Mechanism and Physiological Relevance of Xanthine Oxidase as Nitrate Reductase in Mammalian Organism: An Overview
In living organisms nitric oxide radical is produced by the enzymes NO synthases, with consequent oxidation to nitrate by several oxidation/reduction pathways. The end step of NO oxidation is the formation of nitrite and nitrate, which after their formation enter a reduction phase of NO cycle, thereby playing a physiologically important role of NO donors. However, while in plants and microorganisms the key pathway of nitrate/nitrite reduction is the functioning of the enzymes nitrate reductases (NRs), mammalian organism lacks NRs enzymes. Instead, in mammals, the reduction of nitrates and nitrites occurs by several other, both enzymatic and non-enzymatic, pathways. Based on the present knowledge, one of the key enzymes involved in the nitrate/nitrite reduction process is xanthine oxidoreductase (XOR). XOR is capable of reducing organic and inorganic nitrates and nitrite, and the role of XOR in NO3- → NO2- → ·NO pathway is most studied as compared to other enzymes with nitrate reductase activity. The functions of XOR, the mechanisms of nitrate/nitrite reduction by this enzyme and their physiological relevance will be outlined in this mini review.
Introduction
Nitric oxide radical (·NO) is a vital molecule for all living organisms. It is produced by the ubiquitously present enzymes NO synthases as the result of the conversion of its substrate L-arginine to L-cytrulline, which requires the presence of oxygen and the cofactors: FAD, FMN, NADPH, and tetrahydrobiopterin (reviewed in detail in several reviews [1, 2]). In the course of multiple redox reactions resulting in the formation of several reactive nitrogen species (RNS) both ‘beneficial’, such as nitrosothiols (RSNO), and harmful as peroxynitrite (ONOO-), NO is oxidized, and the end step of the oxidation process is the formation of nitrate, NO3 -.
Toxicity of nitrates for mammals’ physiology is without doubts [3]. However, last decades of research, starting from 90th of XX century, showed great potential of nitrates, and especially nitrites, as the donors of NO, which is produced by nitrate/nitrite reduction by several both enzymatic and non-enzymatic pathways. Nitrate/nitrite reduction to NO becomes especially important under physiological and pathophysiological conditions characterized by the oxygen shortage and the down-regulation of constitutive NO biosynthesis by NO synthases (ischemia, hypoxia [4, 5], hypertension [6], heart failure, diabetes [7], and ageing [8]. The beneficial effects of dietary nitrate and nitrate based therapy on ischemia-reperfusion, hypertension, renal function, and tolerance to hypoxia were observed and reviewed [6, 9, 10, 11]. Meanwhile, there is still insufficient knowledge of the numerous pathways of NO3 -/NO2 - reduction in mammals and their physiological role.
It is well known that in plants and microorganisms NO3 - reduction is accomplished by the enzymes nitrate reductases, which several isoforms and functions were extensively described in the literature [12, 13]. It worth notion, that these enzymes capable of NO3- reduction can reduce nitrite as well. In mammals, there are multiple enzymatic and non- enzymatic pathways of nitrate/nitrite reduction to NO; of these pathways ability of xanthine oxidoreductase to reduce nitrate and nitrite to NO is most studied one. By its structure and the mechanism, mammalian xanthine oxidoreductase is similar to nitrate reductases. There we will focus on the mechanism and physiological relevance of XOR functioning as nitrate reductase in mammals based on published research.
The Key Pathways of Nitrate/Nitrite Reduction in Mammals
According to the present knowledge, mammals lack NRs enzymes [14]. Instead, nitrate and nitrite in mammals’ organism are reduced by several other, both enzymatic and non-enzymatic, pathways. Nitrate reducing pathways presently known include enzyme xanthine oxidoreductase capable of reducing both inorganic and organic nitrates and nitrite [14], and microbiota present in mammals’ organism, which possess several nitrate and nitrite reductase activities [15, 16, 17]. Important function in reducing organic nitrates also belongs to mitochondrial aldehyde dehydrogenase 2 (ALDH2), which catalyzes the conversion of the well-known cardioprotective drug nitroglycerine and other organic nitrates to NO [3]. Down-regulation of this enzyme is implicated in a well-known phenomenon of ‘nitrate tolerance’, which greatly reduces therapeutic effects of nitroglycerine in cardiovascular diseases. So, ALDH2, which primary function is the reduction of aldehydes to related acids, is considered as cardioprotective enzyme because of its nitrate reductase activity [3].
There are still more pathways of nitrite reduction presently known in mammalian organism. On a whole, the function of reducing nitrite belongs to all molybdopterin- containing flavoenzymes presently known in mammals: xanthine oxidoreductase (XOR), aldehyde oxidase (AOX), sulfite oxidase (SO), and mitochondrial amidoxime reducing component (mARC protein 1 and 2 isoforms found in humans) [14]. All these enzymes, especially AOX have much similar with XOR in their chemical structure and the mechanism of nitrite reduction [18]. However, besides XOR, there were no reports of nitrate reduction by SO, AOX, and mARC, so these enzymes rather act as nitrite reductases, while their possible nitrate reductase activity remains unexplored. Besides of Mo-containing flavoenzymes, the ability of NO2 - reduction was shown for several metalloproteins (Fe, Cu) and heme-containing proteins, including hemoglobin, myoglobin, cytochrome c, cytochrome P450, cytochrome c oxidase, and many others [14, 18]. Mitochondrial electron transport chain, which Complexes I, III, and IV can reduce nitrite to NO dependent on energy state [19, 20], too is involved in NO production. Interestingly, that nitrate/ nitrite reductase activity is not a primary function of the enzymes and proteins known to possess such ability, so that these enzymes were designated as ‘non-dedicated nitrite reductases’ [18]. However, in this short review we will focus on the functioning of xanthine oxidase as the most studied of the enzymatic nitrate-nitrite-NO pathways.
Of presently known enzymatic pathways of nitrate reduction in mammals, XOR needs to be mentioned in the first place as most studied multi-functional enzyme with well described physiological functions. Attention of the researchers to XOR can be explained by its important physiological role in purine catabolism and conversion of hypoxanthine and xanthine to uric acid, as well as the involvement in the development of several pathophysiological conditions, because of its known ability to produce superoxide and hydroperoxide [21, 22, 23]. However, while there were several works and reviews devoted to the mechanism of xanthine oxidation and superoxide production by XOR [22, 24, 25], less attention was paid to the nitrate and nitrite reductase activity of this enzyme.
The Structure and Multiple Finctions of Xanthine Oxidoreductase
The studies on xanthine oxidoreductase (XOR) count more than a 100-years’ history. For several decades this enzyme, responsible for the oxidation of hypoxanthine to xanthine and finally, to uric acid, which are the final steps of purine catabolism, was implicated in the development of multiple pathophysiological processes and diseases accompanied by ROS overproduction [4, 21, 23, 26]. A positive correlation was found between the levels of uric acid and the development of cardiovascular diseases: hypertension, atherosclerosis, atrial fibrillation, heart failure [23]. However, the role of XOR function as NR in physiology still remains a little explored field of research. The mechanism of enzymatic activity of XOR was well studied over decades.
Molecular architecture and functions of xanthine oxidase: Common to plant and bacteria nitrate reductases, XOR is a Mo-containing flavoenzyme, which consists of two identical subunits each with molecular mass ~150 kDa [24]. Crystal structure of XOR at the resolution of 2.5 and later 1.5 Å was first obtained by Enroth C and Nishino T [24, 27]. Each subunit consists of a peptide chain of ~1330 amino acids that binds one Mo-pterin cofactor, two 2Fe-2S centers, and a FAD cofactor (Figure 1); Mo center (Mo-cofactor) is localized at each of 85 kDa C-terminal domains; FeS clusters are localized in a small 20 kDa N-terminal domains connected to intermediate 40 kDa domains, which contain a FAD binding site [24]. Three domains of each subunit are connected by extended amino acid linker segments [24]. So called Mo-cofactor is a Mo-pterin complex that is an active site of XOR enzyme, where xanthine oxidation to uric acid occurs by hydroxylation reactions [24, 18]. Meanwhile, XOR is a multi-functional enzyme that, dependent on conditions, can produce superoxide, which is well documented in several pathophysiological processes [4, 21, 28], and function as nitrate/nitrite reductase [14, 18].
Xanthine oxidoreductase exists in two forms: reduced xanthine dehydrogenase (XDH, EC 1.171.4) and oxidized one, xanthine oxidase (XO, EC 1.173.2), of which XDH prevails under normal physiological conditions [18]. XDH can be converted to XO either irreversibly by limited proteolysis, or reversibly, by oxidation of critical SH groups with the formation of disulfide bonds; both mechanisms are relevant under oxidative stress conditions [22, 24]. Limited proteolysis at Lys551 or Lys569 (bovine XOR), which results in irreversible conversion of XDH to XO can occur under ischemic conditions accompanied by the increase in the levels of intracellular Ca2+ and the activation of a protease calpain [22]. Reversible conversion of XDH to XO needs a formation of a disulfide bond between two Cys residues (Cys535 and Cys992 in bovine XOR) at FAD site [24, 29]. Enzymes’ ability to produce superoxide dramatically increases upon transition of XDH to XO.
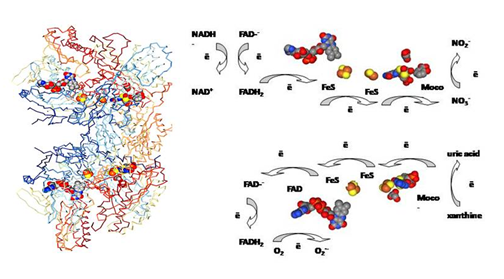
Figure 1: Crystal structure of xanthine oxidase (PDB file 1FIQ) and schematic representation of oxidase and nitrate reductase activity of XO. Redox sites and their orientation correspond to the crystal structure of XO. The structure is obtained from PDB. Source: Enroth, C., Eger, B.T., Okamoto, K., Nishino, T., Nishino, T., Pai, E.F. Crystal structure of xanthine oxidase from bovine milk. (2000) Proc Natl Acad Sci U S A 97: 10723-10728. PDB 1FIQ.
Both XDH and XO are identical in the mechanism of their reactions with substrates at active Mo center [18], which primary function is to accomplish oxidation of hypoxanthine to xanthine and finally, to uric acid (detailed mechanism reviewed in Refs [18, 22]. This hydroxylation reaction occurs at molybdenum center of the enzyme [18]. In oxidized XOR, molybdenum is in a maximally oxidized state Mo+6 surrounded by =O, =S, -OH, and two vicinal S ligands connected to aromatic ring of pterine [18, 24]. Hydroxylation is initiated by the activation of Mo+6-OH ligand by neighbor deprotonated glutamate residue Glu1261. In the reaction with xanthine a bridge is formed between Mo–OH and C atom of the aromatic ring of xanthine resulting in hydroxylated xanthine intermediate followed by H transfer from the substrate to S atom and the reduction of Mo: Mo+6=S → Mo+4- SH [18]. The uric acid formation, as showed kinetic studies, is a two-step process, which results in the reduction of Mo: Mo+6 → Mo+5 → Mo+4 [22, 24]. Summary reactions in XDH (1) and XO (2) are:
Xanthine + H2O + NAD+ → urate + NADH + H+ (1) Xanthine + H2O + O2 → urate + *O2 - + H2O2 (2)
Upon xanthine oxidation, electrons via two FeS clusters are tunneled to FAD site (Figure 1), and Mo center became reoxidized to Mo+6. At FAD site, XOR can be either oxidized by NAD+ to form NADH (1), or react with O2 to form superoxide and hydroperoxide (2).
Main difference between XDH and XO are the reactions occurring at FAD site [24, 27]. Of two forms of XOR, reduced XDH readily reduces NAD+ to NADH, whereas XO is unable to reduce NAD+ and reacts with O2 reducing it to hydroperoxide and superoxide [24, 25]. There is a thermodynamic and structural background, which explains such behavior. As it was shown, upon the conversion of XDH to XO a 11-residue loop Gln423-Lys433 (bovine XOR) in close proximity to FAD undergoes a structural rearrangement, which changes electrostatic surrounding at FAD cofactor. In XDH, there is a strong negative charge at flavin binding site, Asp429 (bovine) in a close proximity to flavin ring (only ~ 3.1 Å) [24], which favors NAD+ binding. Electrostatic milieu is dramatically changed upon the transition to XO because of the replacement of Asp429 by Arg425: positively charged guanidine group of Arg425 becomes an obstacle for the access of NAD+ to FAD site [24, 27]. Meanwhile, these structural rearrangements do not affect reactions at Mo center [24, 27]. Structural data are in the agreement with potentiometric titrations of XDH and XO enzymes.
As NAD+ reduction takes place at the flavin binding site of the enzyme, redox state of flavins is a key determinant of this reaction. Redox titration of XDH and XO has shown differences in the redox state of flavins of these two forms of XOR (Table 1). It worth mention, that FAD- reduction to FADH2 occurs in two steps: FAD- anion reduction to neutral semiquinone form FADH*, and FADH* reduction to FADH2. There were differences found in the redox potentials of FAD-/ FADH* and FADH*/FADH2 couples in XDH and XO [30, 31].
| Enzyme | Conditions | Midpoint Potential, E0, mV | Source | |||||
|---|---|---|---|---|---|---|---|---|
| FAD-/FADH* | FADH*/FADH 2 | FeS I | FeS II | Mo-Cofactor | ||||
| MoVI/MoV | MoV/MoIV | |||||||
| XO cow milk | 25°C, pH7.2, anaerobic | -235 | -215 | -300 | -197 | -300 - | -300 - | [30] |
| -335 | -309 | |||||||
| XDH chiken liver | 25°C, pH7.5, anaerobic | -270 | -410 | -310 | -235 | -320 | [31] |
Table 1: Redox potentials of the redox active sites of xanthine dehydrogenase and xanthine oxidase, literary data [30,31].
The relative order of redox potentials in XO was put in a row: FeS(II) > FADH*/FADH2 > FAD-/FADH* > FeS(I), whereas in XDH it was different: FeS(II) > FAD/FADH* > FeS(I) > FADH*/FADH2 [31] (Table 1). So, while midpoint potential of FAD-/FADH* couple was similar in XDH and XO (-270 and -280 mV respectively), E0 for FADH*/FAD- couple was -410 in XDH and -230 in XO [31]. Considering that E0 for NAD+/NADH couple is about -335 mV, it explains the reaction of NAD+ with XDH and the inability of NAD+ reduction in XO (Table 1). The dependence of NAD+ reduction on redox potentials of oxidized/reduced FAD was shown in enzymes preparation in which native FAD was substituted by synthesized modified FAD analogs with different redox potentials: -360 and -158 [32]. Besides, there is different stability of the redox forms of FAD cofactor in XDH and XO.
As it was shown, XO much faster reacts with oxygen than XDH does [31]. Negatively charged groups in flavin binding site of XDH result in a weak binding of an FAD- anion, while a neutral semiquinone form FADH* in XDH is more stable because of its stabilization by H-bonds [24, 27]. In XO FADH* form is less stable and more reactive to oxygen; thus, in XO molecular oxygen slowly reacts with FADH* resulting in the formation of FAD- and superoxide: FADH* + O2 → O2*- + FAD [24, 33]. This reaction involves FAD binding site and FeS clusters [33]. XDH too can react with oxygen; the reaction results in the FADH2 oxidation to FAD and the formation of H2O2: FADH2 + O2 → H2O2 + FAD [24]. As it was shown [28], hydroperoxide is a main product of XO as well: 70-95% of ROS produced by XO was as H2O2; H2O2 formation increased with the decrease of oxygen concentration.
Nitrate Reductase Properties of Xanthine Oxidase
Xanthine Oxidase as a Nitrate Reductase: Ability of XOR to function as nitrate and nitrite reductase received much less attention than superoxide production, however, the nitrate reductase properties of this enzyme were studied for decades [34, 35, 36, 37, 38]. NR properties of XOR are very similar to those of plant and bacteria NR because of structural similarity in Mo cofactor of these enzymes [13]. Of mammalian molybdenum flavoenzymes, there is a close similarity between XO and AOX [18].
Nitrate reduction is a two-step process with first formation of nitrite and consequent nitrite reduction to NO [36]. These reactions take place at the Mo cofactor, independent of XDH or XO forms of the enzyme (Figure 1). As it was shown [34, 36], for this reaction XOR can accept electrons both from physiological (NADH, xanthine), and non- physiological electron donors (reduced viologen, ditionite, aldehydes). First, nitrate reduction to nitrite occurs, and then nitrite is reduced to NO [36].If electron donors acting at FAD site are involved; nitrate reduction is blocked by flavin site inhibitor diphenyleneiodonium (DPI), a blocker of FAD site [34, 35, 36]. It was already shown in early studies, that reduced methylviologen, ditionite, hypoxanthine, and NADH could be electron donors in vitro for NO3 - and NO2 - reductase activity of XOR; for physiologically relevant electron donors, NADH and hypoxanthine, FAD cofactor was strongly required for NR activity, while methylviologen and ditionite were shown to be FAD-independent ones [34].
Nitrate reduction at Mo site was proved by an effective inhibition of this reaction by XOR inhibitors allopurinol and oxypurinol, known to bind to the Mo-center (5-10 µM of allopurinol was sufficient for complete inhibition of NR activity) [34]. Oxypurinol was shown to inhibit nitrate reduction regardless of reducing substrate present. This is true for allopurinol as well, and indicates nitrate binding and reduction at Mo site. In agreement with this, DPI inhibited nitrate reduction only when NADH was used as reducing agent, and did not inhibit nitrate formation when xanthine was used [36]. This indicated that NADH donated electrons to FAD, and then electrons were transported via FeS clusters to Mo site where nitrate reduction took place (Figure 1). With xanthine or aldehydes (DBA) as electron donors, reduction occurred at Mo site, so that oxypurinol (as well as allopurinol), but not DPI, could inhibit nitrite formation. The studies performed on purified XO enzyme were confirmed by the studies of nitrate reduction and NO formation on hypoxic heart tissue [36]. Of other inhibitors of NR activity, it was shown already in early studies on XOR [34], that NR activity was inhibited by azide (10-2 M) and cyanide (10-3 M) in the presence of ē donors (methylviologen, ditionite, and NADH) [34]. Cyanide acts at Mo=S ligand of Mo center with following loss of S atom, formation of desulfo-enzyme, and XO inactivation. The enzyme can be reactivated by the treatment with sulfide and ditionite [24, 29].
While there are several works on molecular mechanism of nitrite reduction by XO ([18] and references therein), we were unable to find a study on molecular mechanism of the nitrate reduction by XOR enzymes. In a very simplified form based on the nitrate reduction by nitrate reductases with similar structure of Mo center (reviewed in [13]), we can assume that similar to the NO2- reduction mechanism, the mechanism of NO3- reduction involves Mo shuttle between +4 and +6 with an intermediate Mo+5 state [18]. NO3 - binds via 1 oxygen atom to Mo+4 center forming Mo-O-NO2 ligand;
protons are needed for N-O bond cleavage. NO2 - is released accompanied by Mo oxidation and transition to +6. Bound oxygen atom remains as a terminal oxo group, which protonation results in formation of H2O molecule coordinated to Mo+6. Regeneration of a catalytically active Mo center by substrate xanthine or other electron donors is accompanied by Mo transition to +4 state and water release.
Detailed studies on the kinetics of nitrate reduction [35, 36], have shown that of reducing agents used for XO- catalyzed nitrate reduction (xanthine, dibenzaldehyde (DBA), and NADH), NADH showed the slowest rate of NO formation. The concentration dependence of the rate of nitrite production, either on nitrate or reducing agents, (NADH, xanthine), followed Michaelis-Menten kinetics. The rate of NO formation linearly increased with the increase of the amount of nitrite formed upon nitrate reduction. Km found for nitrate reduction constituted 0.29 ± 0.6 mM, and for nitrite reduction 22.9 ± 8.1 mM [35]. Nitrite production from XO-catalyzed nitrate reduction was shown to be of zero order [36], and constant rate of nitrite formation from nitrate was observed with each substrate (NADH, DBA, or xanthine). High xanthine or nitrate levels blocked Mo site and inhibited nitrite formation [36, 37, 39]. Oxygen too was shown to inhibit nitrite reductase activity of XO [36].
Nitrite formation required anaerobic conditions and was pH-dependent with maximal rate at pH 5 [36], which agrees with molecular mechanism that involved protonation steps in NO2 - formation. This is different of xanthine oxidase activity, which exhibits optimum pH 8-9 [34, 35]. As acidosis and fall of pH to about ~6 units is typical for ischemia, this agrees with the data showing an elevation of XOR expression and activity under hypoxia and ischemia and makes an XOR physiologically important source of NO in several cardiovascular diseases [22].
Reduction of Organic Nitrates by XO: As it was shown, XO is capable of reducing organic nitrates, including commonly used nitroglycerine (glycerine trinitrate, GTN), shown under anaerobic conditions [35, 38]. Based on the study of Li H, et al. [38], the reduction of organic nitrates, the same as inorganic ones, is a two-step process, which first step is the reduction of organic nitrate to organic nitrite, R-O-NO [38]. Second step of the reduction process required the presence of thiols or ascorbate, and resulted in generation of NO or nitrosothiols. XO-mediated reduction of organic nitrates occurs by acid-catalyzed mechanism. Different of inorganic nitrate reduction, the reduction of organic nitrate occurred at flavin site, and was inhibited by DPI, and not by the inhibitors of Mo center [35, 38].Nitrite formation from GTN reduction with xanthine as substrate followed Michaelis-Menten kinetics with Vmax 17.4 nmol/s/mg and Km 0.49 mM. Consequent NO
formation required the presence of thiols, and was started by the addition of L-cystein [38]. Without thiols or ascorbate, no nitric oxide formation occurs.
Nitrite Reduction by Xanthine Oxidase: Ability of inorganic nitrite reduction is an intrinsic property of nitrate reductases. The mechanism of nitrite reduction by XO received much attention and was reviewed in details in [18] and references therein. The same as with nitrate, nitrite reduction may occur with multiple reducing substrates (heterocyclic compounds (xanthine), aldehydes, NADH) and results in *NO formation [18, 37].
2NO2 - are consumed and 2*NO radicals are formed per 1 reducing substrate (NADH, xanthine, aldehydes), which is oxidized either at FAD (NADH) or Mo center of XOR (aldehydes, xanthine). Similar to nitrate, NO2 - reduction exhibits Michaelis-Menten kinetics; the slowest rate of NO formation too was observed with NADH as reducing substrate. The same as with NO3 -, anaerobic conditions and slightly acidic pH (pH~6) are required optimum for NO2 - reduction by XOR. Kinetic characteristics for NO formation from nitrite and nitrate are similar in XDH and XO, which agrees with the mechanism of the reaction at Mo center that does not differ between two forms of XOR. Apparent Km for NO2 - was estimated to be ~1.9-4.1 mM, which far exceeds physiological levels of NO2 - (< 20 µM) [18]. This might question XOR ability to reduce nitrite in vivo, but it needs to be considered that shift to the lower pH (pH~6) results in the lowering of Km to micromolar concentration range [18].
The mechanism of nitrite reduction (reviewed in details in [18] and references therein]) ‘starts’ from the reduction of Mo to Mo+4 by any suitable reducing agent. Upon reaction with NO2- first Mo+4-O-N=O bonds are formed between reduced Mo and nitrite; following step is the protonation of the nitrite oxygen atom followed by hemolytic cleavage of O-N bond with the involvement of neighbouring Glu1261 (bovine XO), release of NO molecule and Mo oxidation to Mo+5. As the catalytic cycle is a two electron process, a second cycle of Mo oxidation/nitrite reduction takes place, again starting from a protonation step and ending by second NO molecule release and Mo oxidation to Mo+6 [18]. As nitrite reduction to NO is a one-electron process, and the catalytic cycle of XOR requires the two-electron oxidation of Mo+4 to Mo+6, formation of Mo+5 intermediate could result in XOR inactivation and arrest of catalytic cycle [18]. The same as nitrate, nitrite reduction is inhibited by oxygen, which is dependent on the concentrations of reducing substrate, oxygen, and NO2 -. Xanthine too was shown to inhibit nitrite reductase activity of XO because of competing interaction with Mo center [14].
Physiological Relevance of Nitrate Reductase Activity of Xanthine Oxidase
For many years, the study of the mechanism of XOR functioning, besides xanthine oxidation, was mainly focused on the production of superoxide, and XOR activity was implicated as one of the main sources of ROS under pathophysiological conditions and inflammatory processes [21, 23, 26]. Positive correlation was found between uric acid content and cardiovascular diseases: hypertension, atherosclerosis, heart failure [23]. It was shown that hypoxia and ischemia up-regulated expression of XOR in arterial endothelial cells [22]. What’s more, an oxidative stress conditions and rise in intracellular Ca2+ favor an irreversible conversion of XDH to XO, which explains an increase in superoxide production by this enzyme under ischemic conditions and several diseases [22]. However, it worth mention, that hypoxic conditions and acidic pH (6- 7) are most favorable for NR activity of XO as well. Besides, last decades gave many evidence of physiological relevance of nitrate reductase activity of XO under normoxia. So, XOR was shown to be important in maintaining NO homeostasis under several conditions [4, 5, 6].
Reduced bioavailability of NO is positively correlated with cardiovascular diseases [3]. So, numerous research works and reviews have shown an importance of nitrate rich diet (such as Mediterranean diet) and nitrate based therapy for treatment of cardiovascular disorders and maintenance of NO homeostasis in human organism [3, 6, 8]. Optimal daily intake of nitrates and nitrites as diet was evaluated: 0.07 mg/ kg/day nitrite and 3.7 mg/kg/day nitrate to maintain NO homeostasis in human organism [3]. In this nitrate/nitrite based therapy key role belongs to the enzymes capable of functioning as nitrate reductases: XO and mitochondrial ALDH2 [3, 4].
While most studies on nitrate/nitrite reduction by XO were conducted under anaerobic conditions, and oxygen was shown to inhibit NR activity of XOR [36, 37], presently there is much evidence in the literature showing the ability of XOR to reduce nitrate under aerobic physiological conditions [16, 40]. As it was shown in skeletal muscle, XOR function as nitrate reductase is dependent on oxygen tension, pH, and XOR isoform: XDH or XO [5]. Ability of XOR to reduce nitrite makes it a physiologically important source of NO [3, 6, 8, 16]. However, high nitrate levels can inhibit XOR nitrite reductase activity [39]. Similarly, excess xanthine inhibits NO production by XO because of binding to Mo site [36].
An increase in nitrate levels results in up-regulation of XOR enzyme. Thus, it was shown [16] that after nitrate injection plasma nitrite was 3 times higher in germ-free mice as compared to conventional ones because of XOR up-regulation; accordingly, this effect was inhibited by XOR inhibitor allopurinol. An importance of XOR activity for the regulation of nitrate/nitrite level and NO production in ageing rats too was shown [8]. Dietary inorganic nitrate was reported to be important for the regulation of blood pressure in hypertension and other cardiovascular diseases [6, 10, 11], which implies beneficial role of XO in keeping physiological levels of NO.
Down-regulation of constitutive NOS expression too was shown to be favorable for XOR enzyme up-regulation [41]. This was shown in several pathophysiological conditions marked by cNOS deficiency, such as cardiovascular disorders and diabetes [3, 41]. As we have shown recently, nitrate reductase XOR activity in heart tissue increased in a rat model of type I diabetes, which was accompanied by severe suppression of eNOS [42]. The role of XOR as an NO source based on XOR ability to reduce nitrate and nitrite was shown in diabetes [41], hypertension [6], and ageing [8] and in healthy heart tissue [16, 36, 40]. Interestingly, that NR activity of XOR was found in tissues, and not in circulation; red blood cells did not show any XOR activity [42].
The above examples show the importance of XOR functioning as nitrate reductase for normal physiology and diseases, however, the role of XOR in NO production should not be overestimated as well. At present, it is not quite clear, which form of XOR enzyme can better function as nitrate reductase under physiological conditions. It needs to be evaluated, which of the reactions: oxygen reduction to superoxide, or nitrate reduction to nitrite is more favorable under oxidative stress conditions, and if XO functioning as NR could not be involved in peroxynitrite formation. It seems reasonable to hypothesise that increased oxidative stress in several diseases could favor XOR function in XO form producing both superoxide and NO. It worth mention that simultaneous production of NO and superoxide could produce large amounts of peroxynitrite, which is seen in pathophysiological conditions, and results in severe lipid peroxidation accompanied by the formation of highly toxic lipid and hydroxyl radicals.
Presently, several pathways of nitrite reduction in mammals are known, described in detail in the literature [14]. Of enzymatic pathways capable to reduce nitrite mitochondrial amidoxime reducing component (mARC1 and mARC2), sulfite oxidase, and aldehyde oxidase are most known presently and described in several reviews ([14] and references therein). Common to all these enzymes is Mo- pterin cofactor, where nitrite reduction to NO occurs. Detailed reviews on the structure and functions of these enzymes can be found in the literature [18, 29, 43]. It is Interesting, that besides XOR and cytosolic enzyme aldehyde oxidase (AO), most of the known nitrite reductase activity is localized to mitochondria, which electron transport chain too is capable of nitrite reduction [19, 20]. So, presently, these enzymes capable of functioning as nitrite reductases are considered as putative source of NO, especially under hypoxic and ischemic conditions. However, there were no reports on their nitrate reducing ability yet.
Conclusions and Future Perspectives
So, nitrate-nitrite-NO pathway is supposed to be beneficial in the treatment of several pathophysiological conditions, especially in cardiovascular system (hypertension, pulmonary hypertension, peripheral artery disease and myocardial infarction) [3, 6, 11]. Several enzymes are involved in this pathway, of which XOR enzyme is most studied and most important one according to present views. Besides, presently XOR remains a sole enzyme capable of reducing both inorganic and organic nitrates. However, nitrate reductase activity of XOR should not be overestimated because of its possible side effects such as participation in peroxynitrite production. The time and much effort of researchers are required to decide where there a light, and where the dark side of XOR is. The present data showing nitrate/nitrite reductase activity of XOR indicate that it is a relevant drug target to control a balance between ROS and RNS production in pathophysiological states and diseases.
References
-
Lundberg JO, Weitzberg E (2022) Nitric oxide signaling in health and disease. Cell 185(16): 2853-2878.
-
Tejero J, Shiva S, Gladwin MT (2019) Sources of vascular nitric oxide and reactive oxygen species and their regulation. Physiol Rev 99(1): 311-379.
-
Majuolo J, Oppedisano F, Carresi C (2022) The generation of nitric oxide from aldehyde dehydrogenase -2: the role of dietary nitrates and their implication in cardiovascular disease management. Int J Mol Sci 23: 15454.
-
Cantu-Medellin N, Kelley EE (2013) Xanthine oxidoreductase-catalyzed reactive species generation: a process in critical need of reevaluation. Redox Biology 1(1): 353-358.
-
de Zevallos JO, Woessner MN, Kelley EE (2022) Skeletal muscle as a reservoir for nitrate and nitrite: The role of xanthine oxidase reductase (XOR). Nitric Oxide 129: 102-109.
-
Carlström M, Lundberg JO, Weitzberg E (2018) Mechanisms underlying blood pressure reduction by dietary inorganic nitrate. Acta Physiologica 224(1): e13080
-
Infante T, Costa D, Napoli C (2021) Novel Insights Regarding Nitric Oxide and Cardiovascular Diseases. Angiology 72(5): 411-425.
-
Piknova B, Park JW, Thomas SM, Tunau-Spencer KJ, Schechter AN (2023) Nitrate and nitrite metabolism in aging rats: a comparative study. Nutrients 15(11): 2490.
-
Jones DA, Pellaton C, Velmurugan S, Rathod KS, Andiapen M, et al. (2015) Randomized phase 2 trial of intracoronary nitrite during acute myocardial infarction. Circ Res 116(3): 437-447.
-
Kapil V, Khambata RS, Robertson A, Caulfield MJ, Ahluwalia A (2015) Dietary nitrate provides sustained blood pressure lowering in hypertensive patients: a randomized, phase 2, double-blind, placebo-controlled study. Hypertension 65(2): 320-327.
-
Omar SA, Webb AJ, Lundberg JO, Weitzberg E (2016) Therapeutic effects of inorganic nitrate and nitrite in cardiovascular and metabolic diseases. J Intern Med 279(4): 315-336.
-
Moreno-Vivian C, Cabello P, Martinez-Luque M, Blasco R, Castillo F (1999) Prokaryotic nitrate reduction: molecular properties and functional distinction among bacterial nitrate reductases. J Bacteriol 181(21): 6573- 6584.
-
Sparacino-Watkins C, Stolz JF, Basu P (2014) Nitrate and periplasmic nitrate reductases. Chem. Soc. Rev 43(2): 676-706.
-
Bender D, Schwarz G (2018) Nitrite-dependent nitric oxide synthesis by molybdenum enzymes. FEBS Lett 592(12): 2126-2139.
-
Ahmed KA, Nichols AL, Honavar J, Dransfield MT, Matalon S, et al. (2017) Measuring nitrate reductase activity from human and rodent tongues. Nitric Oxide 66: 62-70.
-
Huang L, Borniquel S, Lundberg JO (2010) Enhanced xanthine oxidoreductase expression and tissue nitrate reduction in germ free mice. Nitric Oxide 22(2): 191- 195.
-
Liu H, Huang Y, Huang M, Wang M, Ming Y, et al. (2023) From nitrate to NO: potential effects of nitrate-reducing bacteria on systemic health and disease. Eur J Med Res 28(1): 425.
-
Maja LB, Moura JJG (2018) Putting xanthine oxidoreductase and aldehyde oxidase on the NO metabolism map: nitrite reduction by molybdoenzymes. Redox Biology 19: 274-289.
-
Bombicino SS, Iglesias DE, Zaobornyj T, Boveris A, Valdez LB (2016) Mitochondrial nitric oxide production supported by reverse electron transfer. Arch. Biochem. Biophys 607: 8-19.
-
Kozlov AV, Staniek K, Nohl H (1999) Nitrite reductase activity is a novel function of mammalian mitochondria. FEBS Lett 454(1-2): 127-130.
-
Berry CE, Hare JM (2004) Xanthine oxidoreductase and cardiovascular disease: molecular mechanisms and pathophysiological implications. J Physiol 555(PT 3): 589-606.
-
Seychell BCh, Vella M, Hunter GJ, Hunter T (2023) The good and the bad: the bifunctional enzyme xanthine oxidoreductase in the production of reactive oxygen species. In: Reactive oxygen species – advances and developments. Intech Open.
-
Yu W, Cheng J-D (2020) Uric acid and cardiovascular disease: an update from molecular mechanism to clinical perspective. Front Pharmacol 11: 582680.
-
Nishino T, Okamoto K, Eger BT, Pai EF, Nishino T (2008) Mammalian xanthine oxidoreductase– mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS Journ 275(13): 3278-3289.
-
Hille R (2023) Xanthine oxidase – a personal history. Molecules 28(4): 1921.
-
Newaz MA, Yousefipour Z, Oyekan A (2006) Oxidative stress-associated vascular aging is xanthine oxidase- dependent but not NAD(P)H oxidase-dependent. J Cardiovasc Pharmacol 48(3): 88-94.
-
Enroth C, Eger BT, Okamoto K, Nishino T, Nishino T, et al. (2000) Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: structure-based mechanism of conversion. Proc Natl Acad Sci USA 97(20): 10723-10728.
-
Kelley EE, Khoo NKH, Hundley NJ, Malik UZ, Freeman BA, et al. (2010) Hydrogen peroxide is a major oxidant product of xanthine oxidase. Free Radic Biol Med 48(4): 493-498.
-
Terao M, Garattini E, Romão MJ, Leimskühler S (2020) Evolution, expression, and substrate specificities of aldehyde oxidase enzymes in eukaryotes. J Biol Chem 295(16): 5377-5389.
-
Porras AG, Palmer G (1982) The room temperature potentiometry of xanthine oxidase. J Biol Chem 257(19): 11617-11626.
-
Saito T, Nishino T (1989) Differences in redox and kinetic properties between NAD-dependent and O2-dependent types of rat liver xanthine dehydrogenase. J Biol Chem 264(17): 10015-10022.
-
Harris ChM, Sanders SA, Massey VJ (1999) Role of the Flavin Midpoint Potential and NAD Binding in Determining NAD Versus Oxygen Reactivity of Xanthine Oxidoreductase. Biol. Chem 274(8): 4561-4569.
-
Hille R, Massey V (1981) Studies on the oxidative half- reaction of xanthine oxidase. J Biol Chem 256(17): 9090- 9095.
-
Alikulov ZA, L’vov NP, Kretovich VL (1980) Nitrate and nitrite reductase activity of milk xanthine oxidase. Biokhimia 45(9): 1714-1718.
-
Millar TM, Stevens CR, Benjamin N, Eisenthal R, Harrison R, et al. (1998) Xanthine xidoreductase catalyses the reduction of nitrates and nitrite to nitric oxide under hypoxic conditions. FEBS Lett 427(2): 225-228.
-
Li H, Samouilov A, Liu X, Zweier JL (2003) Characterization of the magnitude and kinetics of xanthine oxidase- catalyzed nitrate reduction: evaluation of its role in nitrite and nitric oxide generation in anoxic tissues. J Biol Chem 276(27): 24482-24489.
-
Li H, Cui H, Liu X, Zweier JL (2005) Xanthine oxidase catalyzes anaerobic transformation of organic nitrates to nitric oxide and nitrosothiols. J Biol Chem 280(17): 16594-16600.
-
Damacena-Angelis C, Oliveira-Paula GH, Pinheiro LC, Crevelin EJ, Portella RL, et al. (2017) Nitrate decreases xanthine oxidoreductase-mediated nitrite reductase activity and attenuates vascular and blood pressure responses to nitrite. Redox Biol 12: 291-299.
-
Jansson EÅ, Huang L, Malkey R, Govoni M, Nihlén C, et al. (2008) A mammalian functional nitrate reductase that regulates nitrite and nitric oxide homeostasis. Nat Chem Biol 4(7): 411-417.
-
Peleli M, Zollbrecht C, Montenegro MF, Hezel M, Zhong J, et al. (2016) Enhanced XOR activity in eNOS-deficient mice: Effects on the nitrate-nitrite-NO pathway and ROS homeostasis. Free Rad. Biol. Med 99: 472-484.
-
Akopova O, Korkach Y, Sagach V (2024) The effects of ecdysterone and enalapril on nitric oxide synthesis and the markers of oxidative stress in streptozotocin- induced diabetes in rats: a comparative study. Naunyn Schmiedeberg’s Archives of Pharmacology.
-
Lewis SE, Rosencrance CB, De Vallance E, Giromini A, Williams XM, et al. (2021) Human and rodent red blood cells do not demonstrate xanthine oxidase activity or XO-catalyzed nitrite reduction to NO. Free Radic Biol Med 174: 84-88.
-
Clement B, Struwe MA (2023) The History of mARC. Molecules 28(12): 4713.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis