C for further use.
Protein Phosphatase 2A Extraction and Purification
The whole purification process was carried out at 4°C and the enzymes were purified from seafood tissue using the protocol of Tran [21] modified. In short, 10 g of shellfish meat, previously washed with cold water and dried superficially with absorbent paper, was homogenized with 15 mL of extraction buffer (10 mM MOPS, 2 mM MgCl 2, 1 mM DTT, 1 mM TCPK, 1 mM benzamidine, 0.1% PMSF in 2% methanol, pH 7.5) using the bio-homogenizer "biospec 985-370" in 50 mL tubes (extract identified as sample"M1”) and then centrifuged at 16,000 RCF for 10 min (Sorvall RC 6). The supernatant was retained and filtered using 4 layers of gauze. Two volumes of 95% ethanol (with 1 mM PMSF) were added and the mixture centrifuged at 7500 RCF for 5 min. The supernatant was retained (identified as sample "M2"), diluted in an elution solution (10 mM MOPS, 50 mM NaCl, pH 7.5 maintained between 0-4°C) and batch-mixed with anion exchange resin ChromSpeed™ DA103 (Mitsubishi Chemical) previously equilibrated with elution buffer. PP2A remained unbound in the supernatant whereas contaminating proteins were captured by the DA103 resin After 15 min incubation with the resin at 4°C and occasional agitation with a glass stirrer the protein extract was centrifuged at 29000 RCF for 3 min and the supernatant (identified as sample "M3") was distributed in aliquots of 1.5 mL volumes and stored at +4 and -20°C until further use.
Purified PP2A Quantification
Protein concentration was determined by the Bradford method [22] (BIO-RAD, Richmond, CA, USA) using bovine serum albumin (BSA) as the protein standard and triplicate dilution series of 0, 1, 2, 4, 6, 8 and 10 μg mL-1 in. The samples were mixed with the reagent and read after 5 min at 595 nm in a spectrophotometer (Epoch, BioTek Instruments Inc., model EPOCHH) and their concentrations deduced from the standard curve.
Polyacrylamide-gel Electrophoresis and Western Blot
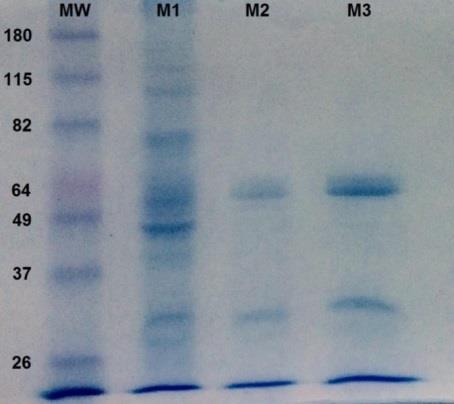
Samples from each of the purification stages (M1, M2, and M3), were analyzed via sodium dodecyl sulfate polyacrylamide-gel electrophoresis (SDS-PAGE) and compared to a protein standard with known molecular weight (MW). Hereto, 10 μg of total protein was mixed with 10 μL of buffer (0.0625 M Tris-HCl, pH 6.8 glycerol 10% sodium dodecyl sulfate 2.3% (w/v), 2- mercaptoethanol 5%, bromophenol blue 0.00125% (w/v)) and homogenized for 1 min in vortex. Proteins were separated on gel (3% and 12.5% polyacrylamide, for the concentrator and separator gel, respectively) submerged in running buffer (0.025 M Tris-HCl, 192 mM glycine, 3.5 mM SDS) using a 100 and 150 volt electrophoresis chamber for the separation and staking processes equally. Gels were stained with 45% methanol, 10% glacial acetic acid and 0.25% Coomassie blue G-250 (w/v) and destained with 45% methanol and glacial acetic acid 10% until the protein bands were satisfactorily visualized.
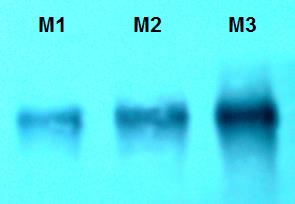
PP2A was identified in samples via Western Blotting with a monoclonal antibody that recognizes a conserved epitope in the catalytic subunits of Homo sapiens PP2A (05-421, Merck, Millipore). Hereto, 10 μL of each extract was separated by SDS-PAGE, transferred to an Immobilon membrane (PVDF Millipore), blocked with BSA and incubated with a 0.1 μg/mL dilution of the PP2A-specific monoclonal in PBS-BSA. Then, the membranes were washed and incubated with a secondary anti-mouse IgG antibody labeled with alkaline phosphatase (Promega). Subsequently, the membranes were washed and developed with the substrates NBT (nitroblue tetrazolium) and BCIP (5-bromo-4-chloro-3-indolyl- phosphate).
Purified PP2A Enzymatic Activity and pH Optimum
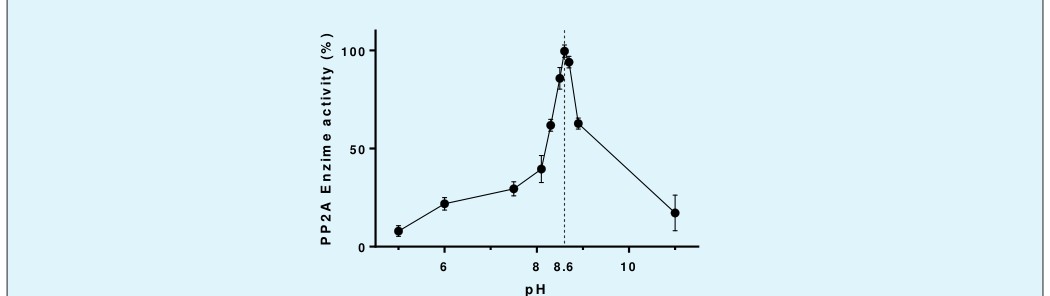
The phosphatase activity of purified PP2A was assayed by cleavage of the phosphate moiety from the substrate para-nitrophenolphosphate (p-NPP) and the subsequent colorimetric reaction (monitored at 405 nm). To determine the pH at which PP2A is maximally active, this reaction was carried out at pH 5; 6; 7.5; 8.1; 8.3; 8.5; 8.6; 8.7; 8,9 and 11 according to Prickett and Brautigan where 20 μL of 5X buffer (200 mM Tris, 20 mM EDTA, 20 mM DTT, buffer concentration) is mixed with 150 μL (25 μg) of stabilized PP2A enzyme, 10 μL of p-NPP at saturation (433 mM) and 20 μL of methanol [23]. The mixture was homogenized in by vortexing and preincubated for 5 min at 4°C. The enzymatic activity was monitored for 45 min at 24 °C and at 405 nm.
PP2A Enzymatic Activity in Function of the p-NPP Concentration and in the Presence of its Specific Inhibitors OA and MC-LR
The PP2A enzymatic activity was analyzed as described above using different concentrations of substrate (16, 32, 75, 96, 128 and 280 mM of p-NPP) for 45 min. Then the PP2A activity was assayed at optimum pH (8.6) and substrate saturation (433 mM) in the function of the different final concentration of its inhibitors OA and MC-LR (0 ng mL-1; 0.125 ng mL-1; 0.25 ng mL-1; 0.5 ng mL-1 and 1 ng mL-1).
PP2A Enzymatic Stability in Function of Time and Storage Conditions
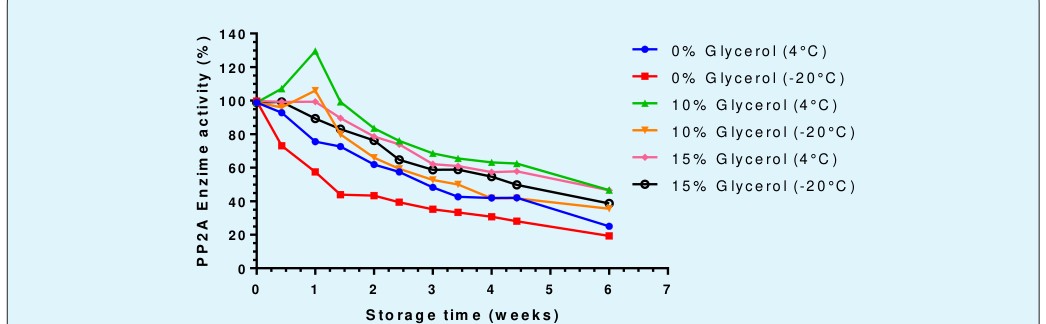
PP2A enzyme was stored at different glycerol concentrations (0, 10, and 15%) and kept at -20 or 4°C, for six weeks. Its remaining enzymatic activity was regularly measured under optimal reaction conditions (pH 8.6 and substrate saturation) to determine the expiration tendency of the enzyme under the different storage conditions.
Results
The purification of PP2A was developed from the total tissue of Mytilidae, by making modifications of the method described by Tran [21]. It was observed that during the purification process (Table 1 and Figure 1), in the 95% ethanol step (M2), most of the total proteins are precipitated, except for the PP2A enzyme. In the resin step (M3), works for a final purification phase.
| Extract | Total protein (mg) |
|---|---|
| M1 | 1,21 |
| M2 | 0,503 |
| M3 | 0,235 |
Table 1: Total protein obtained from the of Mytilidae processed to acquire PP2A, considering the initial shellfish extract (M1),
In the SDS-PAGE (Figure 1) obtained, a band of 65 KDa is appreciated that corresponds to the scaffold subunit (A). Another band of 28 KDa is also observed, corresponding to the catalytic subunit (C), which was corroborated with the Western Blot assay (Figure 2).
When his enzymatic activity was tested with p-NPP as substrate, a classical progress curve was obtained (Figure 3). Then, in order to acquire the optimal conditions for the enzymatic activity through the use of p-NPP as a substrate, an optimum pH of 8.6 was found (Figure 4), achieved from a very pronounced pH curve, that it has an activity with pronounced pH dependence. This optimum pH agrees with the data described in the literature [24].
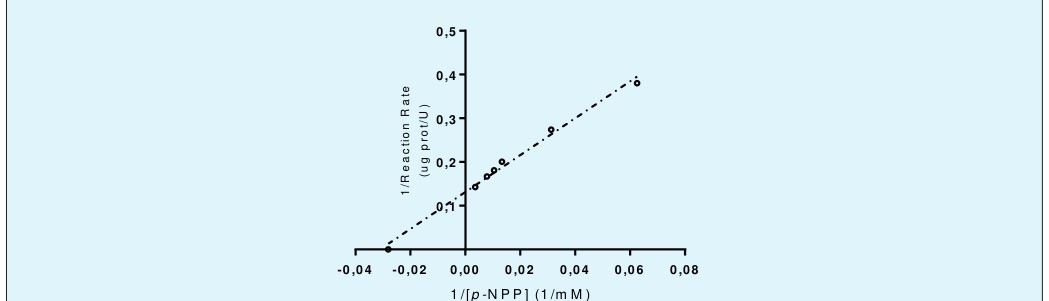
For the enzyme characterization, the kinetic constants were calculated. Thus, for this enzyme a typical Michaelian curve is shown (Figure 5), which begins to saturate at concentrations of 75 mM of p-NPP, this condition becomes more evident at 280 mM of the substrate. Comparing these results with those of the double reciprocal graph Lineweaver-Burk (Figure 6), a Vmax of 7.604 pmol min-1 μg protein-1 and the Km of 32.09 mM were obtained. Both values are within 95% confidence. The trend of Lineweaver-Burk has an R2 of 0.991, implying robust results for these constants.
9), the best in stability is at 4°C, independent of the preservatives. However, the preservation qualities of glycerol, which increase by its concentration, therefore the density of the solution, protects the enzymatic activity by decreasing the deleterious effects of the temperature of -20°C and storage time. However, storage stability with higher activities was achieved at 4°C with 10% glycerol, which decreases enzymatic activity to 25% during the initial 18 days. Different is the case with glycerol 15% that also shows a protective effect, but it is not superior to the previous one. In addition, from a week and a half, they have a higher enzymatic activity than the condition of - 20°C and 10% glycerol.
When PP2A are with phycotoxins, the IC50 values (Figures 7 and 8) shows that the most potent PP2A inhibitor is MC-LR from the toxins tested, where a 50% inhibition was achieved with a toxin concentration of 0.439 nM (0.437 ng mL-1). When comparing this value with the IC50 of 1.06 nM (0.87 ng mL-1) for OA [3, 9, 25].
R e a ctio n R a te Figure 5: Substrate Effect Curve (Michaelis-Menten): Average of tests (n = 8) of enzymatic activity of PP2A at increasing concentrations of substrate p-NPP at pH 8.6.
From the results obtained, it is important to note that researchers such as Tran [21] and Takai [6], applying their purification method, used high amounts of tissue (4 kg of bovine heart or rabbit skeletal muscle), managing to extract 1.6 g of PP2A. Using Mytilidae tissue, with 100 g of tissue is possible to produce the same amount of PP2A enzyme starting from mammal tissue, being forty times less tissue.
The characterization and total purification of the PP2A enzyme, it is confirmed by the SDS-PAGE and Western Blot standards. The PP2A hitherto described are heteromeric proteins, formed by two (A and C) or three (A, B and C) subunits. The A subunit or scaffold has a molecular weight that varies from 60-67 KDa, the B subunit of 24-74 KDa, according to its family, and the C or catalytic subunit varies from 25-36 KDa. This range of molecular weights is presented by practically all PP2A that have been isolated from a variety of mammalian tissues forming different complexes with different molecular weights [26]. Highlighting that an human-PP2A antibody was used, these results shows the conservation of the purified PP2A from an invertebrate such as a Mytilidae, which, since his relevance in the cellular processes, must be expected [5, 27, 28].
For the PP2A kinetic constants, as an antecedent to these values, Bialojan and Takai [29], using as substrate Phosphorylase A obtain a Km of 31 μM and a Vmax of 7350 pmol min-1 μg protein-1 for the PP2A of rabbit skeletal muscle. Unlike the kinetic constants obtained, a different substrate used must be considered, being Phosphorylase A an endogenous PP2A substrate closer to cellular physiology, with an affinity 100 times greater than p-NPP. Regarding the use of p-NPP as a substrate, Takai [24] obtained a Km of 4.8 mM and a Vmax of 233 pmoles min-1 μg protein-1. This data showed greater affinity for rabbit PP2A using the same substrate. However, both Michaelian constants are in the same order of magnitude indicating that they have similar affinities to the same substrate.
About the IC50 values obtained, the MC-LR had a 2.4- fold higher inhibition. These results confirm that this enzyme can be used as a target protein to detect such toxins present in marine and freshwater aquaculture, which, in case of MC-LR are contaminants and indicators of eutrophication in lakes, lagoons and rivers around the world, being useful in the quantification of the phycotoxins produced by Microcystis auroginosa, already specified in previous publications [19, 3, 12].
The calibration curves obtained demonstrate that these phycotoxins can be quantified with high sensitivity and reproducibility, which despite not being within the highest sensitivities, such as those demonstrated by Ikehara [17], it has to be considered that the sample volume used in the assay is 20 μL, and knowing that the IC50 obtained for OA is 0.86 ng mL-1, a detection test would require 16.1 picograms to be at this level. Considering the international safe limit of 160 μg of OA equivalent per one kg of shellfish meat, these values corresponds to 3200 picograms of OA for the proposed test., being more than 360 times the amount required for the IC50. There is evidence in the literature that suggests the use of the inhibitory test of PP2A as a method to detect OA and MC-LR [3, 12, 17, 30], since this enzymatic method is more sensitive than HPLC with Fluorescence on line detection. Therefore, an ideal method for detection in samples with low levels of phycotoxins with values under the safe limits [31, 32, 33]. Additionally, in the case of MC-LR, the IC50 for the Mytilidae PP2A is 0.437 ng mL-1, making the possibility of the use of PP2A enzymatic assay in the detection of this phycotoxin in freshwater samples, since M. aeruginosa is systematically present in the freshwater harmful algal blooms in practically the entire world [3, 9, 25].
The enzymatic activity of Mytilidae PP2A presents a progressive decrease in time, which not only depends on the storing preservatives, it is also temperature dependent. The Results demonstrate that storage at -20 °C is the worst condition among its variables, adding simplicity to the preservation of the purified Mytilidae PP2A enzyme since it can be stored in a normal refrigerator.
Conclusions
Considering the results obtained, the capacity of the PP2A purified from Aulacomia atra as a biotechnological tool to quantify AO and MC-LR is shown, suggesting its use for the VDM and Microcystins produced by eutrophied cyanobacteria in freshwater aquaculture, with levels of detection within the current regulations and with great sensitivity. This allows the realization of a platform that is specific, reproducible, rapid, massive, friendly and economical. Granting the ability to monitor the coast in a short time, strengthening the shellfish industry and protecting the public health of our country. It is important to note that the implementation of an enzymatic assay based on the inhibition of PP2A helps to improve the development of the observation of the seafood resource. Allowing the harvest of the cultures of mussels (and other bivalves for human consumption) during generalized closures, including the timely use of crops that have levels lower than 160 μg of AO per kg of shellfish meat, strengthening the development of purification processes and elimination of toxins from contaminated shellfish. This allows all sectors associated with the exploitation of the seafood resource, to know the presence of contamination of their products in real time, developing processes that allow the management and rational use, making informed decisions making the industry more competitive.
Acknowledgements
This study was supported by FONDECYT Grant #1130037 (N. Lagos) and FONDEF IDeA ID16I10145. Both sponsored by Chilean Government programs.
References
-
Yasumoto T, Oshima Y, Yamaguchi M (1978) Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull Jpn Soc Sci Fish 44(11): 1249- 1255.
-
Reguera B, Velo-Suárez L, Raine R, Park MG (2012) Harmful Dinophysis species: A review. Harmful Algae 14: 87-106.
-
Lagos N (1998) Microalgal blooms A global issue with negative impact in Chile. Biological Research 31(4): 375-386.
-
Xing Y, Xu Y, Chen Y, Jeffrey PD, Chao Y, et al. (2006) Structure of protein phosphatase 2A core enzyme bound to tumor-inducing toxins. Cell 127(2): 341- 353.
-
Xing Y, Li Z, Chen Y, Stock JB, Jeffrey PD (2008) Structural mechanism of demethylation and inactivation of protein phosphatase 2A. Cell 133(1): 154-163.
-
Takai A, Murata M, Torigoe K, Isobe M, Mieskes G, et al. (1992) Inhibitory effect of okadaic acid derivatives on protein phosphatases. A study on structure- affinity relationship. Biochemical Journal 284(2): 539-544.
-
Lad AC, Spegele A, Kleinhenz A, Khalaf FK, Zhang SG, et al. (2018) Evidence of Hepatotoxicity Following Chronic Low Dose Exposure to Microcystin-Lr in a Murine Model of Non-Alcoholic Fatty Liver Disease. Journal of Investigative Medicine 66(4): 824-824.
-
Uribe JC, García C, Rivas M, Lagos N (2001) First report of diarrhetic shellfish toxins in magellanic fjords, Southern Chile. J Shellfish Res 20: 69-74.
-
FAO (2004) Marine Biotoxins. Rome, FAO.
-
del Campo M, Toledo H, Lagos N (2013) Okadaic acid toxin at sublethal dose produced cell proliferation in gastric and colon epithelial cell lines. Mar Drugs 11(12): 4751-4760.
-
Del Campo M, Zhong TY, Tampe R, Garcia L, Lagos N (2017) Sublethal doses of dinophysistoxin-1 and okadaic acid stimulate secretion of inflammatory factors on innate immune cells: Negative health consequences. Toxicon 126: 23-31.
-
Ikehara T, Imamura S, Oshiro N, Ikehara S, Shinjo F, et al. (2008) A protein phosphatase 2A (PP2A) inhibition assay using a recombinant enzyme for rapid detection of microcystins. Toxicon 51(8): 1368- 1373.
-
Fujiki H, Suganuma M (2009) Carcinogenic aspects of protein phosphatase 1 and 2A inhibitors. Prog Mol Subcell Biol 46: 221-254.
-
Fujiki H, Sueoka E, Suganuma M (2013) Tumor promoters: from chemicals to inflammatory proteins. J Cancer Res Clin Oncol 139(10): 1603-1614.
-
Prego-Faraldo MV, Valdiglesias V, Méndez J, Eirín- López JM (2013) Okadaic acid meet and greet: an insight into detection methods, response strategies and genotoxic effects in marine invertebrates. Mar Drugs 11(8): 2829-2845.
-
Garcia C, Rodriguez-Unda N, Contreras C, Barriga A, Lagos N (2012) Lipophilic toxin profiles detected in farmed and benthic mussels populations from the most relevant production zones in Southern Chile. Food Addit Contam Part A Chem Anal Control Expo Risk Assess 29(6): 1011-1020.
-
Ikehara T, Imamura S, Yoshino A, Yasumoto T (2010) PP2A inhibition assay using recombinant enzyme for rapid detection of okadaic acid and its analogs in shellfish. Toxins (Basel) 2(1): 195-204.
-
Moorhead GB, Haystead TA, MacKintosh C (2007) Synthesis and use of the protein phosphatase affinity matrices microcystin-sepharose and microcystin- biotin-sepharose. Methods Mol Biol 365: 39-45.
-
Honkanen RE, Codispoti BA, Tse K, Boynton AL, Honkanan RE (1994) Characterization of natural toxins with inhibitory activity against serine/threonine protein phosphatases. Toxicon 32(3): 339-350.
-
Hayat A, Barthelmebs L, Marty JL (2012) A simple colorimetric enzymatic-assay for okadaic acid detection based on the immobilization of protein phosphatase 2A in sol-gel. Appl Biochem Biotechnol 166(1): 47-56.
-
Tran HT, Ferrar TS, Ulke-Lemée A, Moorhead GB (2007) Purification of PP2Ac from bovine heart. Methods Mol Biol 365: 127-132.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
-
Prickett TD, Brautigan DL (2006) The alpha4 regulatory subunit exerts opposing allosteric effects on protein phosphatases PP6 and PP2A. J Biol Chem 281(41): 30503-30511.
-
Takai A, Mieskes G (1991) Inhibitory effect of okadaic acid on the p-nitrophenyl phosphate phosphatase activity of protein phosphatases. Biochemical Journal 275(1): 233-239.
-
Sukenik A, Quesada A, Salmaso N (2015) Global expansion of toxic and non-toxic cyanobacteria: effect on ecosystem functioning. Biodiversity and Conservation 24(4): 889-908.
-
Shi Y (2009) Assembly and structure of protein phosphatase 2A. Sci China C Life Sci 52(2): 135-146.
-
Eichhorn PJ, Creyghton MP, Bernards R (2009) Protein phosphatase 2A regulatory subunits and cancer. Biochim Biophys Acta 1795(1): 1-15.
-
Liu C, Gotz J (2013) How it all started: tau and protein phosphatase 2A. J Alzheimers Dis 37(3): 483-494.
-
Bialojan C, Takai A (1988) Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem J 256(1): 283-290.
-
Sassolas A, Catanante G, Hayat A, Marty JL (2011) Development of an efficient protein phosphatase- based colorimetric test for okadaic acid detection. Anal Chim Acta 702(2): 262-268.
-
Ramstad H, Shen JL, Larsen S, Aune T (2001) The validity of two HPLC methods and a colorimetric PP2A assay related to the mouse bioassay in quantification of diarrheic toxins in blue mussels (Mytilus edulis). Toxicon 39(9): 1387-1391.
-
Prassopoulou E, Katikou P, Georgantelis D, Kyritsakis A (2009) Detection of okadaic acid and related esters in mussels during diarrhetic shellfish poisoning (DSP) episodes in Greece using the mouse bioassay, the PP2A inhibition assay and HPLC with fluorimetric detection. Toxicon 53(2): 214-227.
-
Kacem I, Bouaïcha N, Hajjem B (2010) Comparison of Okadaic Acid Profiles in Mussels and Oysters Collected in Mediterranean Lagoon, Tunisia. International Journal of Biology 2(2): 238-245.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis