Guanine Rich Oligonucleotide at Survivin Promoter (SVN23), Kills Effectively in Advance Neuroblastoma with MYCN Amplification
Neuroblastoma is derived from the neural crest cells when they are unable to differentiate into mature nerve cells. The clinical hallmark of neuroblastoma is heterogeneous. A subset of neuroblastoma undergoes spontaneous regression while others with unfavorable biology of n-Myc amplification show relentless progression. Current knowledge suggests that Guanine-Rich Oligonucleotides (GRO) are a potential target for anti-cancer drugs. In this study, it has been shown by circular dichroism (CD) that GROs at the promoter regions of the Survivin (SVN-23) gene could form quadruplex in-vitro. Three human neuroblastoma cell lines were used to explore the effect of SVN-23 on different biological properties: cellular growth, differentiation and death. The neuroblastoma cell lines SH-SY5Y and SK-N-AS have a single copy of the MYCN gene (n-Myc amplified), whereas the SK-N-BE2 cell line has multiple copies. In the SK-N-SH cell line, the neuroblastoma cell line consists of both neuronal and epithelial phenotypes, whereas the SH-SY5Y neuroblastoma cell line, which is derived from SK-N-SH cells, are predominantly the epithelioid phenotype. Cellular differentiation of the SVN-23 cell line is induced by the extension of their neurites followed by cell death in the SH-SY5Y cell line, whereas differentiation in the SKN-SH cell line is induced by flattening the cells. Significant growth inhibition has been shown with SVN-23 in n-Myc-amplified SK-N-BE2 cells, (40-75%), whereas modest growth inhibition (10-40%) has been shown in single copy neuroblastoma cell lines (SK-N-AS & SH-SY5Y) by MTT [3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide] assay. This data indicates that SVN-23 is more sensitive to n-Myc amplified neuroblastoma cells than those of neuroblastoma cells with a single copy of n-Myc gene. This sensitivity of SVN-23 is associated with down regulation of survivin protein expression. This data suggest that SVN-23 oligonucleotides could be a potential therapeutic option for unfavorable, n-Myc-amplified groups of human neuroblastoma.
Introduction
Neuroblastoma is the most common childhood solid cancer and accounts for 7 to 10% of all childhood cancers worldwide. Induction of neuronal differentiation has been considered as a novel strategy to treat a certain group of neuroblastoma with aggressive behavior [1]. In aggressive cases of neuroblastoma, the survival rate is 40%. Neuroblastoma originates from neural crest cells during development of the nervous system. Normally, the neural crest cells are involved in the development of the nervous system, but if there is a change in biology (mutation of the gene/genes, abnormal expression of proteins), a consequence that results would be neuroblastoma [2]. Clinically, the symptoms and signs of this cancer vary from person to person. The symptoms usually appear from an early age, even at birth, and there is a high frequency of the spreading of the disease at diagnosis. There are certain subsets of neuroblastoma that have the tendency for spontaneous resolution in infancy. For example, some stage 4S cases (spread only in the bone marrow) diagnosed with neuroblastoma at birth disappear after a few months after birth [3]. A wide variety of genetic alterations have been shown to be associated with different stages of neuroblastoma, including but not limited to, Alk, TrkA, DBNF, GDNF, MYCN, and survivin. High expression of survivin and amplification of the MYCN gene are associated with unfavorable groups of neuroblastoma [3]. The low expression of survivin and the single copy of the MYCN gene are associated with favorable outcome of the disease [4]. Neuronal differentiation refers to the spontaneous maturation of cancer cells. The exact mechanism of certain subsets of neuroblastoma regression is yet to be known in detail, but certain evidence suggests that the self-regression happens through the process of differentiation of the cancer cells. The role of the deprivation of the growth factor (neurotrophic growth factor), loss of telomerase and some other genetics (DNA change) and epigenetics (change of the phosphate group of the DNA) changes have also been proposed [5].
Normally, DNA forms a double helix, which is made up of two polynucleotide strands that spiral around an imaginary axis. Throughout the genome, there are a lot of GC (guanine and cytosine) rich regions, especially in the promoter region of the genes and telomeres (end of the chromosome) [6]. These oligonucleotides of GC-rich regions often naturally form a quadruplex (four-stranded) rather than a double helix in a three-dimensional structure. The ability of the oligonucleotides for encoding genomic G-quadruplex forming sequences has been shown in the silencing of the genes that regulate cancer formation and cellular maturation/ differentiation [7].
The formation of proteins is one of the indicators of gene expression. The information in the DNA transcribes the synthesis of RNA, also known as mRNA because it transfers a genetic message from the DNA to the ribosomes, organelles that transform the linking of amino acids into polypeptide chains. The information in the mRNA translates to the synthesis of a polypeptide. The translation of the nucleotide sequence to the amino acid sequence is handled by the cell through means of the ribosomes [8]. N-Myc and IAP (Inhibitor of Apoptosis Protein) are genes that regulate genetic expression, synthesize proteins (MCYN and survivin protein respectively) and play an important role in the transformation of neuroblastoma [9].
The MYC family gene expression is mostly associated with the proliferation of cell populations. N-Myc was discovered as a proto-oncogene, but due to genetic alteration, this gene transforms into an oncogene. A part of the Myc family of proto-oncogenes, N-Myc is essential for neurogenesis and thus essential for the development of the nervous system. N-Myc proteins (MYCN) are included in cellular processes such as neurogenesis, cell growth, apoptosis, differentiation and the repression of genes that block cellular proliferation [10]. N-Myc also stimulates the expression of genes that control cell size and growth, such as those that encode ribosomal proteins, translation factors, and metabolic enzymes [11]. Amplification of the MCYN gene is considered as a bad prognosis in human neuroblastoma [12].
Survivin is a member of the IAP (Inhibitor of Apoptosis Protein) family genes, expressed in most of the cancer cells but not in normal cells. It is highly expressed in the S and G2M phases of the cell cycle [13]. Its expression levels correlate with more aggressive disease and poor clinical outcome. There is a high level of survivin protein expression in unfavorable groups (Stage 3 & 4) of neuroblastoma and low level of expression in favorable groups (Stage 1, 2 & 4S) [3].
This study wishes to investigate the effect of quadruplex forming DNA/Oligonucleotides at the promoter region of the survivin gene on childhood common cancer neuroblastoma. It is asked if the Guanine-Rich Oligonucleotide (GRO) sequence at the promoter regions of oncogenes, survivin oligonucleotide, forms a quadruple helix rather than a double helix and whether there is a response in neuronal differentiation and growth.
Materials and Methods
Identification of Quadruple
The QGRS program from http://bioinformatics.ramapo. edu/QGRS/analyze.php web page was used. The promoter region of the survivin gene were identified and copied into the QGRS program to identify several putative G-rich oligonucleotides (G-quadruplex) and one sequence with a high probability (high G score) was chosen to perform our study. The oligonucleotides extrapolated from the survivin gene promoter region named SVN-23. An artificially mutated version of the oligos, mSVN-23, was used as an internal control.
Circular Dichroism Spectroscopy
SVN-23 and mSVN-23 oligonucleotides were annealed by boiling for 5 minutes, adding a physiological buffer to final concentrations of 20 mmol/L KH2PO4 dibasic, 120 mmol/L KCl, 5 mmol/L MgCl2, and slowly cooled down to room temperature. Annealed ODNs (Optical Density Nucleotides) were dissolved in a physiologic buffer at a concentration giving an absorbance at 260 nm of 0.8 (about 5 μmol/L). Structures of oligonucleotides in solution were determined by circular dichroism spectroscopy. Spectra was recorded on a Jasco-810 spectropolarimeter (Jasco), using a quartz cell of 1 mm optical path length, an instrument scanning speed of 200 nm/min, the response time of 2 seconds, and over a wavelength range of 340 to 220 nm. The spectra were representative of 3 average scans taken at 25°C and was baseline corrected signal contributions due to buffer.
Cell Culture
The human neuroblastoma cell line SH-SY5Y (CRL- 2266), SK-N-AS, SK-N-SH and SK-N-BE were obtained from ATCC (Manassas, VA, United States) and maintained in DMEM supplemented with 10% FBS, Penicillin Streptomycin (100 U/ml; 100 mg/ml), and 2mM L-glutamine at 37°C in a humidified atmosphere containing 5% CO2/95% air. Semi- confluent culture was washed with phosphate-buffered saline (PBS), detached with 0.25% trypsin-EDTA solution, reseeded as 1 × 105 cells/ml of DMEM containing 10% FBS or 1% FBS or 1% BSA and used for differentiation study after overnight incubation. neuroblastoma cells were incubated with the absence or presence of G-quadruplex Oligonucleotides for the appropriate duration for the study.
Cell Differentiation and/or Cell Death Imaging
Neuroblastoma cells SH-SY5Y and SK-N-SH cells were plated at 300 cells per 60 mm tissue culture dish. The following day, different concentrations of SVN-23 (0, 2.5, 5, 7.5 and 10 μM) were added directly to the media. The cells were incubated in the presence of the oligonucleotides for 5 days. Cells will then be harvested and the dead cells and live cells with trypan blue exclusion were counted every day by light microscopy. Before counting, the cells were photographed for neurite extension, which represents the cellular differentiation. Each study was repeated, and the data was validated.
Western Blot Analysis
After being treated with indicated concentrations of SVN-23 oligo or mSVN-23 for 4 days, the cells were harvested by scraping with media, then pelletized and washed twice with PBS. Then, the cells were exposed to RIPA buffer (25 mM Tris–HCl (pH 7.6), 150 mM NaCl, 1% Nonidet P-40, 0.251% sodium deoxycholate, 1% sodium dodecyl sulfate (SDS): Thermo Fisher Scientific, United States) supplemented with protease and phosphatase inhibitors cocktail (Thermo Fisher Scientific, United States) and incubated for 30 min on ice. The lysates were centrifuged at 13,000 rpm for 20 min at 4°C, and the supernatants were collected as total cell lysate. Protein concentration was determined by the BCA method (Kit). Proteins (30 μg) were separated into 6–12% SDS-polyacrylamide gels and transferred to PVDF membranes (Millipore, Bellerica, MA, United States). The membrane was washed with Tris-buffered saline (TBS; 10 mM Tris–HCl, 150 mM NaCl, pH 7.5) containing 0.5% (v/v) Tween 20 (TBST) followed by blocking with 5% (v/v) non-fat dried milk solution prepared in TBST and then incubated overnight with primary antibodies at 4°C. The antibodies used were survivin, Neuron Specific Enolase (NSE). After this, the membrane was exposed to secondary antibodies conjugated to horseradish peroxidase for 2∼3 h at room temperature and further washed three times with TBST. Immunoreactivity was detected by the luminol- based chemiluminescence (ECL) system. Equal protein loading was assessed by the expression level of β-actin. The densitometric analysis was performed using ImageJ (National Institute of Health, Bethesda, MD, United States) software.
MTT Analysis
The cells were seeded into 96-well plates (Corning, Lowell, MA, USA) at 5000 Cells/well. Twenty-four hours later, the cells were seeded in different concentrations of SVN-23 and a control oligo mSVN-23 dissolved in water was added in the medium directly for an additional 96 hours. MTT dye (5 mg/mL, Sigma, Saint Louis, MO, USA) was added to each well. After four-hour addition of MTT reagent cells were lysed (10% SDS in 0.1N HCl) by a lysis buffer and the optical density was measured at 570 nm on a multi-well plate reader. Background absorbance of the medium in the absence of cells were subtracted. All samples were assayed in triplicate, and the mean for each study was calculated. Results were expressed as a percentage of control, which was considered to be 100%.
Statistical Analysis
Data was expressed as mean ± standard error of the mean (SEM). The significance level of treatment effects was determined by MTT assay using a Coefficient-one-tailed test followed by the student’s t-test.
Results
Identification of G-Quadruplex in Survivin Gene Promoter
Quadfinder: (http://bioinformatics.ramapo.edu/QGRS/
index.php) As shown in Table 1, there are many G-quadruplex sequences present in the survivin promoter region. The sequence with the highest G-Score (59) was selected to investigate in the study (SVN-23). For the controls, G-nucleotides were randomly selected, replacing them with non-guanine substitutes (cytosine, adenine, or thymine) and named as mSVN-23.
| Position | Length | QGRS | G-score |
|---|---|---|---|
| 99 | 28 | GGACTGTGTGGGGGTCGGGGCACACAGG | 20 |
| 235 | 30 | GGTCCTAACTTACACCTGGATGCTGTGGGG | 6 |
| 281 | 26 | GGCTCGGGAGGATGCTGGGGGCCCGG | 19 |
| 346 | 29 | GGGTGTTCAGGTCCAGGTGGACACCTGGG | 21 |
| 421 | 22 | GGAGTTAGGCCGGGCATGCTGG | 18 |
| 468 | 16 | GGGAGGCTGAGGCGGG | 19 |
| 568 | 14 | GGCATGGTGGCGGG | 19 |
| 602 | 13 | GGGGGGGCTGAGG | 17 |
| 627 | 19 | GGAACCCAGGAGATGGAGG | 16 |
| 780 | 29 | GGTTGCCAGTGGAGGTGGTGCGTCCTTGG | 16 |
| 836 | 24 | GGCCATGGTGGTGCCAGGGCCCGG | 18 |
| 1107 | 28 | GGAGAGAGACTAGGGGAGAGAAAGTAGG | 11 |
| 1137 | 29 | GGGGAGAGGGAGAGAGAAAGGAAAACAGA | 18 |
| 1181 | 21 | GGGGAGTAGAGAGAGGGAAGG | 11 |
| 1202 | 29 | GGGGAGAGGGAGAGAGAAAGGAAAACAGG | 21 |
| 1395 | 16 | GGGAGGCCGAGGCAGG | 19 |
| 1530 | 16 | GGGAGGCTGAGGCAGG | 19 |
| 1564 | 13 | GGCGGGGAGGAGG | 20 |
| 1687 | 24 | GGGGGCGGGGTGGGAGGGGTGGGG | 59 |
| 1776 | 30 | GGCCTGCGTGGGGCTGGAGGGCTAATAAGG | 20 |
Table 1: The promoter of survivin gene was analyzed through QGRS program and the putative G-rich oligonucleotides (G-quadruplex)
Selected G-quadruplex sequence for study: 1. SVN-23: 5’ -GGGG CGGGG TGGG AGGGG TGGGG -3’ 2. mSVN-23: 5’ -GAGC CAGTG TGGG ACTGG TTTGG -3’
SVN-23 Nucleotide Sequence but not Mutant mSVN-23 Forms Parallel Guanine-Quadruplex
Next, it was investigated whether these selected oligonucleotide forms the Quadruplex conformation in aqueous solution. The secondary structures of SVN-23 and mSVN-23 were studied using a circular dichroism spectrophotometer and ODNs (Optical Density Nucleotides) were analyzed by CD spectroscopy. SVN-23 formed a quadruplex in physiological buffer, represented by a peak absorbance at 260 nm and a trough absorbance at 240 nm (Figure 1). The mutant mSVN-23 sequence did not form a quadruplex in solution as they did not form a peak at 260 nm.
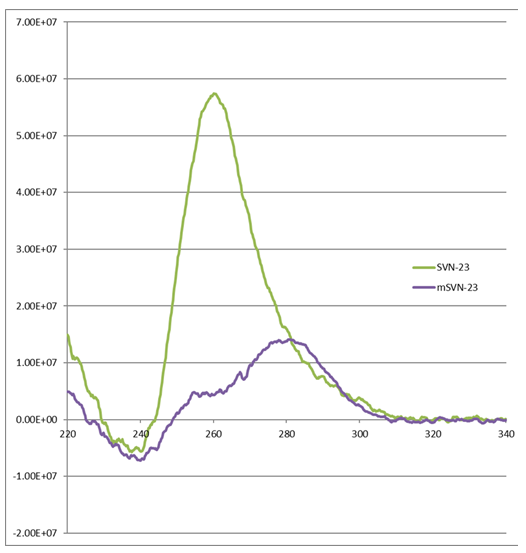
Figure1: Circular Dichroism (CD) Spectroscopy: SVN-23 not the mSVN-23 oligonucleotides represented by a peak absorbance at 260 nm form a quadruplex in solution.
Quadruplex Forming Oligo SVN-23 Induces Cell Differentiation and Cell Death in Human Neuroblastoma
The effect of the quadruplex forming oligonucleotides on different biological properties of neuroblastoma cell line SK- N-SH and SH-SY5Y were tested. SK-N-SH is a neuroblastoma cell line that displays predominantly epithelial morphology and grows in adherent culture. (Figure 2). Treatment with 13 cis-retinoic acid (used as a control) caused these cells to differentiate and adopt a neuronal phenotype, characterized by flattening of cells. SH-SY5Y cells are derived from SK-N-SH neuroblastoma cells and are neuronal subtype.
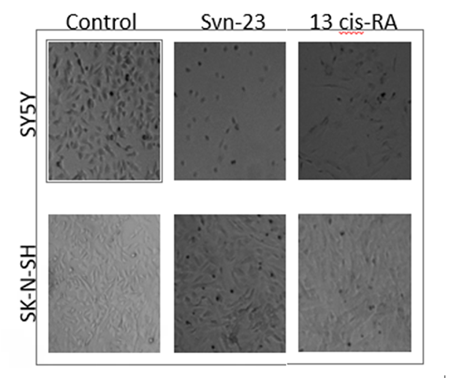
Figure 2: SH-SY5Y and SK-N-SH neuroblastoma cells were treated with 10 µM of SVN-23 oligonucleotides and 13-cis-RA for 5 days. SK-N-SH neuroblastoma cells became flattening (Lower-middle) and SH-SY5Y cells extended their neurites (Upper- middle), both of which represented the neuronal differentiation. 13-cis Retinoic Acid (right panel), a known differentiating agent was used as a positive control.
Oligo SVN-23 induced cellular differentiation by flattening the cells (Figure 2). 13-cis retinoic acid (right panel) was used as a positive control. SH-SY5Y cells, which are derived from SK-N-SH and retain only neuronal properties, induce differentiation by extending their neurites followed by cell death when treated with 10 μM of SVN-23 oligonucleotides (Figure 2) for 5 days.
MTT Assay Revealed SVN-23 Oligonucleotide Induce Growth Inhibition in Neuroblastoma Cells
To substantiate the above morphological changes observed in SH-SY5Y and SK-N-AS cells by SVN-23 an, the potential growth inhibition by MTT assay was investigated.
The MTT assay represents the mitochondrial activity of the cells that is corresponding to the number of cells eventually indicates the growth of the cellular population. SK-N-AS and SH-SY5Y neuroblastoma cells have one copy of the MYCN gene and favorable biology, on the other hand, SK-N- BE2 neuroblastoma cell has multiple copies of MYCN gene (n-MYC amplified) and carries unfavorable biology [14]. SVN-23 oligonucleotides had limited growth inhibition in both SH-SY5Y & SK-N-AS cells; on the other hand, it showed significant growth inhibition in SK-N-BE2 cells, (SVN-23, 20- 40% vs 75%) (Figure 3). This data indicated that SVN-23 oligonucleotides more effectively inhibit cellular growth in the SK-N-BE2 neuroblastoma cells where the MYCN gene is amplified.
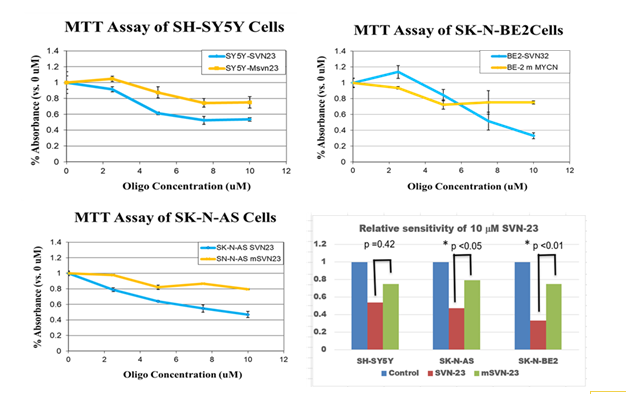
Figure 3: MTT assay of SH-SY5Y, SK-N-AS and SK-N-BE2 neuroblastoma cells. The cells were plated into 96-well plates, treated with different concentrations of oligonucleotides and assay was done on day 5. The cell sensitivity exerted by SVN-23 and mSVN-23 oligonucleotides in SH-SY5Y (upper-left), in SK-N-AS (lower left), and in SK-N-BE2 cells, (upper-right) were shown. Relative sensitivity of SVN-23 and mSVN-23 were shown (lower-right) with p<0.05 in both SK-N-AS and SK-N-BE neuroblastoma cells.
Survivin Gene Expression was Down-Regulated by SVN-23 Oligonucleotides
Multiple levels of evidence suggested that the G-quadruplex in a given gene’s promoter region inhibits the transcription machinery by strand invasion leading to downregulation of protein expression of that gene [15]. So, next it was investigated whether SVN-23 quadruplex has the ability to down-regulate the expression of the respective gene expression. Figure 4 (upper left) shows the bands of protein expression in SK-N-AS cells treated with SVN-23, mSVN-23, oligonucleotides. The first row of bands represents survivin expression while the second row of bands represents β-Actin expression (control). The relative expression of survivin in SK-N-AS cells (Figure 4) when treated with SVN-23 and mSVN-23 oligonucleotides, down regulated by approximately 67% and 30%, respectively. Figure 4 (lower right) shows the relative expression of survivin in SH-SY5Y cells. When treated with SVN-23 and mSVN-23 oligonucleotides, the expression of survivin was down regulated by approximately 39% and 15%, respectively.
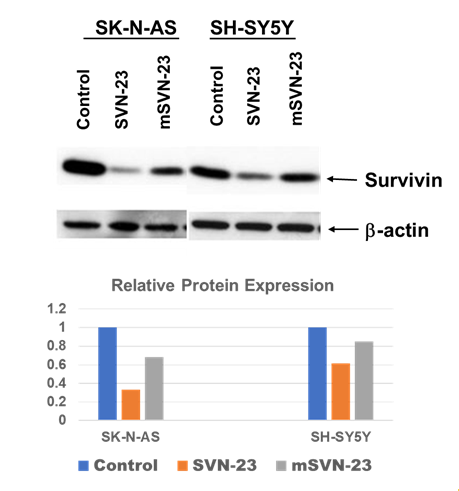
Figure 4: SK-N-AS and SH-SY5Y neuroblastoma cells were treated with 10μm SVN-23 and mSVN-23 oligonucleotides and subjected to Western blotting with antibody against Survivin and β-actin. Survivin expression was downregulated in both cell lines when treated with SVN-23 oligonucleotide (lane 2 & 5 from left, upper-panel). Relative survivin protein expression normalized with β-actin were shown (lower-panel).
Discussion and Conclusion
Purpose of Study: G-rich Quadruplex forming DNA sequence are present throughout the genome specially telomeric region and promoter region of certain genes. The quadruplex forming sequence in the promoter region of certain oncogene has also been shown to slow growth of several cancer cells [16]. The purpose of this study is to test whether the G-quadruplex forming sequences in the promoter regions of the survivin gene can be used to target the unfavorable biological groups of embryonal childhood tumor, neuroblastoma [17]. Certainly, our observations/ results in this study rationalize the potential treatment of human neuroblastoma with this putative G-quadruplex sequence, SVN-23.
Major Findings: For the first time, we were able to identify and validate the structural presence of the G-rich DNA sequences in the promoter region of the survivin oncogene and they formed quadruplex as evident by CD analysis. Human neuroblastoma cell lines were treated (SK-N-SH, SH- SY5Y, SK-N-AS, SK-N-BE2) with SVN-23. The unique results for these neuroblastoma cells are (a) SH-SY5Y (neuronal type) cells derived from SK-N-SH neuroblastoma cells mixed (epithelial & neuronal) type cells and retain only neuroblast cells [14]. It was shown for the first time that SVN-23 G-rich oligo induced differentiation in SK-N-SH by flattening the cell morphology and SH-SY5Y cells by extending the neurites.
Ultimately, these cells underwent growth arrest and perhaps cellular death as well (50 % sensitive, by MTT analysis) (b) SK-N-AS cells, which carries a single copy of N-Myc gene are modestly sensitive to both SVN-23 (50%) and (c) SK- N-BE2 cells, which carries multiple copies of N-Myc genes (N-Myc amplified) and highly sensitive to SVN-23. Lastly, it has been shown that the survivin protein expression is substantially reduced when treating the SH-SY5Y & SK-N-AS neuroblastoma cells with SVN-23, though down regulation is much more in SK-N-AS cells.
Support of Research hypothesis by data: Our data clearly supported the first part of our working hypothesis. We have shown clearly that G-rich sequences in the promoter region of the survivin gene changed their normal canonical conformation, double-helix to non-canonical quadruplex conformation (Figure 1). It has also been evidently proven in our second part of the hypothesis that there were neuronal differentiation and/or cell death by flattening of cells (SN- N-SH) and extension of the neurites followed by cellular morphology suggestive of cell death (SH-SY5Y) (Figures 2 & 4). This differentiation and or cell death of neuroblastoma cells were validated by MTT assay which represents a smaller number of cells in SVN-23 treated cells.
Comparison with other research: The concept of targeting G-quadruplex as a therapeutic strategy was first developed for telomeric DNA and telomerase inhibition.
Studies of several independent laboratories aimed at understanding the repeats in telomere have shown that neuroblastoma with high telomerase activity had a poor prognosis unfavorable biology and advanced stage of neuroblastoma [18]. Amplification of MYCN is associated with adverse outcome in neuroblastoma, although MYCN expression without multiplication is shown to be beneficial [19]. Down-regulation of survivin gene expression has been shown to induce neuronal differentiation in neuroblastoma [3]. Together, these findings indicate that the n-Myc & survivin gene signaling pathways are important in maintaining an undifferentiated phenotype and that the inhibition of n-MYC and survivin gene could contribute to less aggressive neuroblastoma.
Explanation of Findings: A mutated version of G-rich oligonucleotides m-SVN-23 (Table 1) has been treated by randomly replacing most of the guanine with non- guanine oligos to use as an internal control. m-SVN-23 did not form Quadruplex as evident by CD analysis (Figure 1). Interestingly when the neuroblastoma cells were treated with these control oligos, a certain level (~20%) of sensitivity in neuroblastoma cells was observed as well: SVN-23 by MTT analysis (Figure 3). One of the explanations could be that these mutant oligoes serve as an antisense or SiRNA of other prominent genes important for regulation of neuroblastoma biology [20]. The expression of the survivin protein is modestly (~70%) reduced in SK-N-AS and ~40% in SH-SY5Y cells when treated with SVN-23 oligos; however, survivin expression is not changed when targeting the MYCN gene with another G-rich Oligo: MYCN-15 (data not shown). Considering the fact that both the N-Myc and survivin genes are important in regulating neuroblastoma biology, our finding suggests that survivin might work upstream of n-Myc gene to regulate the neuroblastoma biology.
Recommendation for future research: Our study as well as other investigator has shown that the survivin gene is highly expressed in SK-N-AS, SH-SY5Y and SK-N-BE neuroblastoma cells [21]. This study shows that when SVN-23 G-quadruplex oligonucleotide is used, there is a down regulation of survivin in both SK-N-AS & SH-SY5Y cells, a potential beneficial aspect of this product since there is high expression of survivin in neuroblastoma. However, both SK-NAS & SH-SY5Y cells have only one copy of n-Myc gene (non-amplified), and it would be interesting to see in future the expression pattern of survivin protein in an n-Myc amplified neuroblastoma cell line like SK-N-BE2 treated with SVN-23. Our MTT assay indicated SK- N-BE2 cells are more sensitive (>80%) (Figure 3), so it can be assumed that there was complete/ near complete down regulation of survivin protein expression in SVN-23 treated SK-N-BE cells. It has been shown that several G-rich oligos can enter the cells efficiently [22]. As we have identified this unique oligos, it will be conjugated with fluorescent (GFP/ RFP) and the neuroblastoma cells will be treated to validate their entrance into the cells in future.
It is concluded that G-quadruplex forming sequences in the survivin promoter region could be a potential target for the treatment of common childhood solid tumor, neuroblastoma, specially with unfavorable biology.
References
-
Nakagawara A, Li Y, Izumi H, Muramori K, Inada H, et al. (2018) Neuroblastoma. Jpn J Clin Oncol 48(3): 214-241.
-
Brodeur GM, Bagatell R (2014) Mechanisms of neuroblastoma regression. Nat Rev Clin Oncol 11(12): 704-713.
-
Islam A, Kageyama H, Takada N, Kawamoto T, Takayasu H, et al. (2000) High expression of Survivin, mapped to 17q25, is significantly associated with poor prognostic factors and promotes cell survival in human neuroblastoma. Oncogene 19(5): 617-623.
-
Bordow SB, Norris MD, Haber PS, Marshall GM, Haber M (1998) Prognostic significance of MYCN oncogene expression in childhood neuroblastoma. J Clin Oncol 16(10): 3286-3294.
-
Brodeur GM (2018) Spontaneous regression of neuroblastoma. Cell Tissue Res 372(2): 277-286.
-
Paeschke K, McDonald KR, Zakian VA (2010) Telomeres: Structures in need of unwinding. FEBS Lett 584(17): 3760-3772.
-
Bates PJ, Laber DA, Miller DM, Thomas SD, Trent JO (2009) Discovery and development of the G-rich oligonucleotide AS1411 as a novel treatment for cancer. Exp Mol Pathol 86(3): 151-164.
-
Dever TE, Dinman JD, Green R (2019) Translation Elongation and Recoding in Eukaryotes Overview of Translation Elongation HHS Public Access. Cold Spring Harb Perspect Biol 10(8): a032649.
-
Azuhata T, Scott D, Takamizawa S, Wen J, Davidoff A, et al. (2001) The inhibitor of apoptosis protein survivin is associated with high-risk behavior of neuroblastoma. J Pediatr Surg 36(12): 1785-1791.
-
Lam WA, Cao L, Umesh V, Keung AJ, Sen S, et al. (2010) Extracellular matrix rigidity modulates neuroblastoma cell differentiation and N-myc expression. Mol Cancer 9: 35.
-
Knoepfler PS, Cheng PF, Eisenman RN (2002) N-myc is essential during neurogenesis for the rapid expansion of progenitor cell populations and the inhibition of neuronal differentiation. Genes Dev 16: 2699-2712.
-
Dzieran J, Garcia AR, Westermark UK, Henley AB, Sánchez EE, et al. (2018) MYCN-amplified neuroblastoma maintains an aggressive and undifferentiated phenotype by deregulation of estrogen and NGF signaling. Proc Natl Acad Sci U S A 115(6): E1229-E1238.
-
Ambrosini G, Adida C, Altieri DC (1997) A novel anti- apoptosis gene, survivin, expressed in cancer and lymphoma. Nat Med 3(8): 917-921.
-
Thiele CJ (1998) Neuroblastoma Cell Lines. In: Masters JRW, et al. (Eds.), Human Cell Culture. Kluwer Academic Publishers 1: 21-53.
-
Smaali NT, Guittat L, Sidibe A, Ya KS, Trentesaux C, et al. (2009) The G-quadruplex ligand telomestatin impairs binding of topoisomerase IIIα to G-quadruplex-forming oligonucleotides and uncaps telomeres in ALT cells. PLoS One 4(9): e6919.
-
Teng Y, Girvan AC, Casson LK, Pierce WM, Qian M, et al. (2007) AS1411 alters the localization of a complex containing protein arginine methyltransferase 5 and nucleolin. Cancer Res 67(21): 10491-10500.
-
Shalaby T, Fiaschetti G, Nagasawa K, Ya KS, Baumgartner M, et al. (2013) G-quadruplexes as potential therapeutic targets for embryonal tumors. Molecules 18(10): 12500- 12537.
-
Hiyama E, Hiyama K, Ohtsu K, Yamaoka H, Ichikawa T, et al. (1997) Telomerase activity in neuroblastoma: Is it a prognostic indicator of clinical behaviour?. European Journal of Cancer 33(12): 1932-1936.
-
Huang M, Weiss WA (2013) Neuroblastoma and MYCN. Cold Spring Harb Perspect Med 3(10): a014415.
-
Zamore PD (2006) RNA Interference: Big Applause for Silencing in Stockholm. Cell 127(6): 1083-1086.
-
Hossain MM, Banik NL, Ray SK (2012) Survivin knockdown increased anti-cancer effects of (-)-epigallocatechin-3- gallate in human malignant neuroblastoma SK-N-BE2 and SH-SY5Y cells. Exp Cell Res 318(13): 1597-1610.
-
Islam MA, Thomas SD, Murty VV, Sedoris KJ, Miller DM (2014) c-Myc quadruplex-forming sequence Pu- 27 induces extensive damage in both telomeric and nontelomeric regions of DNA. J Biol Chem 289(12): 8521-8531.
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis
- Nutritional, Therapeutic, and Environmental Effect of Oyster Mushrooms: An Editorial