Role of Mitochondrial Unfolding Protein Response in Reproduction and Aging with Utilization of Autologous Mitochondrial Injections in Mitochondrial Diseases along with in Aging Oocytes
There is a very essential role that mitochondria play in controlling energy metabolism with respect to oocytes maturation as well as Preimplantation embryos in which both quantity as well as morphology of mitochondria along with mitochondrial DNA amount get very closely controlled. Lot of mouse experimental kinds having impaired mitochondrial function cause infertility that validates the crucial part played by mitochondria in female reproduction. On finding abnormalities of mitochondrial action both cells as well as organisms utilize methods that are aimed at recuperating or help in get the mitochondria rescued and delete mitochondria which can’t in anyway be salvaged. Of these methods Mitochondrial Unfolded Protein Response (mtUPR) has been associated with ageing avoidance recently, since mitochondrial aids in age associated collection of injured proteins, decreased oxidative phosphorylation as well as reactive oxygen species (ROS) synthesis. Importance of these lies in female reproduction since if mtUPR genes like Clpp are removed following specific targeting it causes female infertility, causing problems in maturing of oocyte along with 2 cell embryo generation and inability to develop blastocyst. Additionally if CLPP is missing it results in exaggerated follicular removal with a phenotype mimic long premature ovarian failure. More studies will give understanding of both physiological and pathological regulation of oocyte as well as initial embryonic mitochondrial action that workers are trying to use in generating therapies that are innovative in aging oocytes and in women presenting with mitochondrial inheritent abnormalities and thus treating female infertility with elderly females.
Introduction
In all eukaryotic cells practically double membrane organelles named mitochondria exist. Probably their origin started by endosymbiotic process that occurred around 1.5 billion years ago with the eukaryotic cell engulfing bacterium. The size and amount /cell differ, and are separate from them having their own DNA, that encodes 13 proteins having crucial part in mitochondrial function. Only separate organelle possessing its own DNA is the chloroplast seen in the plant cells as well as in eukaryotic algae which carries out photosynthesis. Importantly mitochondrial DNA (mt DNA)
gets inherited via the mother in maximum animals that helps the workers involved in genealogy to track back the maternal line long back within time.
Part of mitochondria is that it is a place from where cells energy originates is well accepted [1, 2, 3, 4, 5, 6]. Besides these complicated organelles aid in a large quantity of key cellular processes that has calcium signalling, apoptosis along with control of membrane potential [4, 7, 8].
Over the last 5 decades, part of mitochondria in aging has been evaluated both in animals as well as human tissues. Lots of probable modes via which mitochondria modulate aging of the cells have been proposed. In the same line, a critical part of mitochondria in both male and female reproduction has been shown in animals and examined in Artificial Reproductive Technology (ART). Thus we decided to conduct a systematic review regarding mitochondrial unfolded protein response (UPRmr), that is a mitochondria stress action path which may participate in female reproduction and reproductive aging.
Methods
We carried out a PubMed search for articles related to mitochondrial changes with relation to aging and abnormalities in reproduction. The MeSH terms mitochondrial unfolding response in various animals in relation to oocytes, aging different genes related to UPRmt, ATFS, CLPP, proteostasis; role of General control repressible 2( GCN2) and eukaryotic initiation factor 2(eIF2), mitochondrial unfolding response in C.elegans and other species.
Results and Discussion
We found a total of 340 articles out of which we utilized 74 articles for this review. No meta-analysis was done.
Oocyte Mitochondria and Mitochondria in Preimplantation Embryos
At the time of the formation of mammalian oocyte as well as early embryo, lot of mitochondria, mt DNA copy number, as well as mitochondrial morphology alters in a manner that can be anticipated, that points to a closely controlled and a very important biological function [9]. Roughly 200 mitochondria are there in oogonia. In view of constant escalation in the amount of mitochondria at the time of follicular formation, mature (metaphaseII) oocytes possess roughly 100,000 mitochondria [10] and 550,000 mt DNA copies with lot of differences among samples [11, 12, 13, 14, 15]. Importantly in-spite of a marked enhancement of the total quantity of mitochondria all through the time of follicular maturation, oocyte mitochondria density (number of mitochondria/cytosol area) and mitochondrial coverage (total mitochondrial area, per cytosol area) reduce with the follicular formation, possibly in view of a fast escalation of oocyte volume [16]. Notably, oocyte is the initial mitochondrial source at the time of initial Preimplantation formation, since mitochondria get inherited maternally and the full amount of mitochondria as well as mt DNA copy numbers continue to be same at the time of fertilization as well as cleavage divisions [14, 17]. Further mt DNA replication is 1st seen in the trophectoderm cells on blastocyst formation that corroborates the marked energy enhancement for meeting the demands of the embryo [18, 19, 20, 21]. On implantation mitochondrial replication also gets restarted [14, 22, 23, 24].
There are a lot of variations in the morphology and anatomical structure of oocyte mitochondria as compared to those observed in somatic cells. Those mitochondria from mature (metaphase II) oocytes are lesser in size (<1µmdiameter), have >spherical shape, and possess lesser as well as truncated cristae that are encircled by a dense matrix [25]. Still, oocyte mitochondria, are working and aid in ATP formation using oxidative phosphorylation, all through oocyte maturation, fertilization, as well as initial cleavage multiplications, at the time when pyruvate is the initial fuel [26, 27, 28, 29]. What is valuable is that the full amount of mitochondria as well as mt DNA copy numbers do not undergo change, dramatic alterations in the mitochondrial morphology take place at the time of initial Preimplantation formation. Greater Elongation of mitochondria occurs (2.51µmdiameter, in 8 cell human embryo) [30], having a > cristae amount with a lower density matrix [18]. This morphologic change gets finished when 1st embryonic cell differentiation takes place that leads to the development of the trophectoderm as well as inner cell mass, during 5-6 days following fertilization in humans [29].
These mitochondrial changes are significant in sustenance of fertility is shown by the phenotypes seen in mouse models where crucial controlling genes are removed by targeting them. Actually, decreased fertility is seen in mice that express defective mitochondrial DNA polymerase [30], as well as in oocyte specific removal of mitochondrial fusion factor Drp1 causes female fertility along with hampered folliculogenesis and ovulation [31]. Recently, global germline removal of mitochondrial stress response gene Clpp has been demonstrated to cause infertility with exaggerated follicular removal [32].
Response of Cells to Mitochondrial Dysfunction
Cells can identify mitochondrial impairment and act by a lot of methods. These actions try to retrieve the function in those mitochondria that can be salvaged and remove those that can’t be saved. Ultimate outcome is the sustainance of a working mitochondrial network and get the homeostasis within the cell back Unfolded Protein Response in Mitochondria: Mitochondrial Unfolded Protein Response (UPRmt) represents transcriptional response at the time of mitochondrial functional impairment which aids in the working of the mitochondrial network as well as prevent cell death. Lot of ways exist by which UPRmt can be coordinated, that are transcription factor ATFS1 (ATF5 in case of mammals), that helps in the expression of nuclear–encoded Mitochondrial chaperones as well as proteases ,reactive oxygen species (ROS), enzymes responsible for detoxification, in addition to Mitochondrial protein import parts[33] (Figure 1). Proteins here get into the cell to sustain the action of healthy organelles and aid in getting back the action in the impaired ones.Role of this UPRmt in aging and reproduction is further detailed.
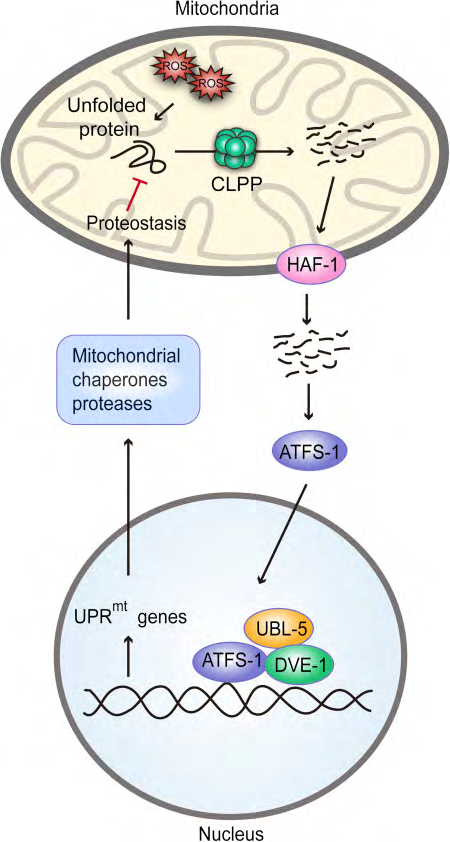
Figure 1: Courtesy ref no-74.Mitochondrial unfolded protein response (UPRmt) characterized in Caenorhabditis elegans, where unfolded or misfolded proteins are degraded by the CLPP protease in the mitochondria and the cleaved products are exported to the cytosol through the HAF-1 peptide exporter. These peptides serve as a signal for activation of transcription factor (ATFS1). Subsequently, ATFS1 activates UBL-5 to form a complex with transcription factor DVE-1 to transcriptionally activate UPRmt genes. This results in the synthesis of mitochondrial chaperones and proteases, reactive oxygen species (ROS) detoxification enzymes, and mitochondrial protein import components, which aim to preserve the function of healthy mitochondria and recover activity in dysfunctional ones.
Repression of Protein Synthesis within the Cytoplasm: Other than 13 of the roughly 1500 Proteins that have a role in the mitochondrial action get formed on the cytosolic ribosomes prior to getting shifted into the mitochondria. Thus these proteins are further acted on by the chaperones present in the mitochondrial matrix. Once the effectiveness of this processing process decreases, protein excess might interfere with the protein homeostasis within the mitochondria also called proteostasis and end in mitochondrial functional impairment. When enhances ROS is present resulting in mitochondrial stress [34, 35], cytoplasmic Protein Synthesis can be decreased by the kinase GCN2 (general control repressible 2) that phosphorylates the eukaryotic initiation factor(eIF2α), that causes reduction of Protein Synthesis and leads to a depression of nascent peptides getting shifted into the mitochondria [36, 37]. This action takes place as role of the integrated stress response and may get stimulated by UPRmt along with different kinds of cellular stress, like endoplasmic reticulum stress.
Mistargeting of Protein Resulting in UPR Activation: Reduced protein shifting efficacy, is a separate outcome of mitochondrial functional impairment that causes collection of wrongly placed mitochondrial Protein within the cytosol. This effect is called activation of UPR by protein mistargeting (UPRam)This leads to breakdown of the abnormally placed proteins having potential to be harmful by resulting in an enhancement of proteasome action and a reduction in Protein Synthesis [38, 39].
Mitophagy: This mitochondrial stress actions detailed earlier try to prevent the injury further in impaired mitochondria and aid in recuperation of those organelles that can be rescued. If these modes don’t work, then as a last attempt mitophagy through which injured mitochondria that are irrepairable get deleted and hence the mitochondrial network gets safeguarded [40]. Usually prior to mitophagy fission alias mitochondrial division leads to badly injured mitochondria parts getting separated from the injured mitochondrial network [41]. For mitophagy to occur PTEN induced putative kinase 1(PINK1) kinase, that in usual situations gets brought into healthy mitochondria from outside and gets destroyed [42, 43]. But in situations which result in disruption of mitochondrial import processes, PINK1 gets stabilized over the outer injured mitochondrial membrane [44]. At that place PINK1 phosphorylates ubiquitin as well as ubiquitin ligase Parkin, that causes the recruiting of Parkin to the outer mitochondrial membrane in which it poly ubiquitinates lot of proteins [45, 46]. This polyubiquitination of proteins on the outer mitochondrial membrane starts the recruiting of the parts required for injured organelles to get swallowed by an autophago some, that further gets moved to lysosomes for breakdown and hence injured mitochondria get expelled [47].
Unfolded Protein Response in Mitochondria: UPRmt
represents a signal transduction pathway via mitochondria to nucleus which makes sure that mitochondrial proteostasis via getting to know the mitochondrial stress or UPR and stimulating a severe transcriptional action which aids in folding, prevents import, and decreases translation of mitochondrial proteins [48, 49, 50]. Multiple mitochondrial stress processes induce UPRmt.
1st delineated in Caenorrhabditis elegans (Figure1), where mitochondrial stress represented unfolded as well as misfolded proteins which up regulated the mitochondrial matrix CLPP protease, that gets cleaving of the misfolded proteins [51, 52, 53]. These proteins that had been cleaved were then sent to the cytoplasm and stimulated the transcription factor ATFS1 that is present within cytoplasm. Further ATFS1 gets transferred to the nucleus where it stimulates ubiquitin liked 5(UBL5) to develop a combination with defective proventriculus in Drosophila homolog) i) as well as for stimulating transcription of chaperones of the mitochondria like heat shock rotein 6(HSP6) and HSP 10 [53, 54, 55, 56]. Additionally UPRmt stimulates coenzyme Q formation, glycolysis as well as mitochondrial fission [30, 54, 57], changing metabolism of mitochondria and its dynamics to aid in its function as well as cell surviving at the time of stress.
UPRmt as well as part of CLPP and ATFS1 (ATF5 has been found to be the mammalian respective part of ATFS1) remains conserved in mammals [30, 48, 51, 55, 58], at the place where stimulation of c-Jun–N terminal kinase takes place. This c-Jun pathway results in the expression of transcription factor CCAAT-enhancer binding protein homologous protein (CHOP) that along with CCAAT-enhancer-binding protein (C/ EBP) modulates the transcription of UPRmt genes [59].
Noticeably, besides stimulating transcription of > 400genes UPRmt) in yeast, Caenorrhabditis elegans and mammals is correlated with phosphorylation of eIF2α by GCN2, leading to globally repressed translation, and mRNA’s which possess open reading frames are translated getting preference [40, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74]. Activation of UPRmt genes by transcription and suppression via translation appears to be carried out by 2 modes which are parallel, with both needing CLPP [30, 48, 51, 53, 55].
Aging and UPRmt
Absence of protein homeostasis or proteostasis appears to have an important part in age associated decrease in function since dysfunctional protein control and collection of misfolded and unfolded proteins have been isolated as a signature part of aging organisms as well as aiding proteostasis via enhanced chaperone expression halting translation, or escalating proteins turnover has been pointed prolong to life span [50, 62].
Current work has suggested that UPRmt is a bridge among proteostasis as well as aging in different organisms. A correlation among UPRmt with long life was 1st demonstrated in 2 long living Caenorrhabditis elegans mitochondrial electron transport chain ETC mutants (isp-1 and clk-1) [62]. If UBL5 or DVE1 (that is modulators of UPRmt are deleted by RNA-interference, it returned the prolongation of life span in both) mutants. In the same line, enhanced long life by muscle particular blockade of ETC complex IV had escalated the expression of UPRmt genes [63]. Notably, a lot of other models having longevity like NAD+/Sirtuin 1 or rapamycin in Caenorrhabditis elegans also needs UPRmt [64].
Total decrease in mitochondrial function is well accepted in aging organisms, and has been demonstrated in yeasts [59], worms [67], flies and mice [54, 69] that interference of mitochondrial ETC action enhances life prolongation [63, 64, 69]. These observations get corroborated by recent findings detailed above, showing that up regulating mitochondrial stress responses adds to escalated life in the ;prolonged lived mitochondrial mutants [63, 67, 70].
If the mitochondrial stress responses get interfered with the lead to age associated accumulation of injured proteins, decreased oxidative phosphorylation .enhanced reactive oxidative stress formation and stimulation of apoptosis of cells. Hence getting an insight into the molecular modes through which mitochondrial stress responses might cause pronation of life might aid in generation of therapies to avoid age associated diseases and thus bettering [49].
Reproduction and UPRmt: Crucial control of UPRmt is done by CLPP. How it works in cleavage of unfolded and misfolded mitochondrial proteins, which then get transferred to cytoplasm and their stimulate the transcription factor ATFS1 remains conserved right from Caenorrhabditis elegans to mammals. Mice developing global germline removal of Clpp showed massive mitochondrial impairment in oocytes having reduced membrane potential as well as ATP generation, along with escalated ROS along with mt DNA copy numbers (Figure 2).
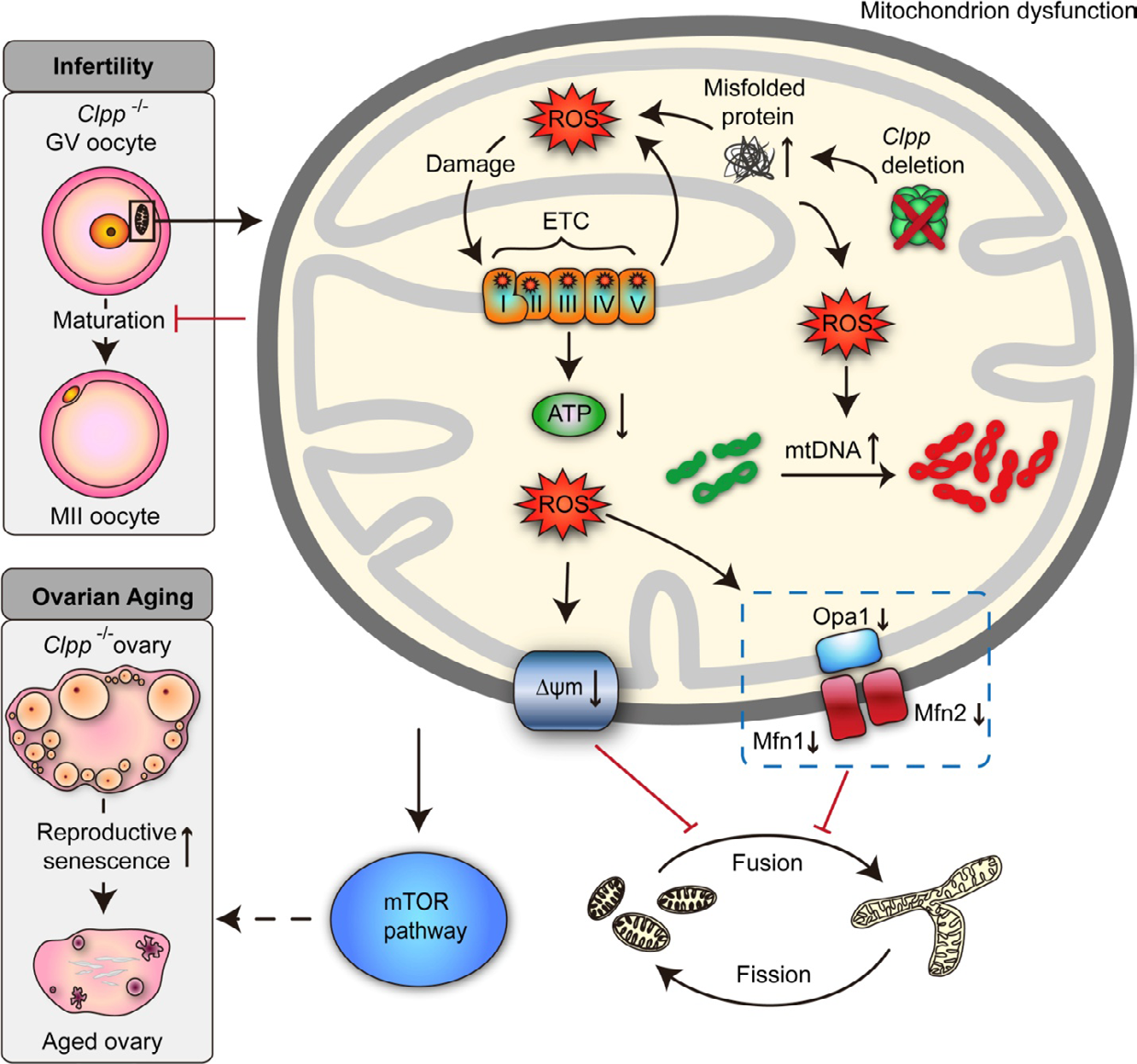
Figure 2: Courtesy reference number 74-A schematic illustration of the effect of Clpp deletion on oocyte mitochondrial function, fertility, and ovarian aging. Targeted global germline deletion of Clpp in the mouse results in mitochondrial dysfunction in oocytes associated with decreased membrane potential and ATP production, and increased reactive oxygen species (ROS) and mitochondrial DNA (mtDNA) copy number. These changes are associated with decreased expression of mitochondrial fusion genes (Mfn1, Mfn2, and Opa1), resulting in smaller and rounder mitochondria. Clpp-knockout mice are infertile with impaired oocyte maturation and inability to form blastocysts, and they show accelerated follicular depletion, associated with activation of mammalian target of rapamycin (mTOR) signaling.
Further metabolic imaging utilizing fluorescence lifetime imaging microscopy showed important changes in the metabolism of Clpp-decreased oocytes, along with alterations in flavin adenine dinucleotide (FAD) and nicotinamide adenine dinucleotide (NAD) lifetimes as well as protein bound part of NAD H [71]. Noticeably mitochondrial dynamics like fusion as well as fission were further afflicted: down regulation of fusion genes, causing rounder and small sized mitochondria [32]. In females with Clpp-decreased mice, infertility resulted from dysfunctional oocyte maturation, 2 cell embryo generation as well as absence of blastocyst formation [32]. These were correlated with marked spindle problems within oocytes. Biggest baffling finding was the exaggerated deletion of Clpp-decreased follicles in, mice; primordial follicles reduced by 1/3rd over 6mths and 50% by 9mths as compared to controls [32]. These alterations correlated with stimulation of mammalian target of rapamycin (mTOR) pathway, with the reproductive phenotype getting partially retrieved utilizing mTOR inhibitor rapamycin.
Management
In humans involves autologous mitochondrial transfer [72] for techniques–reviewed in detail-after mitochondrial transfer, the quantity of ATP needed for old oocytes at the time of fertilization, blastocyst development and further embryonic formation might be another method (Figures 3 and 4 for examples). Mitochondrial transfer from ovarian cells as well as healthy oocytes might result in better fertility results in case of oocytes that are of poor quality. Worldwide trials of AUGMENT technique are going on [73]. Manipulation of mitochondrial activity appears to control the basal metabolism within the target oocytes and thus physiological action of these cells when attempting to overtake age associated fertility in female germ cells [72].
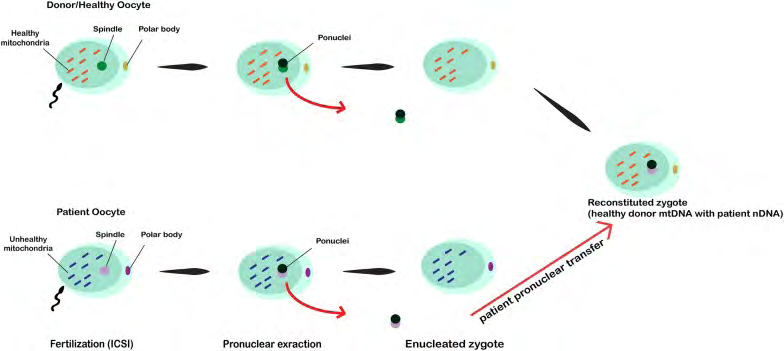
![Figure 4: In this technique, the spindle from defective oocytes is extracted and microinjected into donor healthy oocytes in which the spindle is removed. After fertilization by ICSI procedure, the zygote is reconstituted [72].](/fulltextimages/4876/fig_4.png)
Conclusion
Mitochondrial Unfolded Protein Response represents a mitochondrial stress response. Once stimulated this UPRmt following mild to moderate alterations regarding Mitochondrial action causes extension of the total life span in various animal kinds. On the other hand dysfunctional UPRmt contributes to age associated collection of injured proteins, decreases oxidative phosphorylation, and enhances generation of ROS. Importance of these lies in relation to reproduction since if one removes germline UPRmt regulatory protein CLPP globally it causes infertility and exaggerated follicular removal which is the initial phenotype properties. Part restoration of fertility phenotype in Clpp-deficient mice with rapamycin the mTOR inhibitor is further emphasized. This importance of studies are being carried out with worldwide AUGMENT trials regarding the use of this in improving fertility in older women.
References
-
Cannon B, Nedergaard J (2004) Brown adipose tissue:Function and physiological significance. Physiol Rev 84(1): 277-359.
-
Enerbäck S, Jacobsson A, Simpson EM, Guerra C, Yamashita H, et al. (1997) Mice lacking mitochondrial uncoupling protein are cold sensitive but not obese. Nature 387(6628): 90-94.
-
Ricquier D, Bouillaud F (2000) Mitochondrial uncoupling protein: from mitochondria to the regulation of energy balance. J Physiol 529: 3-10.
-
Andrews ZB, Diano S, Horvath TL (2005) Mitochondrial uncoupling proteins in the CNS: in support of function and survival. Nat Rev Neurosci 6(11): 829-840.
-
Krauss S, Zhang CY, Lowell BB (2005) The Mitochondrial uncoupling protein homologues. Nat Rev Mol Cell Biol 6(3): 248-261.
-
Andrews ZB, Horvath TL, Barnstable CJ, Elsworth J, Yang L, et al. (2005) Uncoupling protein-2 is critical for nigral dopamine cell survival in a mouse model of Parkinson’s disease. J Neurosci 25(1): 184-191.
-
Finkel T (2012) Signal transduction by Mitochondrial oxidants. J Biol Chem 287(7): 4434-4440.
-
Wang C, Youle RJ (2009) The role of Mitochondria in apoptosis. Annu Rev Genet 43: 95-118.
-
Seli E (2016) Mitochondrial DNA as a biomarker for in vitro fertilization outcome. Curr Opin Obstet Gynaecol 28(3): 158-163.
-
Eichenlaub-Ritter U, Wieczorek M, Lüke S, Seidel T (2011) Age related changes in Mitochondrial function and new approaches to study redox regulation in mammalian oocytes in response to age or maturation conditions. Mitochondrion 11(5): 783-796.
-
Steuerwald N, Barritt JA, Adler R, Malter H, Schimmel T, et al. (2000) Quantification of mt DNA in single oocytes, polar bodies and subcellular components by real time rapid cycle fluorescence monitored PCR. Zygote 8(3): 209-215.
-
Reynier P, May-Panloup P, Chrétien MF, Morgan CJ, Jean M, et al. (2001) Mitochondrial DNA content affect the fertizeability of human oocytes. Mol Hum Reprod 7(5): 425-429.
-
Chen X, Prosser R, Simonetti S, Sadlock J, Jagiello G, et al. (1995) Rearranged Mitochondrial genomes are present in human oocytes. Am J Hum Genet 57(2): 239-247.
-
Piko L, Taylor KD (1987) Amounts of Mitochondrial DNA and abundance of some Mitochondrial genes transcripts in early mouse embryos. Dev Biol 123(2): 364-374.
-
Chan CC, Liu VW, Lau EY, Yeung WS, Ng EH, et al. (2005) Mitochondrial DNA content and 4977 bp deletion in unfertilized oocytes. Mol Hum Reprod 11(12): 843-846.
-
Babayev E, Wang T, Szigeti-Buck K, Lowther K, Taylor HS, et al. (2016) Aging is associated with changes in Mitochondrial dynamics, function and mtDNA quantity. Mauritias 93: 121-130.
-
Piko L, Matsumoto L (1976) Number of Mitochondria and some properties of Mitochondrial DNA in the mouse egg. Dev Biol 49(1): 1-10.
-
Van Blerkom J (2011) Mitochondrial function in the human oocytes and embryos and the role in developmental competence. Mitochondrion 11(5): 797- 813.
-
Houghton F (2006) Endergy metabolism of the inner cell mass and trophectoderm of the mouse blastocyst. Differentiation 74(1): 11-18.
-
St John J (2014) The control of mt DNA replication during Differentiation and development. Biochim Biophys Acta 1840(4): 1345-1354.
-
St John JC, Facucho-Oliveira J, Jiang Y, Kelly R, Salah R (2010) Mitochondrial DNA transmission, replication, and inheritance: a journey from the gamete through the embryo and embryonic stem cells. Hum Reprod Update 16(5): 488-509.
-
Murakoshi Y, Sueoka K, Takahashi K, Sato S, Sakurai T, et al. (2013) Embryo developmental capability and pregnancy outcome are related to the Mitochondrial DNA copy numbers and oocyte volume. J Asst Reprod Genet 30(10): 1367-1375.
-
Thundathil J, Filion F, Smith LC (2005) Molecular control of Mitochondrial function in Preimplantation mouse embryos. Mol Reprod Dev 71(4): 405-413.
-
Spikings EC, Alderson J, St John JC (2007) Regulated Mitochondrial DNA replication during oocyte maturation is essential for successful porcine embryonic development. Biol Reprod 76(2): 327-335.
-
Motta PM, Nottola SA, Makabe S (2000) Mitochondrial morphology in human fetal and adult female germ cells. Hum Reprod 15(S2): 129-147.
-
Van Blerkom J, Davis PM, Lee J (1995) ATP content of human oocytes and developmental potential and outcome after in vitro fertilization and embryo transfer. Hum Reprod 10(2): 415-424.
-
Dumollard R, Duchen M, Carrol J (2007) The role of Mitochondrial function in the oocyte and the embryo. Curr Top Dev Biol 77: 21-49.
-
Scott RR, Zhang M, Seli E (2018) Metabolism of the oocytes and the Preimplantation embryos:implications for assisted Reproduction. Curr Opin Obstet Gynaecol 30(3): 163-170.
-
Sathananthan AH, Tounson AD (2000) Mitochondria morphology during Preimplantation human embryogenesis. Hum Reprod 15(S2): 148-159.
-
Aldridge JE, Honbe T, Hoogenraad NJ (2007) Discovery of genes activated by the Mitochondrial Unfolded Protein Response (mtUPR) and cognate promoter elements. PLoS One 2: e874.
-
Udagawa O, Ishihara T, Maeda M, Matsunaga Y, Tsukamoto S, et al. (2014) Mitochondrial fission factor Drp1 maintains oocyte quality via dynamic rearrangement of multiple organelles. Curr Biol 24(20): 2451-2458.
-
Wang T, Babayev E, Jiang Z, Li G, Zhang M, et al. (2018) Mitochondrial Unfolded Protein Response gene Clpp is required to maintain ovarian follicular reserve during aging, for oocyte competence, and development of Preimplantation embryos. Aging Cell 17(4): e12784.
-
Nargund AM, Fiorese CJ, Pellegrino MW, Deng P, Haynes CM (2015) Mitochondrial and nuclear accumulation of the transcription factor ATFS1 promotes OXPHOS recovery during the UPR(mt). Mol Cell 58(1): 123-133.
-
Baker BM, Nargund AM, Sun T, Haynes CM (2012) Protective coupling of Mitochondrial function and Protein Synthesis via the eIF2α kinase GCN2. PLoS Genet 8(6): e1002760.
-
Ishimura R, Nagy G, Dotu I, Chuang JH, Ackerman SL (2016) Activation of GCN2 kinase by ribosome stalling links translation elongation with translation initiation. Elife 5: e14295.
-
Harding HP, Zhang Y, Zeng H, Novoa I, Lu PD, et al. (2003) An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol Cell 11(3): 619-633.
-
Dey M, Cao C, Dar AC, Tamura T, Ozato K, et al. (2005) Mechanistic link between PKR dimerization, auto phosphorylation, and eIF2α substrate recognition. Cell 122(6): 901-913.
-
Wrobel L, Topf U, Bragoszewski P, Wiese S, Sztolsztener ME, et al. (2015) Mistargeted Mitochondrial protein activates a proteostatic response in the cytosol. Nature 524(7566): 485-488.
-
Wang X, Chen XJ (2015) A cytosolic network suppressing Mitochondria mediated proteostatic stress and Cell death. Nature 524: 481-484.
-
Ashrafi G, Schwartz TL (2013) The pathway of mitophagy for quality control and clearance of Mitochondria. Cell death Differ 20(1): 31-42.
-
Twig G, Elorza A, Molina AJ, Mohamed H, Wikstrom JD, et al. (2008) Fission and selective fusion govern Mitochondrial segregation and elimination by autophagy. EMBO J 27(2): 433-446.
-
Narendra DP, Jin SM, Tanaka A, Suen DF, Gautier CA, et al. (2010) PINK1 is selectively stabilized on impaired Mitochondria to activate Parkin. PLoS Biol 8(1): e1000298.
-
Valente EM, Abou-Sheiman PM, Caputo V, Muqit MM, Harvey K, et al. (2004) Herediatary early -onset Parkinson’s Disease caused by mutations in PINK1. Science 304(5674): 1158-1160.
-
Koyano F, Okatsu K, Kosako H, Tamura Y, Go E, et al. (2014) Ubiquitin is phosphorylated by PINK1 to activate Parkin. Nature 510(7503): 162-166.
-
Kazlauskaite A, Kondapalli C, Gourlay R, Campbell DG, Ritorto MS, et al. (2014) Parkin is activated by PINK1 dependent phosphorylation of Ubiquitin by Ser65. Biochem J 460(1): 127-139.
-
Sarraf SA, Raman M, Guarani-Pereira V, Sowa ME, Huttlin EL, et al. (2013) Landscape of the Parkin- dependent Ubiquitinylome in response Mitochondrial depolarization. Nature 496(7445): 372-376.
-
Heo JM, Ordureau A, Paulo JA, Rinehart J, Harper JW (2015) The PINK1- Parkin Mitochondrial Ubiquitinylation pathway drives a program of OPTN/DP52 recruitment and TRK1 activation to promote mitophagy. Mol Cell 60(1): 7-20.
-
Zhao Q, Wang J, Levichkin IV, Stasinopoulos S, Ryan MT, et al. (2002) A Mitochondrial specific stress response in mammalian cells . EMBO J 21(17): 4411-4419.
-
Hill S, Van Remmen H (2014) Mitochondrial stress signalling in longevity: a new role for Mitochondria in aging Redox Biol 2: 936-944.
-
Jensen MB, Jasper H (2014) Mitochondrial proteostasis in the control of aging and longevity. Cell Metab 20(2): 214-225.
-
Benedetti C, Haynes CM, Yang Y, Harding HP, Ron D (2006) Ubiquitin like protein 5 positivitely regulates chaperone gene expression in the Mitochondrial Unfolded Protein Response. Genetics 174(1): 229-239.
-
Martinus RD, Garth GP, Webster TL, Cartwright P, Naylor DJ, et al. (1996) Selective induction of Mitochondrial chaperones in Response to loss of the Mitochondrial genome. Eur J Biochem 240(1): 98-103.
-
Haynes CM, Petrova K, Benedetti C, Yang Y, Ron D (2007) ClpP mediates activation of a Mitochondrial Unfolded Protein Response in C.elegans. Dev Cell 13(4): 467-480.
-
Nargund AM, Pellegrino MW, Fiorese CJ, Baker BM, Haynes CM (2012) Mitochondrial import efficiency of ATFS1 regulates Mitochondrial UPR activation. Science 337(6094): 587-590.
-
Rath E, Berger E, Messlik A, Nunes T, Liu B, et al. (2012) Induction of ds RNA –activated protein kinase links Mitochondrial Unfolded Protein Response to the pathogenesis of intestinal inflammation. Gut 61(9): 1269-1278.
-
Haynes CM, Yang Y, Blais SP, Neubert TA, Ron D (2010) The matrix peptide exporter HAF-1 signals a Mitochondrial UPR by activating the transcription factor ZC376.7 in C.elegans. Mol Cell 37(4): 529-540.
-
Cristina D, Cary M, Lunceford A, Clarke C, Kenyon C (2009) A regulated response to impaired respiration slows behavioral rates and increases life span in Caenor- habditis elegans. PLoS Genet 5: e1000450.
-
Fiorese CJ, Schultz AM, Lin YF, Rosin N, Pellegrino MW, et al. (2016) The transcription factor ATFS1 mediates a mammalian Mitochondrial UPR. Curr Biol 26(15): 2037- 2043.
-
Delaney JR, Ahmed U, Chou A, Sim S, Carr D, et al. (2013) Stress profiling of longevity mutants identifies Afg3 as a mitochondrial determinant of cytoplasmic mRNA translation and aging. Aging Cell 12(1): 156-166.
-
Michael S, Cannone M, Arnold T, Renard P (2015) Inhibition of Mitochondrial genome expression triggers the activation of CHOP-10 by a cell signalling dependent on the integrated stress responses but not the Mitochondrial Unfolded Protein Response. Mitochondrion 21: 58-68.
-
Taylor RC, Dillin A (2011) Aging is an event of proteostasis collapse. Cold Spring Harb Perspect Biol 3(5): 1-17.
-
Durleaux J, Wolff S, Dillin A (2011) The cell –non autonomous nature of electron transport chain mediated longevity. Cell 144(1): 79-81.
-
Pulliam DA, Deepa SS, Liu Y, Hill S, Lin AL, et al. (2014) Complex IV-deficient Surf-/- mice initiate Mitochondrial stress responses. Biochem J 462(2): 359-371.
-
Owusu-Ansah E, Song W, Perrimon N (2013) Muscle Mitohormesis promotes longevity via systemic repression of insulin signalling. Cell 155(3): 699-712.
-
Mouchiroud L, Houtkooper RH, Moullan N, Katsyuba E, Ryu D, et al. (2013) The NAD-Sirtuin pathway modulates longevity through activation of Mitochondrial UPR and FOXO signalling. Cell 154(2): 430-441.
-
Lopez-Otin C, Blasco M, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153(6): 1194- 1217.
-
Dillin A, Hsu AL, Arantes-Oliveira N, Lehrer-Graiwer J, Hsin H, et al. (2002) Rates of behavior and aging specified by Mitochondrial function during development. Science 298(5602): 2398-2401.
-
Liu X, Jiang N, Hughes B, Bigras E, Shoubridge E, et al. (2005) Evolutionary conversation of the clk1-dependent mechanism of-longevity:loss of the mclk1 increases cellular fitness and lifespan in mice. Genes Dev 19(20): 2424-2434.
-
Lapointe J, Hekimi S (2008) Early Mitochondrial dysfunction in long lived Mclk+/-mice. J Biochem Chem 283(38): 26217-26227.
-
Kirchman PA, Kim S, Lai CY, Jazwinski SM (1999) Interorganelle signalling is a determinant of longevity in Saccharomyces cerevisiae. Genetics 152(1): 179-190.
-
Sanchez-T, Wang T, Pedro MV, Zhang M, Esencan E, et al. (2018) Metabolic imaging using fluorescence lifetime imaging microscophy (FUM) accurately detects Mitochondrial dysfunction in mouse oocytes. Fertil Steril 110(7): 1387-1397.
-
Mobarak H, Heidarpour M, JasonTsai PS, Rezabakhsh A, Rahbarghazi R, et al. (2019) Autologous mitochondrial microinjection;a strategy to improve the oocyte quality and subsequent reproductive outcome during aging. Cell Biosci 9: 95.
-
Ishii T, Hobino Y (2018) Mitochondrial manipulation in fertility clinics:regulation and responsibility. Reprod Biomed Soc Online 5: 93-109.
-
Seli E, Wang T, Horvath TL (2019) Mitochondrial Unfolded Protein Response:A stress response with implications for fertility and reproductive aging. Fertil Steril 111(2): 197-204.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’