Analysis via Atomic Force Microscopy of Streptococcus bovis Under Tylosin Phosphate and Virginiamycin Treatment
Antibiotics are widely used in modern animal husbandry towards food production, to increase growth, and reduce pathogen colonization and shedding. The body of literature published concerning antibiotic use demonstrates that the continuous use of antibiotics can undesirably improve and accumulate bacterial antibiotic resistance, therefore imperiling much of modern animal husbandry. Thus, understanding the emergence of antimicrobial resistance and how pathogens respond to antibiotics is critical to reduce antibiotic use and antibiotic resistance. Towards this, mechanical characterizations must be supplied where possible to accompany molecular data to understand the surface, physical interactions of both pathogens, and the surfaces that they interface with. As a first step towards understanding how rumen microbes react to antibiotics, we investigated at a molecular level how Streptococcus bovis, a major contributor to lactic acidosis in the rumen, responds to tylosin phosphate and virginiamycin treatment using a time series exposure experiment while characterizing and imaging microbial responses using atomic force microscopy on various media. It was found that a supply of current typically improves bacterial surface modulus of elasticity, which may pave the way for an effective means of interfacing with bacteria as opposed to the use of antibiotics.
Introduction
Streptococcus bovis (S. bovis) is a commensal bacterium commonly associated with production livestock, such as bovines and equines, as well as humans [1, 2]. As such, a recent study has reported significant correlations between the rates of S. bovis bacteremia and cattle densities with the possibility of zoonotic transmission to humans [2]. Within this study, it was reported S. bovis biotype I is commonly associated with endocarditis and colon neoplasms [2]. Another biotype (II) of S. bovis, has been seen to be associated with infections of primary bacteremia and biliary tract, urinary tract, meningitis, neonatal sepsis, liver disease, and non- colorectal cancer [2]. Concerning rural human populations and outward to urban populations, pets notwithstanding, close contact with and or consumption with infected animal products, could result in an increased spread of disease. As such, additional stress on health systems worldwide, wherein cattle are largely and densely used in agriculture, with economic effects not withstanding [2].
Already, it is noted that under immense immunological stress, early carcinogenesis has been linked with the presence of S. bovis in patients [3]. Thus, controlling S. bovis numbers remain paramount. Still, means of keeping numbers below safe numbers purely with antibiotics is increasingly less tenable as market demands shift animal husbandry toward increasingly dense housing for cattle. As such, this results in an increased transmission with antibiotic use leading to more resistant strains of S. bovis. Simply increasing levels of antibiotics is both irresponsible and expensive, for environmental, health based, and economic considerations, for both human and livestock in mind. Thus, means to control or reduce levels of S. bovis populations in livestock without such is important.
In this study, we examined now minimally used treatments used in the beef cattle industry, the administration of tylosin phosphate and virginiamycin, and through pilot experiments aimed at characterizing the response of S. bovis under antibiotic stress as well as without. Tylosin phosphate is a growth promoter and has long been used towards reducing the formation of liver abscess, increasing feed efficiency, and improving weight gain among livestock. However, tylosin has been restricted and now requires a veterinarian’s approval [4, 5]. Tylosin phosphate, as with Virginiamycin, and numerous others, have been banned by the EU [5]. Virginiamycin is a complex of two antibiotics, virginiamycin M and virginiamycin S, and they work together in controlling enteritis infection, which would otherwise become necrotic in modern husbandry [6, 7]. In some cases, virginiamycin has been noted to produce behavioral changes, thus giving increased reasons for moderation in administration to livestock [8]. Both antibiotics are added to feeds or are given orally, and their effects have prompted concerns with effects on humans who consume animals that are fed these antibiotic compounds [5, 8]. Although a veterinarian’s approval is needed to utilize these antibiotics in the US, these antibiotics are still used in some capacity by large beef producers to treat sick animals and by producers raising high-risk animals. As such, investigating the effect of such antibiotics and its effects on members of the rumen commensal microbiome and emergence of antibiotic resistance is remains important to monitor for livestock in recovery. In this study we investigated the effect of Tylosin phosphate and Virginiamycin on S. bovis to evaluate how rumen microbes might be affected and response to common antibiotic treatments. This may assist in the long term goal of reducing or eliminating antibiotic dependence in animal husbandry by providing mechanical data that can be correlated with tylosin phosphate and virginiamycin use. It must be noted that data will be of greater use with a wider array of more commonly used antibiotics and treatments, but for now, the use of the applied antibiotics can serve as a model for following experiments in this vein.
As demonstrated in prior work, atomic force microscopy (AFM) has shown remarkable value in biological materials, especially mammalian cells and tissues, concerning surface forces and morphologies using AFM cantilever tips [9, 10, 11, 12, 13]. This also extends to subcellular features such as collagen nanofibers in multiple environmental conditions, allowing researchers to gauge the variability of mechanical features of subcellular biological compounds under differing environmental conditions. In such work, minimal deformation of samples through indentation applied over such samples surfaces allows extraction of material properties that can reach the nanometer scale for soft surfaces and the pico-scale scale for hard surfaces [14]. Additionally, this is important for the examination of mechanical changes of both cancerous cells and tissues, in which AFM is increasingly relevant, given that high resolutions that it allows can yield powerful insights into mechanical cell changes as indicated by recent research [15, 16, 17, 18, 19]. In such, this work adds specifically to the examination of bacteria under AFM which delivers a helpful addition to the current and strong body of prior work by others [20, 21, 22, 23, 24, 25]. Fang, et al. reported the use of AFM to quantify bacterial adhesion forces [24]. Doktycz, et al. reported the use of AFM to image bacteria in liquid media that were immobilized on mica surfaces coated with gelatin [20]. Schaer-Zammaretti and Ubbink reported the use of elasticity and adhesion maps in their AFM-assisted characterization of lactic acid bacteria, along with inferring relationships to biological and structural data [22]. Lastly, Kailas, et al. and Meyer, et al. reported characterizations of live bacteria via AFM to glean various physiological processes [21, 23]. Recent work by Dutta, et al. reported a comparative study of force measurements within solutions between micron and nano sized probes [9]. Together, these works provide a basis for comparative measurements samples to build from, from which a limited, preliminary study is provided for further development.
Limitations in fixation encouraged a limited study with resources available. The goal was to provide a preliminary study to help begin to understand the mechanistic changes that occur in rumen microbes in response to antibiotic treatment (the hypothesis being if a change in Young’s Modulus could be detected). Fully addressed, this can help address knowledge gaps in mechanical properties of bacterial surface changes. To this end, we investigated how S. bovis a common organism found in the rumen may mechanically respond to some antibiotics used in industry to treat sick and high-risk animals.
Materials and Methods
Pure Cultures and Antibiotic Exposure Experiment Preparation
S. bovis ATCC 33317 was grown on BD BBL™ Brain Heart Infusion (BHI) for 24 hours at 37°C. The overnight culture was aliquoted (1000 ml into sterile 2.0 ml Eppendorf tubes) and incubated with either tylosin phosphate (Huvepharma, Lincoln NE, USA) or vir-giniamycin (Cayman Chemical, Ann Arbor MI, USA) at a final concentration of 100mg/ml. Samples were collected immediately, at 15 minutes, 30 minutes, 60 minutes, 4 hours, 6 hours and 24 hours to be immediately fixed in 4 % formaldehyde for examination. Briefly, the cells were centrifuged at 5000 XG for ten minutes immediately after collection and the supernatant was removed and discarded. The resulting pellet was re-suspended in 4% formaldehyde and stored at 4°C until subsequent microfluidic assessment.
Atomic Force Microscopy (AFM) and Data Acquisition across Experiments
Microscopy techniques used were similar to those from multiple previous works [9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19]. An atomic force microscope (AFM) from Oxford Instruments, Santa Barbara, CA, was used to scan the samples and also measured the forces on the samples including cells, PDMS, and related. AFM consists of a cantilever and probe. In this study, MSNL silicon nitride AFM probes with multiple cantilevers from Bruker, Santa Barbara were used for scanning samples of interest for force data collection. AFM can work in contact mode, tapping, and also intermittent mode. Here we used the contact mode method to get a better resolution of the AFM images. Before performing the actual AFM measurements, we measured the resonance frequency of the cantilever. The value of the cantilever’s resonance frequency was found to be between 85 and 155 kHz. The thermal method was employed before each experiment to calculate the spring constant, which was k = 859 pN/nm. AFM probe was calibrated before experimentation with known sample and the sample was PDMS. Forces measured over the PDMS surface and the force curve was fitted with the Sneddon model and the measured Young’s modulus was 1.8MPa. AFM was used to image S. bovis cells incubated with Virginiamycin and Tylosin phosphate at different time duration in both air and PBS. Force vs displacement curves were performed above the samples in different chemical treatments at different time duration. The force curves were fitted with the Sneddon model, which was shown in equation 1. F is the force, E is Young’s modulus and AFM tip Young’s modulus was 290 GPa, α is the half- angle of the intender and the angle was 720, ν is the Poisson ratio, sample Poisson ratio was 0.5 and AFM tip Poisson ratio was 0.25 and δ is the indentation depth that is very from experiment to experiment. Force measurements were performed over the samples and the force vs displacements curves were fitted Sneddon model using Igor Pro 6.23 software package. After analyzing the force vs displacement curve, samples of Young’s modulus were measured with Igor Pro 6.23 software package.
$$ F = \frac {2}{\pi} \frac {E}{\left(1 - v ^ {2}\right)} \tan (\alpha) \delta^ {2} $$ N represents the number of force curves that were analyzed in Table 1. 144 force curves were analyzed for S. bovis cells incubated with Virginiamycin at 0-time duration. The number of force curves was analyzed in each condition as shown in Table 1. When indentation was performed, the range of the loading force was from 0.5 nN to 2 nN. AFM probe can approach the surface when force is measured and also retract at constant velocity and the speed was 300 nm/s. Force maps were performed above the samples that are shown in the AFM images in a rectangular box. By changing trigger values, multiple force versus displacement curves were carried out on the cells at different chemical treatments.
Statistics
Two sample t-tests were performed for S. bovis cells incubated with Virginiamycin and Tylosin phosphate at different time durations. Values were determined statistically significant when p values equaled less than 0.05. P values show less than 0.05 in most of the combinations, except P- Value for 1-V to 24-V in air was 0.0707852 and P-Value for 0-T to 1-T in air was 0.0209970.
Results
Figures 1-4 below include height images bacterial cells as imaged through atomic force microscopy. Scale bars are provided to their right for understanding of depth, per image. Figure 5 provides Modulus of Elasticity data for S. bovis, under differing environmental regimes.




Figure 2: AFM images of S. bovis incubated with tylosin phosphate in air at 0 hour at increasing magnification under phase imaging. (a) The top image includes a zoom out of a bacterial cell, while the bottom includes a closer image. (b) This image includes a phase image, which is a reconstructed image derived from height deflection data. A flattened surface and structure can be observed, which demonstrates accuracy of height image data. Phase image does not show the roughness of the surface; however, they show chemical composition of the sample.



Figure 4: AFM images of S. bovis samples at 1hr and 24 hr time-points (a) AFM image of S. bovis incubated with Virginiamycin (Cayman Chemical) in air at 1 hour. A curvy surface can be observed. (b) AFM image of S. bovis incubated with tylosin phosphate in air at 1 hour. (c) AFM image of S. bovis incubated with Virginiamycin (Cayman Chemical) in air at 24 hours. This is a further zoomed out image of 3a and 3b (d) AFM image of S. bovis incubated with tylosin phosphate in air at 24 hours. Bacteria in this image are more spread out incomparison to those in 4c.
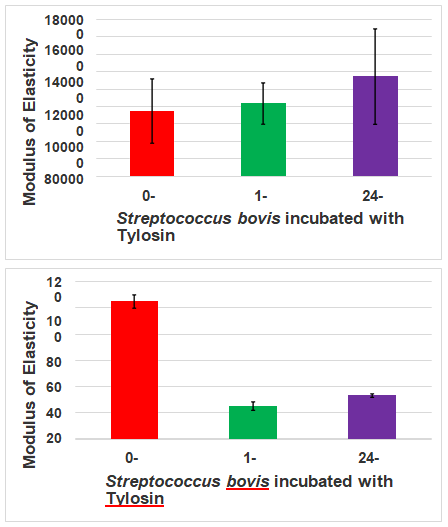
Figure 5: Modulus of Elasticity differences found at different time points and different media. (a) Modulus of Elasticity of S. bovis ATCC 70040 incubated with Virginiamycin in air at 0 hour, 1 hour and 24 hours; (b) Modulus of Elasticity of S. bovis ATCC 70040 incubated with Virginiamycin in PBS at 0 hour, 1 hour and 24 hours; (c) Modulus of Elasticity of S. bovis ATCC 70040 incubated with Tylosin phosphate in air at 0 hour, 1 hour and 24 hours; (d) Modulus of Elasticity of S. bovis ATCC 70040 incubated with tylosin phosphate in PBS at 0 hour, 1 hour and 24 hours.
| Virginiamycin Treatment | Air | Virginiamycin Treatment | PBS | ||||
|---|---|---|---|---|---|---|---|
| Measurement time | E (KPa) | ±SE (±SD) | N | Measurement time | E (KPa) | ±SE (±SD) | Number of Force Curves Analyzed |
| 0 hours | 85,100 | 2100(23100) | 144 | 0 hours | 0.88 | 0.54 (0.17) | 15 |
| 1 hour | 131,200 | 7000(65200) | 137 | 1 hour | 2.92 | 0.12 (0.76) | 24 |
| 24 hours | 119,200 | 7000(41000) | 135 | 24 hours | 2.04 | 0.10 (0.49) | 19 |
Table 1: Modulus of Elasticity, Standard Error, Standard Deviation and N represent number of force curves analyzed of S. bovis AT
| Tylosin phosphate Treatment | Air | Tylosin phosphate Treatment | PBS | ||||
|---|---|---|---|---|---|---|---|
| Measurement time | E (KPa) | ±SE (±SD) | Number of Force Curves Analyzed | Measurement time | E (KPa) | ±SE (±SD) | Number of Force Curves Analyzed |
| 0 hours | 75000 | 2600(36800) | 139 | 0 hours | 105.1 | 11.7 (5.2) | 23 |
| 1 hour | 83500 | 1500(23600) | 144 | 1 hour | 24.8 | 2.9 (3.3) | 35 |
| 24 hours | 114400 | 2700(54900) | 137 | 24 hours | 33.2 | -1.4 | 10 |
Table 2: Modulus of Elasticity, Standard Error, Standard Deviation and N represent number of force curves analyzed of S. bovis AT
In examining S. bovis via AFM, we utilized contact mode to mechanically assess the fixed samples over the course of a single day at 3 separate time points (0 hours, 1 hour, and 24 hours) in both air and Phosphate-Buffered Saline (PBS) conditions. Topographic imaging revealed that the S. bovis were distributed among the sample plate after fixation and retained their cocci shape (Figure 1). Samples exposed to tylosin phosphate were more dispersed and flattened with no neighbors to crowd, as shown from zoomed-in topographic and phase imaging (Figure 2). In contrast, the same imaging revealed dense, bunched up S.bovis that retained a more spherical shape (Figure 3). Time exposure does not seem to be a factor in the shape, given the existence of the spherical form of S.bovis evident with exposure to virginiamycin (Figure 3). They more or less retain their dispersal and shape, as shown on the sample plate, globally, from the zoomed- out images (Figure 4). The formaldehyde fixation process may have played a role in the mechanical measurements, as formaldehyde is known to modify surface proteins. However, the preserved shapes as well as differences in structure suggest some reasonable insights can be gleaned. Testing revealed different mechanical responses, showing changes in each sample’s Young’s Modulus of Elasticity in samples at each time point, regardless of being in air or PBS, suggesting meaningful changes to the cytoskeleton, which confers both structure and mechanical strength to the cell.
In examining the modulus of elasticity of S. bovis under both of these regimes, it was found that in an hour, the modulus of elasticity increased widely. In the air, under a virginiamycin regime from t=0 to t=1, a 54% increase in modulus of elasticity was observed, with a slight decrease between t=1 and t=24 to 40.07%, reflecting a wide window in which these changes within S. bovis could be exploited (Figure 5A). In PBS, under a virginiamycin regime from t=0 to t=1, a 232% increase in modulus of elasticity was observed, with a considerable decrease between t=1 and t=24 of 132%. This reflects a less wide, but still useful window for S. bovis could be exploitation as the difference between t =0 and t = 24 is a 231% increase (Figure 5B). The modulus of elasticity values for S. bovis under tylosin phosphate reflected the differences seen under contact mode observation. In the air, under a tylosin phosphate regime from t=0 to t=1, an 11% increase in modulus of elasticity was observed, with a further increase of 53% between t=1 and t=24, reflecting a dramatically wider window in which these changes within S. bovis could be exploited (Figure 5C). In the air, under a virginiamycin regime from t=0 to t=1, a -76.4% decrease in modulus of elasticity was observed, with a slight increase of 33.8% between t=1 and t=24, reflecting a narrow window in which these changes within S. bovis could be exploited upon stress (Figure 5D). Deviation across measurements, per time-point, was rather minimal with respect to measurement times. Together the data from these graphs suggest that samples treated under air show more stability with respect to Modulus of Elasticity values.
Discussion
In general, samples tested in the air were magnitudes higher in their Young’s Modulus compared to those probed in PBS (Figure 5). This is easily attributed to drying and is less biomimetic, thus warranting tests in PBS, which has a pH and wetness closer to physiological conditions allowing for softer mechanical conditions. For samples tested in air at 24 hours of antibiotic exposure, S. bovis samples revealed an overall increase in their Young’s Modulus of Elasticity, regardless of antibiotic used, compared to their measurement at brief exposure at t-0 (Figure 5). These reflect an overall hardening of the sample. However, in PBS, these changes did not hold for S. bovis exposed to tylosin phosphate, as its Young’s Modulus was highest at the beginning versus at 24 hours of exposure. Tylosin phosphate might be more effective for halting synthesis of vital cell wall proteins and such occurred before the fixing of these samples; in such, the initial high modulus of elasticity could reflect changes induced by the fixing method versus the modulus of the S. bovis. Virginiamycin and tylosin phosphate are noted to inhibit protein synthesis through targeting and binding to the 50s ribosome, but it’s possible that the S. bovis strain tested might be appreciably resistant to virginiamycin [25, 26, 27, 28, 29]. Protein synthesis occurs in three steps, which is previously described by McDermott, et al. [30]. This allows for antibiotics to target different sites of protein synthesis depending on the type of antibiotic. Virginiamycin and tylosin phosphate fall under two different antibiotic classes. As such, virginiamycin target activity is not completely understood [30]. Thus, its inhibitory activity appears to affect multiple aspects of the ribosome [30]. Misreading of certain mRNA, blocking the formation of the functional initiation complex, and inhibition of translocation has all been observed mode of actions for virginiamycin [30]. Tylosin phosphate prevents protein synthesis by binding to the tRNA binding site on the 50S subunit [30]. This causes the disassociation of molecules from the ribosome [30]. The following work from this could be strengthened through genetic and chemical analysis to biologically validate findings, providing a more complete picture.
Overall, what can be gleaned from this is that mechanical differences can be observed and under the conditions given, they demonstrate that S. bovis samples give varying modulus elasticity values at different time points. This provides difficulty in precisely narrowing down the degree of changes that antibiotics can induce, from this method alone. What is helpful is that this study demonstrates the variance in mechanical loads that exist for samples, under reasonable imaging settings over 24 hours of measurement. Supplementary figures 1 and 2, which respectively show Gauss Model Measurements of Substrates in Air and PBS, reveal relatively reliable measurements, with R^2 values averaging greater than 89% and 95%, respectively. In general, they demonstrate that the contact techniques used were proper, as shown by their bell distribution. Therefore, one undertaking similar studies would be wise to obtain more intermediate measurements to study how their sample changes mechanically over time. Studies taking minimal time points may be hiding flaws in their mechanical characterization.
The extent to which antibiotics do so will require a more interdisciplinary approach that looks at genes linked to resistance, molecular assays examining protein components, as well as less harsh fixative reagents to reduce the blunting of mechanical data. Curious to note would be the changes that affect the bacterial cytoskeleton, as assessing the filamentous proteins within with as little disruption as possible can shed additional light on nanomechanical properties within bacteria [31]. Already, it has been shown that in cytoskeletons in eukaryotic systems impact glucose metabolism – such may be the case within prokaryotic systems as well, which would be worth further investigation [32, 33, 34]. Such a finding may reveal means to attack both structure and cell metabolism simultaneously as cellular elasticity affects reproduction and withstanding physical environmental stresses [34]. Thus, changes to such can reveal mechanisms that can be exploited for therapeutic benefit. These characteristics will be further explored and employed to design a microfluidic device to segregate different bacteria types given that the cytoskeletal elements affect motility as well, which can provide a novel, rapid and economical diagnostic tool in animal husbandry, from farm to farm [31].
As a preliminary work, this paper demonstrates at the least that one can begin probe structural changes in S. bovis in response to changes to antibiotic exposure. To deliver deeper, substantial conclusions, following experiments beyond will need to assay S. bovis properties in increasingly closer, biomimetic conditions. This entails tighter farm partnerships and wider sampling of measurements in times and session modalities. This work will be helpful as projects such as these become more interdisciplinary.
Conclusion, Limitations and Future Work
Key limitations were the lack of characterizing S. bovis from within or after removal from a bovine host. A deficiency in positive and negative lack of positive and negative controls limits the usefulness of this data. It is hoped that a follow up study can provide this data for comparison, as such would surely strengthen the value of what is presented. Key interactions with the host immune system, buffers, extracellular matrices, and proteins are excluded from this study as a result. Such results as listed within this paper give a preliminary look into the behavior and otherwise the existence of S. bovis after exposure to 2 different antibiotics and do not fully reflect S. bovis in nature. Further, the immobilization of the samples with formaldehyde modifies surface proteins, blunting the degree to which mechanical measurements can be utilized, limiting this work to just an exploratory scope. A follow-up study that uses an alternate means can further strengthen this work provided that it does not similarly increase the modulus of elasticity. As it stands, the changes brought by the antibiotics cannot be further confirmed. It is wisely suggested that follow up experiments use a control with only bacteria fixation and without any antibiotic treatment.
Nonetheless, mechanical differences noted still suggested another look and thus merit the publishing of results for those willing to follow up and improve this study. In the meantime, comparison with standards in prior AFM work may be of assistance [9, 10, 11, 12, 13]. This work is overall emphasized as a preliminary work towards carefully examining bacteria exposed to antibiotics. Future work necessitates working more closely with farms to obtain S. bovis samples under closer time points, with an alternative, less disruptive means of immobilizing S. bovis samples. Additionally, such work can open avenues into more closely studying other pathogens and even helpful bacteria. This can lead to therapies designed at reducing bacterial pathogens and facilitating the continued presence of helpful bacteria with less dependence on antibiotics though revealing surface, thermal, and electrical properties that can be exploited. In sum, this work is best understood as an attempt to make as much sense from fixed S. bovis samples, and following or derivative work from his can benefit from improving on sample preparation before imaging, paired with additional intermediate time points and more biologically based screening to con-firm effects of tylosin phosphate and virginiamycin on S. bovis.
Author Contributions: “Conceptualization, D.D. and S.F.; methodology, D.D. and S.F.; validation, D.D., S.F.; formal analysis, X.P., L.M., S.F., and D.D.; investigation, A.B. and D.D.; resources, A.B. and D.D.; data curation, D.D.; writing— original draft preparation, X.P. and L.M.; writing—review and editing, X.P. and L.M.; visualization, D.D.; supervision, D.D.; project administration, D.D.; funding acquisition, D.D. All authors have read and agreed to the published version of the manuscript”.
Funding: The author(s) disclosed receipt of the following financial support for the research, authorship, and/ or publication of this article: The project described was supported by grants from the UNK Undergraduate Research Fellows Program. Use of AFM facility at Nanoimaging Core Facility at University of Nebraska Medical Center (UNMC, Omaha, NE) was supported by Nebraska Research Initiative (NRI) grant, Atomic Force Microscope AFM UNL/UNMC, NRI- 5132103200 (DD).
This work is supported by Animal Nutrition, Growth and Lactation grant no. 2018-67015-27496, Effective Mitigation Srategies for Antimicrobial Resistance grant no. 2018- 68003-27545, and Multi-state research project accession no. 1000579 from the USDA National Institute of Food and Agriculture awarded to SCF.
Data Availability Statement: All data is available upon reasonable request. Acknowledgments: AFM imaging was performed at the UNMC Nanoimaging Core Facility. The Facility is partly supported with funds from the Nebraska Research Initiative (NRI).
Conflicts of Interest: The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
-
Murray Henry W, Roberts RB (1978) Streptococcus Bovis Bacteremia and Underlying Gastrointestinal Disease. Arch Intern Med 138(7): 1097-1099.
-
Corredoira J, Miguez E, Mateo LM, Rodriguez RF, Rodriguez JFG, et al. (2018) Correlation between Streptococcus Bovis Bacteremia and Density of Cows in Galicia, Northwest of Spain. Infection 47(3): 399-407.
-
Tjalsma Harold, Schöller Guinard M, Lasonder E, Ruers TJ, Willems HL, et al. (2006) Profiling the Humoral Immune Response in Colon Cancer Patients: Diagnostic Antigens from Streptococcus Bovis. International Journal of Cancer 119(9): 2127-2135.
-
Brown H, Elliston NG, McAskill JW, Muenster OA, Tonkinson LV (1973) Tylosin Phosphate (TP) and Tylosin Urea Adduct (TUA) for the Prevention of Liver Abscesses, Improved Weight Gains and Feed Efficiency in Feedlot Cattle. Journal of Animal Science 37(5): 1085- 1091.
-
Van Lunen TA (2003) Growth performance of pigs fed diets with and without tylosin phosphate supplementation and reared in a biosecure all-in all-out housing system. Can Vet J 44(7): 571-576.
-
Welton LA, Thal LA, Perri MB, Donabedian S, McMahon J, et al. (1998) Antimicrobial Resistance in Enterococci Isolated from Turkey Flocks Fed Virginiamycin. Antimicrobial Agents and Chemotherapy 42(3): 705- 708.
-
George BA, Quarles CL, Fagerberg DJ (1982) Virginiamycin Effects on Controlling Necrotic Enteritis Infection in Chickens. Poultry Science 61(3): 447-450.
-
Johnson KG, Tyrrell J, Rowe JB, Pethick DW (1998) Behavioural Changes in Stabled Horses given Nontherapeutic Levels of Virginiamycin. Equine Veterinary Journal 30(2): 139-143.
-
Dutta D, Schmidt R, Fernando SC, Dastider IG (2020) A Comparative Study of Force Measurements in Solution Using Micron and Nano Size Probe. World Journal of Nano Science and Engineering 9(1): 1-14.
-
Dutta D, Asmar A, Stacey M (2015) Effects of nanosecond pulse electric fields on cellular elasticity. Micron 72: 15- 20.
-
Dutta D, Palmer XL, Asmar A, Stacey M, Qian S (2017) Nanosecond pulsed electric field induced changes in cell surface charge density. Micron 100: 45-49.
-
Dutta D, Palmer XL, Kim J, Qian S, Stacey M (2018) Energy dissipation mapping of cancer cells. Micron 105: 24-29.
-
Dutta D, Russell C, Kim J, Chandra S (2018) Differential Mobility of Breast Cancer Cells and Normal Breast Epithelial Cells Under DC Electrophoresis and Electroosmosis. Anticancer Res 38(10): 5733-5738.
-
Muller DJ (2008) AFM: a nanotool in membrane biology. Biochemistry 47: 7986-7998.
-
Stolz M, Gottardi R, Raiteri R, Miot S, Martin I, et al. (2009) Early detection of aging cartilage and osteoarthritis in mice and patient samples using atomic force microscopy. Nature Nanotechnology 4(3): 186-192.
-
Zhang CY, Zhang YW (2007) Effects of membrane pre- stress and intrinsic viscoelasticity on nanoindentation of cells using AFM. Philosophical Magazine 87(23): 3415- 3435.
-
Lemoine P, McLaughlin JM (1999) Nanomechanical measurements on polymers using contact mode atomic force microscopy. Thin Solid Films 339(1-2): 258-264.
-
Costa KD, Yin FCP (1999) Analysis of indentation: Implications for measuring mechanical properties with atomic force microscopy. J Biomech Eng 121(5): 462- 471.
-
Liu J, Sandaklie Nikolova L, Wang X, Miller JD (2014) Surface force measurements at kaolinite edge surfaces using atomic force microscopy. Journal of colloid and interface science 420: 35-40.
-
Doktycz MJ, Sullivan CJ, Hoyt PR, Pelletier DA, Wu S, et al. (2003) AFM imaging of bacteria in liquid media immobilized on gelatin coated mica surfaces. Ultramicroscopy 97(1-4): 209-216.
-
Meyer RL, Zhou X, Tang L, Arpanaei A, Kingshott P, et al. (2010) Immobilisation of living bacteria for AFM imaging under physiological conditions. Ultramicroscopy 110(11): 1349-1357.
-
Schaer Zammaretti P, Ubbink J (2003) Imaging of lactic acid bacteria with AFM— elasticity and adhesion maps and their relationship to biological and structural data. Ultramicroscopy 97(1-4): 199-208.
-
Kailas L, Ratcliffe EC, Hayhurst EJ, Walker MG, Foster SJ, et al. (2009) Immobilizing live bacteria for AFM imaging of cellular processes. Ultramicroscopy 109(7): 775-780.
-
Fang HH, Chan KY, Xu LC (2000) Quantification of bacterial adhesion forces using atomic force microscopy (AFM). Journal of microbiological methods 40(1): 89-97.
-
Gadde UD, Oh S, Lillehoj HS, et al. (2018) Antibiotic growth promoters virginiamycin and bacitracin methylene disalicylate alter the chicken intestinal metabolome. Sci Rep 8: 3592.
-
Papich MG (2016) Tylosin. Saunders Handbook of Veterinary Drugs, pp: 826-827.
-
Thal LA Zervos MJ (1999) Occurrence and epidemiology of resistance to virginiamycin and streptogramins. Journal of Antimicrobial Chemotherapy 43(2): 171-176.
-
Butaye P, Devriese LA, Haesebrouck F (2003) Antimicrobial Growth Promoters Used in Animal Feed: Effects of Less Well Known Antibiotics on Gram-Positive Bacteria. Clinical Microbiology Reviews 16(2): 175-188.
-
Catry B, Feyen B, Raeymaekers T, Opsomer G, Timmerman T, et al. (2006) Antimicrobial Resistance in Bovine Commensal Nasal Pasteurellaceae. Semantic Scholar.
-
McDermott PF, Walker RD, White DG (2003) Antimicrobials: modes of action and mechanisms of resistance. Int J Toxicol 22(2): 135-143.
-
Cabeen MT, Jacobs Wagner C (2010) The Bacterial Cytoskeleton. Annu Rev Genet 44: 365-392.
-
Liu Z, Zhang Y, Chang Y, Fang F (2006) The role of cytoskeleton in glucose regulation. Biochemistry (Moscow) 71(5): 476-480.
-
Heaslip AT, Nelson SR, Lombardo AT, Previs SB, Armstrong J, et al. (2014) Cytoskeletal Dependence of Insulin Granule Movement Dynamics in INS-1 Beta- Cells in Response to Glucose. PLoS ONE 9(10): e109082.
-
Guck J, Schinkinger S, Lincoln B, Wottawah F, Ebert S, et al. (2005) Optical Deformability as an Inherent Cell Marker for Testing Malignant Transformation and Metastatic Competence. Biophysical Journal 88(5): 3689-3698.
- Solution-Processed Chiral Perovskites for Biomedical Applications
- Nanotechnology in Health Chemistry and Medicine: Current Challenges and Future Directions
- Human Exposure to Micro- and Nanoplastics: Pathways, Toxicity, and Intervention Strategies
- Exosome Nanomedicine for Cancer Therapy
- Micro and Nanoplastics–Plastisphere, Biotoxicity, Impact on Human Health, and Mitigation Strategies
- Process Validation of Cefixime Powder for Suspension Dosage Form, 50 mL