Fish Carboxyl Esterase Activity as a Possible Hypoxia Biomarker
Physiology and biochemistry adaptations studied in fish are important to comprehend how they endure the challenges of living in aquatic environments. Oxygen variations are a frequent natural and cyclic phenomenon that occurs in the Pantanal and Amazon watersheds and it is the limiting factor to survival of the aerobic species. The growing of pollution in these water basins threaten many species as anthropic hypoxia intensifies the production of reactive oxygen species (ROS) and lipid peroxidation inside the cells, leading organisms to a condition known as oxidative stress. Our research group, studying the effects of the oxygen variations on the carboxyl esterase (CarbE) enzyme of pacu, a typical fish from Pantanal, observed that CarbE activity diminished by half in pacus exposed to 42 hours of hypoxia. Based on this finding, we decided to investigate this phenomenon in other fish species, including hypoxia-tolerant fishes. CarbE activities were determined using p-nitrophenyl acetate (p-NPA) as a substrate. All five species of fish studied had both serum and liver CarbE activity significantly decreased. Pacus had its serum and liver activities decreased by approximately 50% under severe hypoxia.
Introduction
Fish and Oxidative Stress
Pacus (Piaractus mesopotamicus), Orcars (Astronotus ocellatus) and Piavussu, (Leporinus macrocephalus) are Neotropical fishes that inhabit Amazonia, Pantanal, Paraná and Paraguay wetland basins. Fish from these regions evolved to resist against hyperoxia and hypoxia conditions since these watersheds are characterized by flood pulses which causes seasonal and daily oxygen variations inside the water [1, 2, 3]. These variations happen in detriment of intense sunlight combined with rainfall, providing high photosynthesis rates during sunlight periods, in contrast to the high rates of oxygen consumption during the night. Oxygen variation can also be initiated by xenobiotic discharges into the aquatic environment which may lead to eutrophication followed by a trophic chain unbalance and depletion in oxygen supply in the water. The fact is that, abrupt changes in the amounts of oxygen inside the water can cause oxidative stress in aquatic organisms. Oxidative stress is characterized by an unbalance between oxidants and antioxidants molecules, which results in an increased production of reactive oxygen species (ROS) and/or reduction in the amounts of the antioxidants. Thus, ROS can be toxic to cells as higher intracellular levels favours alterations of proteins and nucleic acid, resulting in impairment of many cell functions. Biomembranes are also susceptible to ROS attack, resulting in lipid peroxidation followed by fragmentation of polyunsaturated fatty acids and aldehydes production [4, 5]. Oxidative stress encompasses a wide variety of physiological and pathophysiological, endogenous and exogenous processes that directly or indirectly affects cellular homeostasis [6]. In order to survive, fishes developed some strategies such as swim up to superficial waters to breathe [7], besides physiological responses also occurs, such as hyperventilation followed by the increase of the volume and the number of erythrocytes [8, 9, 10]. One of the most important biochemical mechanisms that make feasible the survival of certain species of fish during long periods under low oxygen availability is the depression of the metabolic rate [11, 12, 13]. Another biochemical response is extremely important: the enzyme defence system, an efficient strategy to prevent cell damage against the effects caused by xenobiotic and oxidative stress.
Enzymes
Carboxylesterases (CarbE) (EC 3.1.1.1) are enzymes classified as B-esterases that hydrolyse compounds containing ester, tioester and amide groups. Physiologically, CarbE isoenzymes exhibit lipase activities and are involved in lipid metabolism [14, 15] as hydrolyses cholesterol esters and fatty acids [16, 17, 18]. It has an important role in the xenobiotic metabolism, pro-drugs activation, antitumoral drugs and narcotics, such as cocaine and heroin [19, 20], including the control of the cholinergic impulse. It can also acts as a scavenger of organophosphates and carbamates agrochemicals, functioning as a barrier preventing cellular toxicity [18, 19, 21], and ROS production.
CarbE are found in several tissues including liver, blood, kidney, lower intestine, lungs, adipose tissue and brain of many organisms, but not in human blood [22]. In mammals, the highest levels were found in the liver [19]. CarbE are anchored to the endoplasmic reticulum of hepatocytes trough a Lis-Asp-Glu-Leu (KDEL) amino acid sequence located at the C-terminal enzyme. This sequence can be cleaved to liberate the enzyme in the cytosol and then secreted to the blood [18, 19]. Such importance prompted the esterases to be studied in several areas such as toxicology, biochemistry, pharmacology and medical clinics [19, 23]. In this study we propose to investigate if CarbE enzyme activity from serum and liver of fishes exposed to hypoxia will diminish and suggest this enzyme activity as a possible hypoxia biomarker. We believe that these findings will be of great importance to aquaculture since suitable oxygen levels inside the tanks are essential to fish development, growth and reproduction. CarbE activity as hypoxia biomarker can still contribute to clinical medicine since many organism disturbances and diseases such as cancer, inflammation and embrionary development [24, 25] are related with reduced oxygen levels in the cells.
Material and Methods
Animals and housing
Pacu specimens (Piaractus mesopotamicus) of both sexes, weighing 250 g were supplied by Morro Grande Fish Farm (Cachoeira de Macacu, Rio de Janeiro, Brazil). Carps (Cyprinus carpio) weighing 122 g, Oscars (Astronotus ocellatus) weighing 298 g and Goldfish (Carassius auratus) weighing 25 g were bought in a pet shop in the city of Nova Iguaçu (Rio de Janeiro, Brazil). Piavussu (Leporinus macrocephalus) weighing 888 g were supplied by São João Fish Farm located at Itaocara city, Rio de Janeiro, Brazil. Procedures of experiments with the fishes were carried out in conformity with the ethical principles in animal experimentation elaborated by the Brazilian College for Animal Experimentation (COBEA), which is in accordance with the uniform requirements for submission of manuscripts to biomedical journals. The fishes were acclimated in 1000 L water tanks and fed with standard fish food. Water temperature was kept at 23 ºC and dissolved oxygen concentration (OC) at 6.0 mg L-1. Water pH was maintained at 6.4 ± 0.2. Fishes were kept in these conditions for one week until submitted to hypoxia.
Hypoxia experiment
In order to investigate the effect of hypoxia on serum and liver CarbE activities, the fishes were distributed in two groups: normoxia and hypoxia. The oxygen concentration inside the normoxia tank was maintained at 6.0 mg L-1 along the whole experiment. Nitrogen gas was bubbled inside the hypoxia tank for five hours until the oxygen concentration reached 0.5 mg L-1 and the fishes were submitted to this oxygen concentration until the end of the experiment. Aerial surface respiration was avoided by covering all tanks flush at the water level with tight mesh nylon. The time of hypoxia exposure varied according to the species, as fish species do not exhibit the same resistance to hypoxia. All fish from the different groups were euthanatized by severing their spinal cords and had their liver excised.
Serum Preparation
Blood was collected from the caudal vein of the fishes in the end of each experiment. Afterwards, blood was kept inside the ice until coagulate. The coagulated blood was centrifuged at 3.500 rpm during 10 min and the supernatant (the serum) was used in CarbE assays.
Hepatic Preparation
Livers were weighted and homogenized using a Potter- Elvehjem type apparatus in four volumes of 0.1 M potassium phosphate buffer, pH 7.0, containing 0.25 M of sucrose. The homogenate were stored at - 20 ºC until assays. Protein content was determined according to Peterson GLA, et al. [26] using bovine serum albumin as standard.
Enzymatic Assays
The carboxylesterase (CarbE) activities were determined in liver homogenate and serum from each fish using a Shimadzu UV-160 A spectrophotometer according to a kinetic reaction [27]. The p-NPA hydrolyses was measured at 400 nm by absorbance increase of the product reaction (p-nitrophenol) along 1 min. CarbE activities were calculated using the extinction coefficient of 13,000 M- 1.cm-1.
Serum CarbE Assays
The reaction was started by adding the substrate p-nitrophenyl acetate (p- NPA) in a final concentration of 3 mM in a cuvette containing 1µL of serum and a volume of a potassium phosphate buffer 0.1 M, pH 7.7 at a 25 ºC to reach 0.5 mL. Serum CarbE activities were expressed in U.mg-1 protein and U.mL-1.
Liver CarbE Assays
The reaction was started by adding p-NPA in a final concentration of 3 mM in a cuvette that contained 5 µL of liver homogenate and a volume of a potassium phosphate buffer 0.1 M, pH 7.7 at a 25 ºC to reach 0.5 mL. The CarbE liver activities were expressed in U mg-1 protein or U g-1 wet tissue.
Statistical Analysis
Data are presented as mean ± SD of CarbE assays carried out using one sample from one serum or homogenate prepared from each tissue. Comparisons between data were made by nonparametric T-test analysis (p < 0.05) using the software GraphPad Prism, San Diego, California, USA.
Results
Blood partial oxygen pressure analysis
The oxygen levels were determined in the blood of two fish species (Astronotus ocellatus and Piaractus mesopotamicus) using AVL / Roche OMNI 7 instruction. These analyses were made to assure that fishes were indeed submitted to hypoxia during the experiments (Tables 1 & 2).
| Blood partial oxygen pressure | |
|---|---|
| Normoxia | 65.0 ± 9.0 mm Hg |
| Hypoxia | 18.0* ± 2.0 mm Hg |
Table 1: Blood partial oxygen pressure in oscar, Astronotus ocellatus. *Denotes statistical significance (p < 0.05) from control
| Group | Blood partial oxygen pressure |
| Normoxia | 30.25 ± 4.25 mm Hg |
| Hypoxia | 1.16* ± 0.6 mm Hg |
Table 2: Blood partial oxygen pressure in pacu, Piaractus mesopotamicus.*Denotes statistical Significance (p < 0.05) from control
| Serum | Liver | |||||||
|---|---|---|---|---|---|---|---|---|
| FISH | NORMOXIA U.mL-1 | HIPOXIA U.mL-1 | NORMOXIA mU.mg-1 ptn | HIPOXIA mU.mg-1 ptn | NORMOXIA U.g-1 w t | HIPOXIA U.g-1 w t | NORMOXIA mU.mg-1 ptn | HIPOXIA mU.mg-1 ptn |
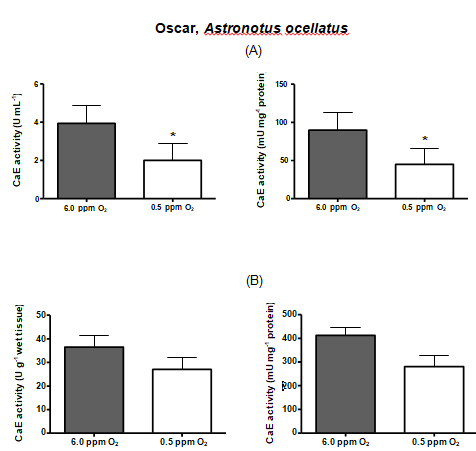
| OSCAR | 3.93 | 2.0* | 89.68 | 45.03* | 36.53 | 27.02 | 412.25 | 280.93 |
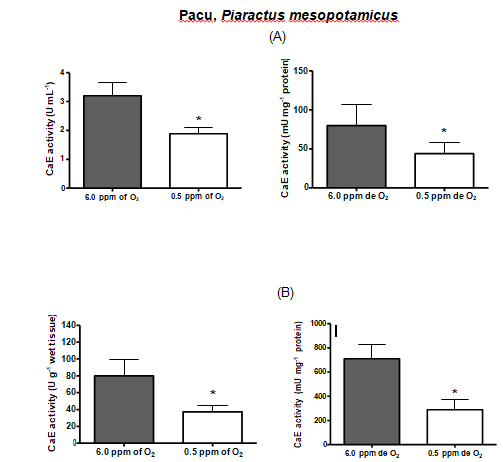
| PACU | 3.2 | 1.88* | 79.9 | 43.94* | 79.84 | 37.27* | 709.58 | 288.80* |
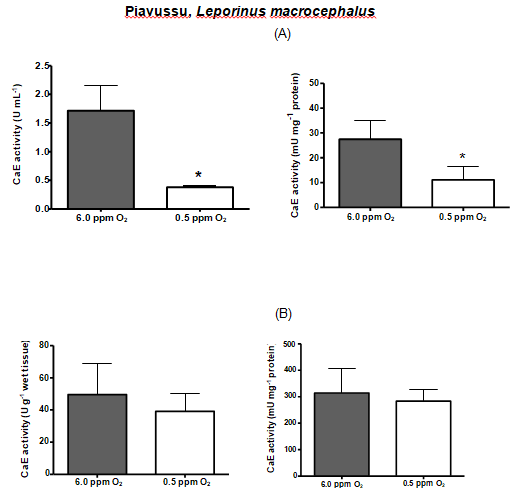
| PIAVUSSÚ | 1.71 | 0.38* | 27.48 | 11.11* | 49.63 | 39.22 | 314.36 | 283.6 |
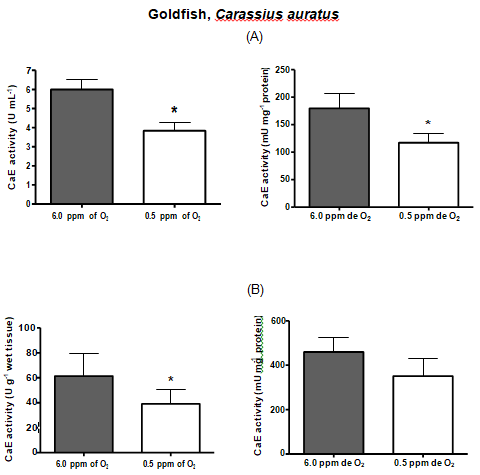
| GOLDFISH | 6 | 3.84* | 179.73 | 117.3* | 61.46 | 39.1* | 459.63 | 351.1 |
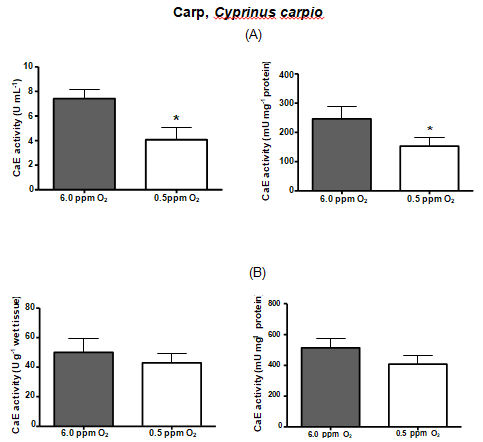
| CARP | 7.41 | 4.06* | 246.58 | 153.1* | 50.1 | 43 | 513.58 | 407.1 |
Table 3: Comparison between serum and liver carboxylesterase activities among the fish species.* Denotes statistical significance
Discussion and Conclusion
There are some organisms that are able to withstand changes in the amount of dissolved oxygen in their blood for months. This is possible thanks to physiological and biochemical adaptations that occurs in their organs to overcome hypoxia [28, 29, 30, 31]. Hypoxia induces a family of transcriptional factors, called hypoxia induced factors (HIF) that is linked to the hypoxia response elements (HRE) to do proteins transcription and genes regulation. These genes are associated to erythropoietin, endothelial vascular growth factor, glucose transport, glycolytic enzymes and also enzymes related to iron metabolism and cell’s survival [32, 33]. In Goldfish, for example, its gills excrete ethanol produced in its muscles from lactate originated from glucose, which avoids the disadvantageous acidosis that would result if lactate were accumulated in hypoxia [34]. Severe hypoxia is known to produce the release of catecholamines and other stress hormones that are responsible for the hyperglycemic response through stimulation of glucogenolysis and/or gluconeogenesis [35, 36, 37]. Indeed, some fishes survive by mobilizing carbohydrate reserves to generate ATP molecules by anaerobic glycolysis. Carps tissues contain large glycogen stores and also show some important additional metabolic adaptations in their use [13]. A depression in the levels of free fatty acids of carps and rainbow trout plasma were also observed during hypoxia, and the mechanism for that is most probably a reduction of free fatty acids oxidation and mobilization, a consequence of the high plasma glucose concentration along hypoxia [37]. More recently, hypoxic inhibition of lipolysis has been demonstrated in humans [38] and results suggest increased concentration of glucose and fatty acid mobilization is also present in fish [39].
Hypoxia is thought to produce a general shift in oxidative fuel preference, from predominantly lipids and proteins to carbohydrates. Whereas carboxylesterase (CarbE) is the main nervous tissue lipase [16], a diminished flux of fatty acids and its reduced levels, could be linked to a decrease in the CarbE enzymatic activity during hypoxia. Indeed, human hepatocytes (HEPG2) maintained in hypoxia for 24 hours had several proteins with their gene expression depleted, including the carboxyl esterase [40]. Another experiment with human tumor cells, and tumor cells suffer from hypoxia, shows a reduction in the CarbE expression [41]. Moreover, studies realized by Suto D, et al. [42] demonstrated that singlet oxygen, generated during hypoxia, is a strong inhibitor of cysteine and serine proteases and can possibly inhibit CarbE activities as catalytic CarbE sites exhibits serine and histidine residues. In this present study, we attempted to submit the fish species to equal time of hypoxia, however, this was not possible. Each fish species exhibit different tolerance to the low concentrations of oxygen in the water and some species eventually died along the experiment. Therefore, the time of hypoxia exposure for each fish species was settled particularly and was the limit time between the fish death and the experiment accomplish. Pacu was the most resistant fish species, surviving 42 hours of hypoxia and had their both serum and liver CarbE activities diminished almost by half. Oscars also had their activities diminished by half but only endured 24 h of hypoxia. Piavussu and carps also endured 24 h of hypoxia. Piavussu exhibited the lowest CarbE activities among all fish species while carps exhibited the highest activities. Goldfish also exhibit high CarbE activities, however they only stud up for 8 hours of hypoxia. The relevance of this study is that all fish species submitted to hypoxia in this present study had their both serum and liver CarbE activities diminished. We believe that serum CarbE activities decreased significantly because fewer liver CarbE enzymes were secreted to the blood. Perhaps, the CarbE are more useful in the liver rather than in the blood, since lipid metabolism could be modified in response to hypoxia. The decreases in oxidation and mobilization of fatty acids have been previously related to the high glycemic levels during hypoxia [37]. Since CarbE activities are essential for mobilize and oxidizelipids, it is understandable that these activities are also diminished during hypoxia. In addition, such depletion is a convenient adaption to the lipid metabolism because fishes submitted to hypoxia produce ethanol. Ethanol is a substrate of CarbE to form ethyl esters of fatty acids, a chaotropic product of mitochondrial membranes, resulting in the formation of reactive oxygen species. Thus, as CarbE activity decreases, the formation of the ethyl fatty acid esters also decreases and the integrity of membranes are preserved during low oxygen periods. Our goal in this study was achieved. All species had both liver and serum carboxyl esterase (CarbE) activity diminished, but serum activities were mightily decreased. These findings suggest that CarbE activities, especially in serum, can be used as a hypoxia biomarker in fish. What´s more, to draw the blood, it will not be necessary to kill the fish for enzyme analyses, an advantage for ecotoxicology studies and environmental preservation [43]. Another important conclusion is that fish may be more susceptible to xenobiotics when abiding in waters with low oxygen content, since their serum and hepatic CarbE, an important drug metabolism enzyme, decreases during hypoxia.
References
-
Calheiros DF, Seidl AF, Ferreira CJA (2000) Participatory research methods in environmental science: local and scientific knowledge of a limnological phenomenon in the Pantanal wetland of Brazil. Journal of Applied Ecology 37(4): 684-696.
-
Bastos VLFC, Salles JB, Valente RH, León IR, Perales J, et al. (2007) Cytosolic glutathione peroxidase from liver of pacu (Piaractus mesopotamicus), a hypoxia tolerant fish of the Pantanal. Biochemie 89(11): 1332-1342.
-
Leite CAC, Florindo LH, Kalinin AL, Milsom WK, Rantin FT (2007) Gill chemoreceptors and cardio-respiratory reflexes in the neotropical teleost pacu, Piaractus mesopotamicus. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 193(9): 1001-1011.
-
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and biochemistry of 4-Hydroxynonenal, malonaldehyde, and related aldehydes. Free Radical Biology & Medicine 11(1): 81-128.
-
Yang Y, Sharma R, Sharma A, Awasthi S, Awasthi YC (2003) Lipid peroxidation and cell cycle signaling: 4 -hydroxynonenal, a key molecule in stress mediated signaling. Acta Biochimica Polonica 50(2): 319-336.
-
Ma Q (2009) Transcriptional responses to oxidative stress: Pathological and toxicological implications. Pharmacology & Therapeutics 125(3): 376-393.
-
Wu RSS (2002) Hypoxia: from molecular responses to ecosystem responses. Marine Pollution Bulletin 45(1- 12): 35 45.
-
Nikinmaa M (2001) Haemoglobin function in vertebrates: evolutionary changes in cellular regulation in hypoxia. Respiration Physiology 128(3): 317-329.
-
Nikinmaa M, Rees BB (1987) Oxygen-dependent gene expression in fishes. The American Journal of Physiology – Regulatory, Integrative and Comparative Physiology 288(5): 1079-1090.
-
Storey KB (1987) Tissue-specific controls on carbohydrate catabolism during anoxia in goldfish. Physiological Zoology 60(5): 601-607.
-
Van Wavesveld J, Addink ADF, van den Thillart G (1989) The anaerobic energy metabolism of goldfish determined by simultaneous direct and indirect calorimetry during anoxia and hypoxia. Journal of Comparative Physiology 159: 263-268.
-
Storey KB, Storey JM (1990) Metabolic rate depression and biochemical adaptation in anaerobiosis, hibernation and estivation. Quarterly Reviews in Biology 65(2): 145- 174.
-
Hochachka PW, Somero GN (1984) Biochemical adaptation. Princeton University Press, USA, pp: 355.
-
Satoh T (2005) Toxicological implications of esterases – From molecular structures to functions. Toxicology and Applied Pharmacology 207(2): 11-18.
-
Ross MK, Streit TM, Herring KL, Xie S (2010) Carboxylesterases: dual roles in lipid and pesticide metabolism. Journal of Pesticide Science 35(3): 257-264.
-
Soni KG, Lehner R, Metalnikov P, O’donnel P, Semache M, et al. (2004) Carboxylesterase 3 (EC 3.1.1.1) is a major adipocyte lipase. The Journal of Biological Chemistry 279 (39): 40683-40689.
-
Okazaki H, Igarashi M, Nishi M, Tajima M, Sekiya M, et al. (2006) Identification of a novel member of the carboxilesterase family that hydrolyzes triacylglycerol: A potential role in adipocyte lipolysis. Diabetes 55(7): 2091-2097.
-
Satoh T, Hosokawa M (2006) Structure, function and regulation of carboxylesterases. Chemico-Biological Interactions 162(3): 195-211.
-
Redinbo MR, Potter PM (2005) Mammalian carboxylesterares: from drug targets to protein therapeutics. Drug discovery Today 10(5): 313-325.
-
Holmes RS, Cox LA, Vandeberg JL (2008) Mammalian carboxylesterase 5: Comparative biochemistry and genomics. Comparative Biochemistry and Physiology 3(3): 195-204.
-
Wheelock CE, Nakagawa Y (2010) Carboxylesterases – from function to the field: an overview of carboxylesterase biochemistry, structure–activity relationship, and use in environmental field monitoring. Journal of Pesticide Science 35(3): 215-217.
-
Li B, Sedlacek M, Manoharan I, Boopathy R, Duysen EG, et al. (2005) Butyrylcholinesterase, paraoxonase, and albumin esterase, but no carboxylesterase, are present in human plasma. Biochemical Pharmacology 70(11): 1673-1684.
-
Ghazala SM, L Ahmad S, Sultana K, Alghanim F, Al-Misned Z, et al. (2014) Fish cholinesterases as biomarkers of sublethal effects of organophosphorus and carbamates in tissues of Labeo rohita. Journal of Biochemical and Molecular Toxicology 28(3): 137-142.
-
hockel M, Vaupel P (2001) Biological consequences of tumor hypoxia. Seminars in Oncology 28(2): 36-41.
-
Rodway GW, Hoffman LA, Sanders MH (2003) High- altitude-related disorders-part I: pathophysiology, differential diagnosis, and treatment. Heart & Lung: The Journal of Acute and Critical Care 32(6): 353-359.
-
Peterson GLA (1977) Simplification of the protein assay method of Lowry et al., which is more generally applicable. Analytical Biochemistry 83(2): 346-356.
-
Morgan WE, Yan B, Greenway D, Petersen DR, Parkinson A (1994) Purification and characterization of two rat liver microsomal carboxylesterases (hydrolase A and B). Archives of Biochemistry and Biophysics 315(2): 495- 512.
-
Costanzo JP (1989) A physiological basis of prolonged submergence in hibernating garter snakes Thamnophis sirtalis: evidence for an energy-sparing adaptation. Physiological Zoology 62(2): 580-592.
-
Willmore WG, Storey KB (1997) Glutathione systems and anoxia tolerance in turtles. American journal of physiology Regulatory, integrative and comparative physiology 273(2): R219-R225.
-
Hermes-Lima M, Storey JM, Storey KB (2001) Antioxidant defenses and animal adaptation to oxygen availability during environmental stress. In: Cell and Molecular Responses to Stress, Storey KB, Storey JM (Ed.), Protein adaptations and signal transduction. Elsevier Press, pp: 263-287.
-
Lushchak VL, Bagnyukova TV (2006) Effects of different environmental oxygen levels on free radical processes in fish. Comparative Biochemistry and Physiology Part B: Biochemestry and Molecular Biology 144 (3): 283-289.
-
Huang Le, Bunn HF (2003) Hypoxia-inducible factor and its biomedical relevance. The Journal of Biological Chemistry 28 (22): 19575-19578.
-
Guzi RD, Schumaker PT (2006) Oxygen sensing by mitochondria at complex III: the paradox of increased reactive oxygen species during hypoxia. Experimental Physiology 91 (5): 807-819.
-
Shoubridge EA, Hochachka PW (1980) Ethanol: novel end product of vertebrate anaerobic metabolism. Science 209(4453): 308-309.
-
White A, Fletcher TC (1989) The effect of physical disturbance, hypoxia and stress hormones on serum components of the plaice, Pleuronectes platessa L. Comparative Biochemistry and Physiology 93(2): 455-
-
Wright PA, Perry SF, Moon TW (1989) Regulation of hepatic gluconeogenesis and glycogenolysis by catecholamines in rainbow trout during environmental hypoxia. The Journal of Experimental Biology 147: 169- 188.
-
Van Raaij MTM (1996) Substrate mobilization and hormonal changes in rainbow trout (Oncorhynchus mykiss, L.) and common carp (Cyprinus carpio, L.) during deep hypoxia and subsequent recovery. J Comp Physiol 166: 443-452.
-
Wolfe RR, Peters EJ (1987) Lipolytic response to glucose infusion in human subjects. American Journal of Physiology 252(2 Pt 1): E218-E223.
-
haman F, Zwingelstein G, Weber J (1997) Effects of hypoxia and low temperature on substrate fluxes in fish: plasma metabolite concentrations are misleading. The American Journal of Physiological-Regulatory, Integrative and Comparative Physiology 273(6): 2046- 2054.
-
sonna LA, Cullivan ML, Sheldon HK, Pratt RE, Lilly CM (2003) The effect of hypoxia on gene expression by human hepatocytes (HEPG2). Physiological Genomics 12 (3): 195-207.
-
Guichard S, Terret C, Hennebelle I, Lochon I, Chevreau P, et al. (1999) CPT-11 converting carboxilesterase and topoisomerase I activities in tumour and normal colon and liver tissues. British Journal of Cancer 80(3/4): 364- 370.
-
Suto D, Iuchi Y, Ikeda Y, Sato K, Ohba Y, et al. (2007) Inactivation of cysteine and serine proteases by singlet oxygen. Archives of Biochemistry and Biophysics 461(2): 151-158.
-
Hochchaka PW, Buck LT, Doll CJ (1996) Unifying theory of hypoxia tolerance: Molecular/metabolic defense and rescue mechanisms for surviving oxygen lack. Proceedings of the National Academy of Sciences of the United States of America 93 (18): 9493-9498.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh