Utilizing the Knowledge of Role of Matrix Matrixmetalloproteinases Along with Tissue Inhibitors of Matrix Metalloproteinasesin Various Gynaecological Disorders with Great Emphasis on Preeclampsia, PCOs Along with Gynaecological Malignancies-A Systematic Review
Our insight has escalated over the past decade regarding the understanding of Molecular mechanisms underlying the generation of preeclampsia, with us reviewing the role of epigenetics in the development of preeclampsia along with intra uterine growth restriction (IUGR) generation. Matrix Metalloproteinases (MM’s) MMP’s represent calcium-based zinc-endopeptidases that belong to the metzincin superfamily possessing a lot of functions like a physiological part in tissue morphogenesis, cell migration along with angiogenesis as well as in pregnancy in invasion along with migration of trophoblast besides spiral artery remodeling. Matrix Metalloproteinases (MMP), cytokines, growth factors have a role in physiological events like embryo implantation, trophoblast invasion as well as migration along with decidualization with studies documented that blastocysts are direct generators of MMP2. Thus here we decided to conduct a systematic review on the role of Matrix Metalloproteinases (MMP’s) along with their inhibitors,namely the, Tissue inhibitors of Matrix Metalloproteinases (TIMPs) in gynaecological disorders specifically in generation of preeclampsia along with IUGR; in women with Polycystic Ovarian Syndrome(PCOS); besides in spontaneous abortion along with generation of various cancers of the whole body but main emphasis on gynaecological cancers .Hence utilizing the search engine Thus we utilized Pubmed, MEDLINE, Google Scholar, Scopus, Embase, Web of Sciences, Cochrane library search engines where we utilized the MeSH terms like MMP’s in pregnancy physiological roles; pathological in preeclampsia, IUGR; miRs in these disorders; spontaneous abortion; gynaecological disorders like PCOS; gynaecological cancer along with association of certain MIR’s abnormal in IUGR that were linked to certain gliomas along with MMP’S and IUGR and further utilization of MMP inhibitors clinically, besides that since no efficacious therapy available other therapies like role of extracellular vesicles (ECV’s) that target combined miR’S that are altered and associated with MMP’s in various pathologies and effectively we can treat those disorders from 1970 till January 2021 till date. We found a total of 500 articles out of which we selected 145 articles for this review following evaluation of cross references. No meta-analysis was done. Here we provide an insight on how this knowledge on molecular mechanisms interactions of MMP’s, TIMP’s micro RNA’S might get utilized in generating therapies to prevent or treat preeclampsia, IUGR, various gynaecological malignancies developing extracellular vesicles carrying miR;s targeting specific MMP’s in particular disease or malignancy.
Introduction
MMPs represent a family of zinc-based endoproteases possessing lot of parts in tissue refashioning and breakdown of different proteins in the extracellular matrix (ECM). MMPs facilitate cell proliferation, migration, along with differentiation besides playing a part in cell apoptosis, angiogenesis, tissue repair, along with immune response. MMPs might further influence bioactive molecules on the cell surface and manipulate different cellular in addition to signaling pathways. Changes in MMP expression in addition to activity take place in normal biological events like during pregnancy , wound healing, nevertheless, have also been seen in cardiovascular diseases like atherosclerosis, aneurysms and varicose veins, musculoskeletal disorders such as osteoarthritis and bone resorption, and in various cancers. MMPs have also been believed to be involved in tumor propragation along with invasiveness.
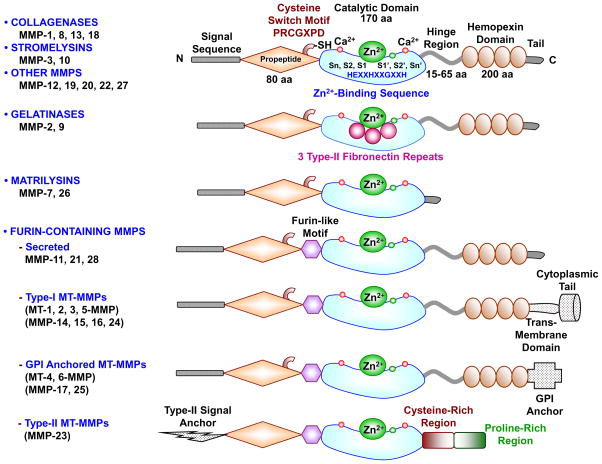
Figure 1: Courtesy reference no-1-Major MMPs subtypes and their structure. A typical MMP consists of a propeptide, a catalytic metalloproteinase domain, a linker peptide (hinge region), and a hemopexin domain. The propeptide has a cysteine switch PRCGXPD whose cysteine sulfhydryl (–SH) group chelates the active site Zn2+, keeping the MMP in the latent pro MMP zymogen form. The catalytic domain contains the Zn2+ binding motif HEXXHXXGXXH, two Zn2+ ions (one catalytic and one structural), specific S1, S2,…Sn and S1′, S2′,…Sn′ pockets, which confer specificity, and two or three Ca2+ ions for stabilization. Some MMPs show exceptions in their structures. Gelatinases have 3 type-II fibronectin repeats in the catalytic domain. Matrilysins have neither a hinge region nor a hemopexin domain. Furin-containing MMPs such as MMP-11, 21 and 28 have a furin-like pro- protein convertase recognition sequence in the propeptide C-terminus. MMP-28 has a slightly different cysteine switch motif PRCGVTD. Membrane-type MMPs (MT-MMPs) typically have a transmembrane domain and a cytosolic domain. MMP-17 and -25 have a glycosylphosphatidylinositol (GPI) anchor. MMP-23 lacks the consensus PRCGXPD motif, has a cysteine residue located in a different sequence ALCLLPA, may remain in the latent inactive proform through its type-II signal anchor, and has a cysteine-rich region and an immunoglobulin-like proline-rich region.
Classically, MMPs possess a propeptide sequence, a catalytic metalloproteinase domain along with catalytic zinc, a hinge region or linker peptide, besides a hemopexin domain. Usually classification of MMPs is done depending on their substrates along with the organization of their structural domains into collagenases, gelatinases, stromelysins, matrilysins, membrane-type (MT)-MMPs, and other MMPs. MMPs are liberated by lot of cells that include fibroblasts, vascular smooth muscle (VSM) and leukocytes. MMPs are controlled at the level of mRNA expression and by activation of their latent zymogen form. MMPs are often liberated as inactive pro MMP form which is cleaved to the active form by various proteinases that are other MMPs. MMPs result in the breakdown of ECM proteins like collagen along with elastin, nevertheless, could affect endothelial cell function in addition to VSM cell migration, proliferation, Ca2+ signaling along with contraction. MMPs participate in tissue remodeling at the time of different physiological events like angiogenesis, embryogenesis, morphogenesis along with wound repair, besides in pathological conditions like myocardial infarction, fibrotic disorders, osteoarthritis, along with cancer. Escalation in specific MMPs could participate in arterial remodeling, aneurysm generation, venous dilation along with lower extremity venous disorders. MMPs further participate in leukocyte infiltration along with tissue inflammation. MMPs have been documented in cancer, and enhanced MMP amounts have been correlated with tumor propagation along with invasiveness. MMPs can be controlled by endogenous tissue inhibitors of metalloproteinases (TIMPs), and the MMP/TIMP ratio often determines the extent of ECM protein brealdown along with tissue remodeling. MMPs have been posited to be biomarkers for a lot of pathological conditions along with getting evaluated as potential therapeutic targets in different cardiovascular (CVS) along with musculoskeletal disorders in addition to cancer [1] (Figures 1 & 2). Thus here we decided to conduct a systematic review on the role of Matrix Metalloproteinases (MMP’s) along with their inhibitors, namely the, Tissue inhibitors of Matrix Metalloproteinases (TIMPs) in gynaecological disorders specifically in generation of preeclampsia along with IUGR; in women with Polycystic Ovarian Syndrome(PCOS); besides in spontaneous abortion along with generation of various cancers of the whole body but main emphasis on gynaecological cancers.
Methods
Hence utilizing the search engine we utilized Pubmed, MEDLINE, Google Scholar, Scopus, Embase, Web of Sciences, Cochrane library search engines we utilized the MeSH terms like MMP’s in pregnancy physiological roles; pathological in preeclampsia, IUGR; miRs in these disorders; spontaneous abortion; gynaecological disorders like PCOS; gynaecological cancer along with association of certain miR’s abnormal in IUGR that were linked to certain gliomas along with MMP’S and IUGR and further utilization of MMP inhibitors clinically, besides that since no efficacious therapy available other therapies like role of extracellular vesicles (ECV’s) that target combined miR’S that are altered and associated with MMP’s in various pathologies and effectively we can treat those disorders from 1970till January 2021 till date.
Results
We found a total of 500 articles out of which we selected 145 articles for this review following evaluation of cross references. No meta-analysis was done.
Matrix Metalloproteinases (MMP’s), Tissue Inhibitors of Matrix Metalloproteinases (TIMPS) as well as Gynaecological Abnormalities
Metalloproteinases and TIMPs are normally expressed in variety of tissues. Nevertheless, it has been documented that at the time of generation of variety of human diseases, metalloproteinases 1, 2, 3, 7, 8, 9,13, and 14 get overexpressed in particular tissues. These tissues are the kidneys, liver, colon, placenta, intestines, stomach, bladder, pancreas, ovary, uterus, along with bone marrow, among others. TIMPs have varying expression in particular tissues like the breast, brain, lungs, liver, kidneys, colon, skin, ovaries, and heart. In these tissues, TIMP-1 is overexpressed, TIMP-3 is under expressed, and TIMP-2 and TIMP-4 can be either under expressed or over expressed [2] Figure 3.
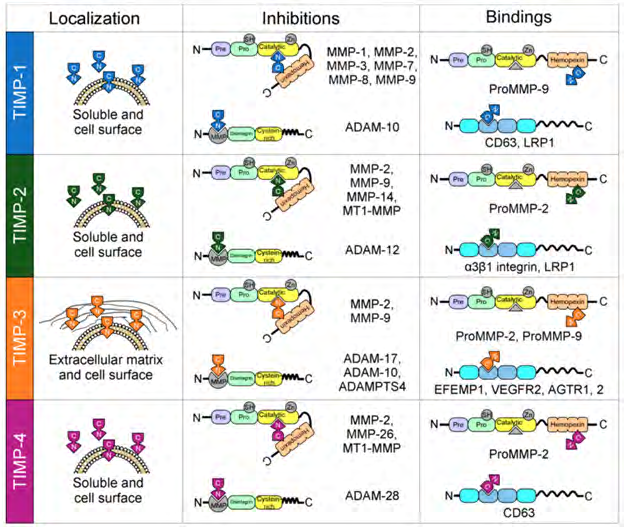
Figure 3: Courtesy ref no-2-Localization and interactions between tissue inhibitors of matrix metalloproteases (TIMPs) and matrix metalloproteases (MMPs). All TIMPs are secreted, but only TIMP-3 is incorporated into the matrix. Structurally, TIMPs are comprised of two domains that pack side-by-side (N-terminal and C-terminal domains). The N-terminal domain is sometimes referred to as the “inhibitory domain”. TIMP-1 inhibits MMP-1–3 and MMP-7–9; TIMP-2 inhibits MMP-2, MMP-9, MMP-14, and membrane-type matrix metalloproteases 1 (MT1-MMP); TIMP-3 inhibits MMP-2 and MMP-9; and finally, TIMP-4 inhibits MMP-2, MMP-26, and MT1-MMP. In addition, TIMPs interact with the proforms of MMPs in a non-inhibitory manner, and they also have functions independent of MMP inhibition by directly binding to cell surface receptors (TIMP-1 to CD63; TIMP-2 to α3β1integrin and LRP1; TIMP-3 to EFEMP1, VEGFR2 and AGTR1,2; and TIMP-4 to CD63).
Matrix Metalloproteinases (MMP), cytokines, growth factors have a role in physiological events like embryo implantation, trophoblast invasion as well as migration along with decidualization with studies documented that blastocysts are direct generators of MMP2 [3]. Of the cytokines implicated in decidualization, IL-11,that is generated by stromal as well as epithelial cells along with escalated by IL--1α, Tumor necrosis factor alpha ( TNF α),transforming growth factorβ(TGFβ), that get stimulated the generation of human endometrium [4]. Besides the implicated cytokines, escalated amounts of MMPs, like MMP-2, MMP-3 as well as MMP-9 that get expressed in the deciduas are correlated with an enhancement of trophoblast invasion [5].The existence of MMPs In addition to gonadotropin chorionic hormone as well as VEGF, aids in manipulations of the endometrium which are needed for human implantation to take place, In addition to a part in the avoidance of aberrant placental generation of the placenta [5, 6].
MMP, along with TIMP regulate the area along with refashioning of polycystic ovarian tissue, besides being correlated with a lot of physiological as well as pathological events. The appropriate balance among MMPs as well as their inhibitors have a big part in the structural along with functional vascular alterations of women possessing complicated pregnancies [7].
Polycystic Ovarian Syndrome (PCOS)
PCOS represents the commonest condition of endocrine initiation in women from reproductive age. Its clinical expression usually are oligo or anovulation, hyperandrogenism (clinical/biochemical) along with the existence of polycystic ovaries In addition to its association with insulin resistance(IR), Type 2 diabetes mellitus (T2DM), obesity, cardiovascular disease (CVD), along with escalated cancer chances [8]. PCOS represents the commonest etiologies of anovulatory infertility, implicating 6-10% of premenopausal women. Adrenal In addition to pituitary diseases need to be ruled out prior to diagnosing it along with the counting of primordial follicles is further significant [9].It has significant incidence in younger women, as well as might get underdiagnosed in view of greater amount of follicles. Although its etiology is not fully clarified, proof exists that a genetic factor which influences patients at the time of oogenesis [10]. The escalated amount of androgens In addition to LH generation, that represses the generation of SHBG, along with stimulation of adrenal as well as androgens generation correlated with anovulatory cycles [11]. Evaluation of MMP-2 along with MMP-9 expression in the ovaries was conducted in certain studies [12, 13]. Follicular atresia In addition to generation of follicular cysts is the hallmark of histological properties of PCOS. MMP-9 is associated with an imbalance in the escalated reconstruction of the ovarian follicles as well as walls, resulting in irregular ovulation, Follicular occlusion In addition to escalated stromal tissue [14]. TIMP-1, TIMP-2 as well as TIMP-3 are present in the stroma along with teak of developing follicles. Besides serum amounts of MMP-9, MMP-9: TIMP-1 ratios are significantly > in women with PCOS as compared to healthy women [15]. Changes in balance among serum amounts of MMPs as well as TIMP in women with PCOS has got documented recently .Implying that a reduction in serum TIMP amounts, along with proportions of MMP-9 to TIMP-1 as well as MMP-2 to TIMP-2 escalate significantly [16]. TIMP-1 amounts might specifically depict both systemic chronic inflammatory along with immune responses. Escalated amounts of MMP-8, along with MMP-8: TIMP-1 ratio in saliva along with serum appear to be more accentuated in women with PCOS, In addition to getting augmented by gingival inflammation [17]. Besides that studies have documented that testosterone(T) amounts associated positively with the percentage of MMP-9 to TIMP- 1 besides correlating negatively with TIMP-2 [18].One can presume that enhanced androgens liberation can change the balance of MMPs , along with TIMP in the ovaries under physiological situations ,causing propagation of fibrosis in women with PCOS [19]. Suppression of the MMP-9: TIMP- 1 ratios might have a significant manipulative actions on progesterone (P) liberation [20]. Conversely, other studies have demonstrated that the amounts of TIMP-3 mRNA is significantly less in PCO ovaries. These changes seen in the liberation as well as/or distribution of type IV collagen along with TIMP-3point the implication of basement membrane (BM) in the pathogenesis of PCOS [21]. Escalated amounts of TIMP-1 have got observed in PCOS patients, as well as women who get pregnant possess greater amounts of TIMP-1 [22].
Spontaneous Abortion
Spontaneous Abortion (SA) by definition is the expelling of fetal products prior to getting viable at the gestation age (GA) of 22 wks or at the time the fetus weigh s 50gm (usually among the 6th-10th wk of pregnancy [23]. Maximum SAs take place in women among 20-45 yrs age, with the peak occurring at the ages of 40-44 yrs (90%) [24].
The basic etiology of SA are maternal, ovofetal along with implantation factors [25]. As far is implantation is considered, the early loss of fetus gets enhanced when implantation takes >10days, but other 20% of cases take place once trophoblast refuses to implant. The commonest issues taking place among 0-10 wk GA associate with fertilization or malformations in the fetus secondary to chromosomal aberrations [26]. There are disorders that stop the right generation of the fetus, like systemic maternal disease, infections congenital abnormalities in the reproductive system, or psychosomatic alterations [27]. Clinical presentation of SA are haemorrhage in the deciduas secondary to implantation failure, causing expulsion of the products (14-22 wks GA).Retention within the cavity might also take place prior to 8-14wks of GA or rupture of membranes (ROM), with the expulsion of the fetus but not placenta, that persists in the uterine wall along with resulting in significant haemorrhage [9].
Human pregnancy needs haemostasis for avoiding haemorrhage, along with normal placentation, decidualization, as well as modulation of spiral arteries. The upregulation of MMP as well as cytokines are directly correlated with spontaneous preterm birth [28]. MMP, along with TIMP signalling In addition to their pleiotropic actions are needed for a normal pregnancy [29]. The MMP, along with TIMP expression is needed at the time of trophoblast invasion gets guided by trophoblast cells. MMP-9 that gets expressed by trophoblast cells present in the embryo, as well as TIMP-3 expressed by maternal uterine cells, are the major enzymes implicated in the invasion of trophoblast cells in the endometrium, being necessary for the implantation events along with proper generation of embryos [30]. MMP-2 generation starts at the initiation of gestation, with its major function being trophoblast bed invasion [31]. Upregulation of MMP-9, MMP-2, as well as TIMP-2 has a part in spontaneous pregnancy termination, mostly prior to 12 wks of GA [32]. Anumba, et al. observed that enhanced TIMP- 2 in women with history of recurrent pregnancy loss (RPL), correlated its aberrant implantation along with generation of the placenta, besides being an anticipatory marker for the same [33]. Expression of MMP mRNA along with MMP- 9: TIMP-3 mRNA ratio also have a correlation with SA [30]. Invasive capacity exhibited by EVT’s is given partly to regulation of extracellular matrix degradation mediated by matrix metalloproteinases (MMP’s) like MMP-2 and MMP- 9.Differential expression of these MMP’s and their tissue inhibitors (TIMPs) has been related to histone H3K9/27me3 [34]. Impairment of TIMP-1 expression correlates with infertility as well as early pregnancy loss [35]. Decidual stromal cells expressing CD82 Bupregulate the expression of TIMP-1 via an autocrine method along with inhibition of first trimester trophoblast cells invasiveness partially via β1 integrin /MAPK signalling pathway [36].
Preeclampsia (PE)
Preeclampsia (PE) as part of the hypertensive Disorder of pregnancy (HDP), is believed to be a serious complication worldwide. It has the properties of new onset hypertension in the 2nd part of pregnancy in correlation with proteinuria or thrombocytopenia, renal insufficiency, dysfunctional liver function, pulmonary oedema as well as cerebral or visual symptoms. These symptoms are secondary to poor placentation which causes vascular impairment along with main cardiovascular complications [37]. MMP’s get correlated with implantation of the embryo along with invasion of the trophoblast into the endometrium as well as inner myometrium. Thus impairment of MMP action is associated with aberrant placentation along with vascular impairment as observed in PE [38]. In the early few wks of pregnancy MMP’s start making a proper surroundings in the placental bed. The amount of MMP-2 and MMP-9 get sustained all through pregnancy with the major action being to act at the time of implantation In addition to conduct invasion of the trophoblast (MMP-9) along with the sustainance of of the placental bed (MMP-2) [39]. This invasion of the trophoblast cells gets guided by autocrine as well as paracrine stimuli from the uterine along with trophoblast cells. Additionally, various molecules having influence in PE generation like proteinases , cytokine as well as growth factors have a part in markedly shallow invasion of the trophoblast observed during pathophysiology of PE, resulting in impairment along with inefficient remodeling of the spiral arteries ,hence decreasing blood flow to the intervillous space resulting in systemic hypertension along with fetal hypoxia [40]. Plaks, et al. Evaluated a mouse model of PE had inactivation of maternal as well as embryonic MMP- 9, revealing a need for adequately balanced MMP-9 activation in normal placentation [41]. Reduction in MMP-2 and MMP- 9 expression /activation results in altered perfusion of the fetal-placental unit resulting in the liberation of factors within the maternal circulation along with the formation of endothelial dysfunction [42]. Enhanced MMP-2 activation/ amount in patients with PE have been documented in studies, since it can facilitate Platelet aggregation, vascular refashioning membrane In addition to the liberation of vasoactive peptides like endothelin1. These amounts have been demonstrated to get modulated via vascular endothelial growth factors (VEGF) [43]. There is an enhancement of MMP- 1 in patients with PE, although without enhanced TIMP-1. It is implicated in the breakdown of vascular collagen type1, that enhances vascular permeability promoting edema as well as proteinuria, that are properties seen in patients with PE. There is a stretch association among their neutrophils activation, along with vascular impairment observed in PE; like a part in leukocytes migration along with vascular impairment similarly its expression gets stimulated by activated neutrophils that also produce ROS along with TNF α [44]. The amounts of MMP in the urine specifically MMP- 2 have been hypothesized as being anticipating biomarkers for patients having great risk for PE among 12-16 wks GA. This demonstrates that an imbalance that is present in MMPs during pregnancy can influence the vasculature at both structural along with functional status with discernible alterations taking place along with clinical signs onset [45]. The regulated expression along with appropriate balance among MMPs, along with TIMP, that decides the net MMP activity, thus control the invasive capacity of placental trophoblasts are key for sustenance of normal placentation [46]. Generally, the reason that this disequilibrium can result in the generation of pathological disorders particularly, at the time of pregnancy, this balance disturbance influences the trophoblastic invasive cascade [31]. TIMP-1 represents an endogenous inhibitor of MMP-9 which might influence the responsively of HDP to treatment [47]. Levels of TIMP-1 are enhanced in PE [48]. Placentas Evaluated from pregnant women diagnosed with PE have demonstrated that at the transcriptional level, the aberrant expression of TIMP-1 can result in inadequate trophoblast invasion along with superficial placentation [49]. Nevertheless, rest of studies have demonstrated that decreased MMP-1 amounts in the umbilical serum placenta, In addition to decidua in pregnant women who had PE generation [50], Levels of TIMP-2 are further greater in women generating a hypertensive Disorder of pregnancy [51] Nevertheless, a particular part of TIMP-2 in PE generation needs to be clarified . TIMP-3 expression is significantly decreased in placentas of PE women, since TIMP-3 is an endogenous inhibitor of TACE, In addition to down regulation of TIMP-3 activity in trophoblasts cause an enhanced TACE expression as well as consequently TNF-α generation [52]. Histone deacetylases facilitate trophoblast cells migration along with trophoblast by repression of TIMP-promoter via histone hypoacetylation [53]. Moreover studies demonstrated that TIMP-3 promoter might be hypo methylated placentas of PE women, pointing that epigenetic changes might be linked with decreased trophoblast invasion [54]. Hypomethylated as well as placental particular TIMP-3 might be a potential marker for the early diagnosis of PE in matermal plasma. In case of PE escalated circulating TIMP-4 as compared to those in healthy pregnant women, along with these amounts are associated with clinical parameters of the disease. Rest of the studies have demonstrated that plasma TIMP-4 amounts are not changed prior to the presentation of clinical symptoms ,hence they are not good anticipators of PE generation [55]. It has been pointed that polymorphisms in TIMP-4 protect from the PE generation [56] along with TIMP-4 is a downstream target of the transcription factors glial cell missing (GCM)which might associate the sequences of decreased GCM 1 guided trophoblast differentiation to histological as well as functional components of imbalanced placentation in robust PE cases [57].
MMP along with TIMPs in Cancer
Cancer is a big cost for the caregivers in society, being a main etiology for mortality as well as morbidity althrough world. MMP along with TIMPs have a significant part in lot of stages of tumor generation. Disturbance in the balance among MMP along with TIMPs has been believed to be responsible for the propagation of Cancer [58].
To begin with, MMPs had been believed to take part in the initiation of metastases, in view of the disruption of the physical barriers which are the ECM In addition to the BM collagen. Nevertheless, at present insight is there that MMPs participate in all steps required for tumor propagation, implicating a lot of biological functions like the modulation of signalling pathways, the control of cytokines implicated in the immune response along with tumor growth (Specifically by developing angiogenesis and hence the cancer spread) [58]. MMP activity escalates in view of their part in refashioning of the ECM, the control of biological functions, along with the cytokines along with growth factors viability. Based on the MMP implicated in the tissue site where the tumor is present, they can further tumor repressive actions, like an e.g given by MMP-8 as well as MMP12, that have been appreciated as antitumor MMP’s [59]. MMP-11 influences a double action on tumors. MMP-11 facilitates cancer generation by inhibition of apoptosis along with escalation of invasion of cancer cells. Conversely, in case of animal models, MMP-11 has a negative part against cancer generation by repression of metastases [60]. It is significant to emphasize on the double action since MMP-11 might work as an important tumor biomarker in cancer for tumor staging, prognostic evaluation, monitoring of recurrence at the time of follow up, regards to immunotherapy, along with for early detection.
The event of pre metastases generation of a predetermined site is determined by refashioning of the ECM. MMPs are implicated in the liberation of soluble factors implicated in extra cellular recruitment In addition to expression of growth factor receptors in metastatic cells. These growth factors are VEGFR2, in Bone marrow (BM) obtained haematopoietic progenitor cells. MMP-9 has been seen to be overexpressed by endothelial cells as well as myeloid cells [61]. Tumor cell metastases is promoted by pre metastatic niches pre represented by BMDC along with Type IV collagen in the BM. This kind of collagen is liberated by a primary tumor in response to the hypoxic situations in the tumor surroundings, where BMDC’s breakdown Type IV collagen via the expression of MMP-2 that aids in invasion as well as metastases growth [62].
Although lots of MMP inhibitors got generated maximum were a flop ,pointing that still lots has to be deciphered as far as the correlation of MMPs with cancer is concerned. Despite the negative factors lot of experts in this arena are persisting with the trying to generate useful anti MMP drugs [63].
The TIMP family inhibits the proteolytic action of various MMPs implicated in the invasion of tumors as well as metastases, Nevertheless, TIMPs can also take part in tumor growth, apoptosis along with angiogenesis [64]. All TIMPs get liberated but only TIMP-3 gets incorporated into the matrix. The glycosylation of TIMP-3, escalates its affinity for glycan- bound MMPs. Abnormalities of glycosylation is generally common in cancer [65]. Besides TIMP-3 antagonizing Matrix Metalloproteinases activity along with repress tumor growth, angiogenesis, the invasion of tumors as well as metastases [66], TIMP-3 possesses other functions not associated with MMP inhibition, like the apoptosis stimulation [67] along with inhibition of endothelial cell motility as well as proliferation [68, 69]. As compared to TIMP-1 up regulation, there is lot of proof that TIMP-3 silencing occurs in a lot of human cancers [70].
Various authors demonstrated that silencing of the TIMP- 3 gene is correlated with a good prognosis in various kinds of cancer [71, 72, 73, 74]. Additionally, other than its MMP inhibition function, it is also clear that TIMP-1 facilitates cell growth, along with proliferation besides inhibiting apoptosis, along with is possibly implicated in the control of angiogenesis. enhanced amounts of TIMP-1 mRNA along with TIMP-1 protein are observed in various kinds of cancers, as well as various clinical studies have demonstrated a positive correlation of raised TIMP-1 protein with a poor prognosis or tumor propagation in lung, brain prostate, can have both pro as well as anti tumoral actions. as compared to the constant along with reverse patterns of TIMP-1 along with TIMP-3, separate trends have been documented for TIMP-2 along with TIMP-4 in ovarian cancer that might be secondary to heterogeneity of the kinds along with stages of cancer [75]. TIMP- reduces cell proliferation, neo angiogenesis, along with tumor growth. The overexpression signalling proliferation decrease of TIMP-2, both have been reported in relation to a lot of clinical studies of breast, lung, gastric as well as colorectal cancer. Similarly of TIMP-2, silencing by epigenetic means via promoter hyper methylation in prostate cancer [76]. Expression of TIMP-2 points towards a good prognosis in certain kinds of cancers [77, 78]. Conversely clinical studies demonstrate a correlation of greater TIMP-2 amounts with poor prognosis [79]. Akin to TIMP-2, observations have been there that TIMP-4 is up/down regulated in a variety of cancers. Studies in breast as well as prostate cancer have demonstrated an escalatation in TIMP-4 in the early stages of the disease at the end during the invasive stages. Correlations with other TIMP’s, lesser documentation of alterations in TIMP-4 have been seen [80, 81, 82].
Degranulation of TIMP results in a disturbed balance in key signals that start the pathways which result in the classical cancer steps of maintained signalling of proliferation,escape from growth repressors along with resistance to cell death, that aids in replicative immortalithy [75].
Currently, no particular changes known to be implicated in the propagation of cancer, like i)proliferation ii)survival iii) angiogenesis iv) aiding in replicative immortalithy v) invasion/migration along with vi)immunity escape [83], show that MMP along with TIMPs are implicated in the event of invasion along with metastases [84].
1) Control of Cell Growth MMP-1, MMP-2, MMP-3 along with MMP-7 can regulate the growth of tumor cells by modes like liberation of precursors of growth factors which are attached to the cell membrane, manipulating the bioavailability of growth factors along with controlling cell proliferation signals [85]. MMPs inhibit tumor growth via the activation of transforming growth factor beta (TGF β) as well as formation of propoptotic molecules [84]. TIMP’s are thought to have a lot of cellular functions, Specifically, the comparing actions of inhibiting tissue breakdown enzymes along with facilitating cellular growth [86]. TIMP-1 as well as TIMP-2 escalates the amount of Ras-GTP, Nevertheless, they utilize separate signalling pathways. TIMP-1 utilizes the tyrosine kinase/MAPK, signalling pathways, whereas TIMP-2 signalling utilizes protein kinase A, that has a direct role in the generation of Ras/phosphoinositide-3 – kinase (PI3-K) Complex [87]. Certain studies have pointed that the influence of TIMP-2 on cellular growth is brought about by its binding to MMP-2 on the Cell surface along with consequent activation of extra cellular-regulated-kinase (ERK) ½ [88]. Nevertheless, TIMP- 2 has separate functions as well as characteristics which are not dependent on its protease inhibitor activity. These characteristics are the activation of Shp-1 protein tyrosine phosphatase activity along with inhibition of tyrosine kinase receptor signalling by binding to α3β1integrin on the cell surface. This mode of receptor in activation causes inhibition of growth factor-modulated proliferation of both tumor (fibroblast along with endothelial cell) as well as normal cells [77]. TIMP-2 further inhibits growth by activating adenylate cyclise, escalating intracellular cyclic adenosine monophosphate (cAMP) amounts as well as activating the SH2 tyrosine phosphatase-1 (SHP-1) protein, that decreases the activity of growth factor receptors along with inhibits the activation of MAPK [89].
2) Control of Apoptosis MMP-7 along with ADAM10imparts antiapoptotic activity to cancer cells by cleavage of the Fas ligand from the Cell surface [90]. Further this antiapoptotic activity of MMPs gets regulated by proteolytic detachment of proteins correlated to the tumor linked major histocompatibility complex (MHC) Class 1. TIMP-1 inhibits apoptosis [91]. It binds to the CD63 receptor as well as crosstalks with the β1 subunit of integrins [92]. Subsequently the TIMP-1/CD63/β1integrin complex constitutively activates the survival signals via the activation of focal adhesion kinase (FAKs), PI3-K, along with ERK [93, 94]. TIMP-3 binds to ECM following liberation, that aids it to regulate peri cellular proteolysis via its robust inhibition of MMP, ADAM, along with ADAMTS [95]. Certain studies hypothesized that the antiapoptotic activity of TIMP- 3 occurs secondary to inhibition of TACE (ADAM17) with the subsequent stabilization of TNF receptors [96]. Other evaluation corroborated that TIMP-3 stabilizes 3 unique receptors [-TNF receptor1, FAS along with TNF related apoptosis inducing ligand receptor1 (TRAIL 1) [97].
3) Control of Angiogenesis Angiogenesis is necessary for tumor propagation. MMPs, mainly MMP-2, MMP-3, MMP-10, MMP-11, MMP-1, MMP- 8, along with MMP-13, possess 2 functions in this event, facilitating in beginning (but simultaneously, they possess a role in inhibition of angiogenesis [98, 99]. MMPs start angiogenesis via breakdown of type IV collagen in the BM, Specifically MMP-9, that liberates angiogenic factors like VEGF, fibroblast growth factors b(FGF-b), which are bound to the ECM [100]. On the other hand MMPs further inhibit the event of angiogenesis by facilitating the cleavage of type XVIII collagen, generating andotanine along with liberation of urokinase type plasminogen activator receptors which are bound to the cell surface that are needed for the invasion from endothelial cells to fibrin [101]. At present it has been determined that all the 4 TIMPs possess antiangiogenic action [102]. Possibly TIMP-1 is implicated in the control of angiogenesis, though this is still a controversial topic [103]. TIMP-2’s antiangiogenic action is brought about via crosstalk with α3β1integrin. As per studies over expression of TIMP-2 inhibited tumor growth along with angiogenesis by the up regulation of MAPK kinase phosphatase1, that dephosphorylates p38 MAPK, a molecule implicated in the proliferation along with migration of endothelial cells [104]. Additionally, TIMP-2 improves RECK expression that is a membrane–anchored protein possessing inhibitory action for mainly MMP-2, MMP-9, MMP-14, along with ADAM-10 [105], This antiangiogenic action TIMP-2 got localized to an area of a sequence of C terminal domain [106]. That might be the site of crosstalk with α3β1integrin. TIMP-3, similarly is a robust inhibitor of angiogenesis by binding directly to VEGFR2 along with blocks the VEGF action on endothelial cells [107]. Further TIMP-3, binds to angiotensin II type 2 receptor, along with overexpression of TIMP-3. In addition to angiotensin II type 2 receptor has additive inhibition of angiogenesis [108]. Little proof exists that crosstalk among endothelial cells along with pericytes, stimulates the expression of TIMP-3 in pericytes. TIMP-3 from this area in addition to TIMP-2 expresed in endothelial cells, takes part in vascular stabilization via inhibition of various MMPs In addition to ADAMs [109] along with probably by crosstalk with VEGFR2 [14].
4) Control of Invasion along with Metastases Progression of tumor takes place by a lot of processes in which cell motility along with migration combine with proteolysis, tumor cell proliferation at the newer area, along with angiogenesis, besides being correlated with crosstalk of cells with the ECM [110]. MMPs are implicated in cell invasion along with migration that is correlated with the later generation of colonies in earlier anticipated areas. Basically utilizing the aid of MMP-2, along with MMP-9, they succeed in breaking down of Type IV collagen of the BM, and hence get into the blood along with lymphatic vessels [111].
The overexpression of every TIMP within cancer lines inhibits the migration of these cancer lines via migration, invasion, metastases along with consequent growth [8]. Various MMP-s possess the capacity for cleaving cell adhesion molecules like cadherins, that bring about cell-cell contact along with integrins [112]. TIMP-1 represents an inhibitor of ADAM-10 along with TIMP-3 is the only inhibitor of ADAM- 17. The overexpression of these TIMPs inhibits NOTCH signalling [80]. Certain transcription factors like TWIST 1 along with SNAIL bring about a switch in the expression of cadherin (like E- cadherin loss or N- cadherin gain ) which promotes motility along with cell depolarization. SNAIL results in stimulation of TIMP-3 along with akin to that though independent of the metalloproteinases inhibitory function, the expression of TIMP-1 stimulates TWIST 1 to negatively control E- cadherin in cancer cells [113], whereas upregulation of this cadherin is induced via TIMP-2 .In case of fibroblasts ,the lack of TIMP-1 [114] as well as TIMP-3 [115] interferes with the action of cadherin along with cell-cell contact. Akin to that, the lack of TIMP-3 results in abnormal cellular distribution of E- cadherin along with associated β-catenin signalling [115]. Proof is there to demonstrate the liberation of TIMP-1, TIMP-3,or TIMP-4 inhibits the invasive event, whereas the liberation of TIMP-2 escalates this event [115]. Despite the TIMP-1-Metalloproteinase balance could regulate the matrix alignment, density along with rigidity, the proof corroborating this posit is minimal. Lysyl oxidase (LOXL) that crosslinks the collagen fibrils facilitates tumor cell motility [116]. Deletion of LOXL2 in cancer cells reduces the invasion along with TIMP-1 amounts. The generation of a metastatic niche is thought to be dependent on particular components of the ECM which are tenascin 2004:48:proteins [117], along with the overexpression of the isoforms of tenascin changes the expression of TIMP-3 along with MMPs [118].
5) Control along with Surveillance via Immune System MMPs possess a significant part in immunity escape along with activation again of TGF β, by which it indirectly controls the action of T lymphocytes. Implication of this is that in cancer tumor MMPs can indirectly influence the function of leukocytes, since chemokines are the direct MMP targets which avoid these molecules from working like chemo-attractants [119]. In the tumor evolution aberrations of cell proliferation along with death signals In addition to, inflammation which facilitates the tumor along with the pleiotropic cytokine TNF-β having the capacity to start these events .Akin to the physiological inhibitor of ADAM-17, TIMP-3, besides controlling TNF binding to the membranes against the systemic TNF, it also controls its2 receptors (TNFR1 as well as TNFR2) that are liberated into the blood. Lack of TIMP-3 results in escalatation of TNF signalling along with the inflammatory response secondary to the dysregulated action of ADAM-17 [120]. TIMP-3 is necessary for restricting inflammation. TIMP-3 deletion might escalate IL-6 generation through the TNF-αnuclear factor κB (NF κB) axis [121].
6) Control of Inhibitory Signals for Growth Trying to find the part of MMPs along with cancer propagation via proteolysis has aided in the generation of drug inhibitors of these enzymes [122]. To begin with, the transcription of MMPs gets inhibited, that persists in blocking the pathway which guides the transduction that initiates signals, like MAPK pathway along with ERK pathway, hence sending nuclear factors for inhibition of the expression of MMPs [396]. TIMPs can be controlled at the transcriptional level of different cytokines along with growth factors, causing tissue particular constitutive, or inducible expression. TIMP-1 expression that encodes genes can be controlled by TGF β. Rest of inhibitors (TIMP-2 along with TIMP-4) have lesser proneness to the actions of cytokines [123]. Overexpression of particular miR’s robustly escalating the start along with propagation of cancer via repression of TIMP proteins. miR-221 along with/or miR-222 can act on TIMP-3, stimulating drug resistance In addition to promoting cellular growth in various kinds of cancer via ADAM-10 along with ADAM-17 based signalling [75]. TIMP-3 repression is brought about by members of the family members of miR- 181. Intriguingly it has been demonstrated that TGF β, whose action is controlled by TIMPs, stimulate miR-181 via SMAD4 [75]. The miRs implicated in inflammatory diseases, like miR-21 stimulate the silencing of TIMP-3 [124]. miR- 196 causes repression of TIMP In addition to escalates the expression of MMP-1 In addition to MMP-9 indirectly via mitochondrial protein nucleoside diphosphate kinase 4 (NME4) along with N-terminal kinase of June[120]. The miRNA–TIMP axis has been posited as a treatment target for vicious or drug resistant human cancers. Hence new insight regarding the part of MMPs along with TIMPs in all stages of cancer propagation In addition to tumor surroundings could open new directed strategies regards to cancer therapy in the form of adjuvant treatments in early or advanced disease stages [125]. Although here our aim had been to discuss with regards to gynaecological malignancies, this is applicable for all body malignancies.
Utilization of MMP and TIMP Inhibition for Therapy
With the insight with regards to biology of MMP along with the control resulted in the generation of basic as well as preclinical experimental work in animal models besides clinical research in patients with any kind of pathologies involving gynaecological, other fields along with inflammatory problems but maximum was in cancer. These studies have mainly laid concentration on MMPs, in view of thoughts that their aetiology is related to changes in significant physiological modes which result in aberrant angiogenesis, apoptosis along with immune modulatory action, thus aiding in growth of tumor as well as/or metastases [126]. Pharmacological industry gave monetary support for research with regards to the generation of broad spectrum MMP inhibitors, nevertheless, the exact physiology in addition to biology of MMP’s, besides how they have evoluted with regards to time along with control by separate molecules are not clear. Clinical trials did not get the anticipated outcomes, deteriorating the prognosis of disease, in view of delivery in advanced stages resulting in side effects like musculoskeletal toxicity [126]. Currently it is known that MMP’s participate in physiological events like embryogenesis, angiogenesis, tissue remodeling, bone generation, wound healing, Mammary involution, promotion of liberation of signalling molecules, cell migration, activation of signalling molecules along with immunity [126]. The following are the MMP inhibitors In addition to the treatments which work against MMP’s are detailed here.
Hydroxamate Dependent Inhibitors
The initial molecules that got generated were hydroxamate dependent inhibitors. Their bases is on the structure of collagen along with mainly implicate compounds that possess a backbone that have been fashioned to simulate the normal peptide substrate of the wanted MMP’s along with a group which chelates the catalytic Zn2+ ion. They decrease the aid of the remaining of the compounds to the inhibitor- enzyme binding event, thus aiding widespread inhibition. Certain examples of this kind of drug are marimastat, ilomastat as well as batimastat. Of these batimastat was the first MMP inhibitors to enter clinical trials, In addition to that it was demonstrated to inhibit various MMP’s, that are MMP- 1, MMP-2, MMP-7, along with MMP-9 showing antitumor actions in animal models of human ovarian cancer, colorectal cancer ,melanoma along with haemangioma. Marimastat was observed to be inefficacious besides resulting in musculoskeletal side effects in a randomized phase III trial for metastatic breast cancer cases which were stable or responding following first line chemotherapy. The reason for the musculoskeletal pain is believed to be secondary to the inhibition of ADAM as well as ADAMTS family members, that are ADAM17.Usually MMP inhibitors have been found to be delivered very late for making any change, since basic work was conducted in early stages of the disease as well as Clinical work was meant to be a last resort in the therapy of terminally ill patients [126].
Newer Hydroxamate Dependent MMP Inhibitors Formation
These ones have been formed to have greater specificity, with a view of reducing the side effects. MMI270, MMI-166, PD-166793, ABT-770, cipemastat, along with prinomastat. MMI-166 represents a selective inhibitor of MMP-2, MMP- 9, along with MMP-14. PD-166793, prinomastat as well as ABT-770 were generated to prevent binding to the’’shallow pocket’’ of MMP-1, dependent on the thought (at that particular time) that MMP-1 sparing inhibitors might not result in musculoskeletal side effects. Cipemastat, that inhibits MMP-1, TIMP-3, along with MMP-9, was utilized for the therapy of rheumatoid arthritis (RA), as well as osteoarthrithitis, although it could not avoid the propagation of joint damage.
Non - Hydroxamate Dependent MMP Inhibitors
Hydroxamic acid are usually metabolically labile but there exist a lot of other zinc –binding groups which are stable .Reverse Hydroxamates as well as non hydroxamate inhibitors, like carboxylates, hydroxy carboxylates, sulfhydryls. Phosphoric acid derivatives as well as hydantoins got generated to prevent the drawbacks linked to with the 1st generation MMP inhibitors, like metabolic inactivation, besides the chelation of metals of rest of metalloproteins [126]. MMP structure has been illustrated with the aid of crystallography along with novel inhibitors have been fashioned with different peptidomimetic as well as non peptidomimetic backbone, structures. Rebamastat, which is a thiol zinc–binding groups, represents a wide spectrum MMP inhibitor; nevertheless, a phase II trial for early stage Breast Cancer, besides a phase III trial in nonsmall cell lung carcinoma both documented side effects [126]. Tanomastat was shown to have problems with the dosing along with timing of delivery linked to disease propagation , besides the effectiveness of this agent was demonstrated to be variable ,with controversial outcomes being derived based on the timing of delivery [79]. Ro28-2653 inhibits MT –MMP, MT3- MMP, MMP2, MMP8 as well as MMP9 yet spares MMP-1 along with ADAM17 activity. Various promising experimental work in animal models has shown its antitumor along with anti angiogenic action but they did not move to Clinical trials [126]. Robust inhibitors of MMP-13 as well as MMP-12 got generated utilizing the alternate zinc –binding group hydantoin. Various biphenyl sulphonamide carboxylate MMP inhibitors got fashioned for therapy of osteoarthrithitis via inhibition of MMP-13.Tetracycline antibiotics, like doxycycline along with minocycline, possess innate MMP inhibitory ability. Doxycycline has been indicated for the therapy of periodontal disease as well as is the only collagenase inhibitor that has got approval from US Food and Drug Administration (FDA) for any kind of human disease [126].
Alternate Binding Sites Getting Targeted
For decreasing the off target actions seen in clinical trials, besides preventing wide MMP inhibition in view of a big structural homology of the various MMPs. Current work has shifted from targeting the catalytic area to alternate ones that have been lesser conserved sites. Besides the Zn2+ ion in their catalytic area, MMPs have subsitess (S) labeled as unprimed or primed. The P’-S1’ crosstalk is the major factor that determines the affinity of inhibitors along with cleavage positions of peptide substrates. Like extension of the P1 substituent was utilized to obtain selectivity of MMP- 13 above the markedly homologous MMP-2. The utilization of NMR along with Xray crystallography techniques in addition to, computational strategies that aid in modeling of drug-protein Crosstalk with Non-Hydroxamate dependent MMP inhibitors which bind to sites other than catalytic. Lot of these inhibitors showed good MMP-13 selectivity which lead to decreased clinical symptoms when evaluated in a mouse model of RA as well as osteoarthritis. The combined techniques with computational anticipation showed hidden areas in the MMP structure, that cannot get utilized to form a rational fashioning of novel molecular effectors as well as therapeutic targets [126].
Treatment which is Antibodies Based
These molecules possess great selectivity besides having lot of functional blocking antibodies. Selectively they target membrane anchored MMPs. The markedly selective antibody–dependent MMP-14 inhibitor DX-2400 has demonstrated antitumor anti angiogenic, along with anti invasive characteristics, besides blocking MMP-14- based pro- MMP-2 processing. MMP-14 inhibitory antibodies have got successfully evaluated in vivo and in vitro. Depending on the 3-D structure as well as amino acid sequence of MMP- 13, that is a neutralizing antibodiy which binds to the active form of MMP-13, without binding the latent form or to other MMPs was generated [126].
Endogenous Inhibitors of MMP Function-Α
Polymyxin-conjugated α-2 macroglobulin is a big serum protein which controls MMP function. MMP’s are embedded within the macroglobulin, hence avoiding the MMP’s from coming across large substrates. Polymyxin-conjugated α-2 macroglobulin showed protective action in mouse models of sepsis, with this action being correlated with binding to along with neutralizing inflammatory cytokines; MMP inhibition has not got evaluated [126].
Theoretically TIMPs represent the basis for another innovative class of MMP inhibitor, nevertheless, they have hardly been thought of. They have been utilized in model system’s to give key clues regarding the effectiveness of MMP inhibitors in various diseases. Nevertheless, TIMPs besides inhibiting MMPs, in particular cases they can indirectly facilitate MMP action. Like, domain- particular overexpression of TIMP-2 as well as TIMP-3 illustrated the in dependent functions of TIMPs at the time of generation [126].
Despite TIMPs have been believed in present treatment approaches for the control of MMPs,in particular in various diseases, no success. Propagation has got attained in this field , that is probably in view of the fact that certain diseases share akin pathways or modes of action where MMPs as well as TIMP-3 intervene besides not playing common parts. Usual events of the MMPs various diseases are inflammation, cell death, along with migration, of rest. Getting insights regarding subtleties of MMP biology is not enough, hence it is essential to think of crucial points to enhance understanding in this particular area of study.
Discussion
Despite detailing the etiopathogenesis by which MMPs and TIMPs modulate various obstetric and gynaecological diseases like PCOD, IUGR, preeclampsia, spontaneous abortion and newer roles in DUB, Endometriosis in addition to various aspects of cancer manipulation the limitations of this study is that although a lot of MMP inhibitors have been developed yet no satisfactory agent has been generated thus far, with their complications being the limiting factor. Nevertheless, getting this insight is significant in the sense that with newer advances and knowledge with regards to the extracellular Vesicles (ECV’s) one can generate innovative therapies of supplying a particular MiRNA, other substances and as per desire modulate as per the etiopathogenesis of the diseases via the ECVs and hence might aid in cancer regulation be sides placentation abnormality etc.
Conclusion
Till date, no precise treatment dependent on MMP inhibition has become available for the Clinical utilization in chronic diseases, hence newer approaches are required for proper utilization of the advantages by the understood modes of MMPs as well as TIMP-3.The inhibition of MMP liberation as well as/or function in the right place with the aim of giving a better quality of life (Q.O.L) to those patients with an illness linked to the impairment of control of MMPs implicated in maximum diseases might be beneficial. It is essential to keep conducting research for evaluation of the part played by in maximum diseases like in MMP-2 along with MMP-9. The functions of MMPs are tough to anticipate as already known, Nevertheless, by having insight regards to mode by which they work for deterioration of diseases, can be advantageous, besides working in form of treatment, as bio-markers, along with risk anticipators of diseases. It is clarified here how much heterogeneity in the status of TIMPs based on the various diseases, since they vary in the kinds of crosstalk that they go through various MMPs along with independently of MMPs. Nevertheless, a generalized escalation of in TIMP-1 along with generalized silencing of TIMP-3can be seen as the the disease advances along with deterioration of prognosis occurs. Extra studies are essential for the particular evaluation of these endogenous inhibitor in various chronic diseases. Alternate strategy to the treatment of MMP-associated diseases could get utilized for engineering of TIMPs possessing limited inhibitory specificities. Further widening of our horizon on the particular part of MMPs as well as TIMP-3 via studies along with evaluation of the MMPs actions at the post translational level, in view of maximum studies in the literature have only analyzed transcriptional action, with little insight on intracellular actions of MMPs to know the relevant functions of pathological significance which could yield greater knowledge on the various parts played by MMPs as well as TIMP-3 in the various diseases. Moreover it is essential to carry out greater in vivo work to illustrate the functions of MMPs as well as TIMP-3, since it is not currently possible to illustrate how these are controlled/ impairment of control, besides finding which substrates interfere in various events. More Evaluation is needed for finding the actions of MMPs under pathological along with non pathological disorders.
Further recently Timokhina et al conducted a retrospective study that included 92 pregnant women at a gestational age of 26-38 weeks, of which the principal group consisted of 61 patients with severe PE. They divided the principal group into two subgroups: the first subgroup was named the severe early-onset preeclampsia (EO-PE) group and consisted of 30 pregnant women. The second group was designated the severe late-onset preeclampsia (LO-PE) group, comprising 31 patients. They determined the plasma concentrations of MMPs 2 and 9 in the groups with an ELISA. They observed that in the group of PE patients with both EO- PE and LO-PE, the amount of MMP-2 was significantly higher as compared to the women undergoing normal pregnancy; besides that they did not find any significant variation on comparison of the amounts of MMP-2 in the subgroups with EO-PE as well as LO-PE. Evaluation of the amounts of MMP- 9 in EO-PE and LO-PE subgroups illustrated ameliorated amounts of MMP-9 in both groups relative to the control group. Further they observed a decreased amount of MMP-9 in the EO-PE group as compared to the LO-PE group. Thus concluding that the significantly escalated amounts of MMP- 2 in women-both in the EO-PE and LO severe PE subgroups- reason out the participation of this enzyme in endothelial impairment in the second stage of severe PE. A decrease in MMP-9 in the EO-PE group validated the participation of MMP-9 in the event of spiral artery transformation [127].
Earlier we have reviewed role of prokineticins in reproduction [128] Su MT, had documented the role of miR- 346 and miR-582-3p in controlling EG-VEGF expression and trophoblast invasion via matrix metalloproteinases 2 and 9 [129]. Further earlier we had reviewed the role of epigenetics in Preeclampsia,and recently the role of Extracellular Vesicles
(ECV’s),in human reproduction and how we can utilize their cargo like miRNA’s in treating various metabolic diseases like DM, and similarly might utilize them for targeting miR’s along with MMP’s in Preeclampsia [130, 131, 132].
Ding et al. revealed that miR-519-d-3p suppresses invasion and migration of trophoblast cells via targeting MMP2 [133].
Further members of the placenta-specific miRNA cluster C19MC, including miR-519d, are secreted by fetal trophoblast cells within extracellular vesicles (EVs). Earlier we have reviewed the role of ECV’s in human reproduction besides their roles in carrying different microRNA’s besides proteins, lipids and nucleic acids in their cargo [131, 132]. Recently Chaiwangyen et al. demonstrated that trophoblast-derived EVs can be internalized by the autologous trophoblast and surrounding maternal immune cells, resulting in coordination of cellular responses. The study of functions and targets of placental miRNAs in the donor and recipient cells might aid in getting insight of the immune tolerance that is necessary in pregnancy. Here, they documented that miR-519d-3p levels were associated positively with cell proliferation and negatively with migration in trophoblastic cell lines. Inhibition of miR-519d-3p in JEG-3 cells increases caspase-3 activation and apoptosis. PDCD4 and PTEN are targeted by miR-519d-3p in a cell type-specific manner. Transfection of trophoblastic cell lines with miR-519d mimic results in secretion of EVs containing elevated levels of this miRNA (EVmiR-519d). Autologous cells enhance their proliferation and decrease their migration ability when treated with EVmiR-519d. NK92 cells incorporate EV-delivered miR- 519d-3p at higher levels than Jurkat T cells. EVmiR-519d increases the proliferation of Jurkat T cells but decreases that of NK92 cells. Altogether, miR-519d-3p regulates pivotal trophoblast cell functions, can be transferred horizontally via EVs to maternal immune cells and exerts functions therein. Vesicular miRNA transfer from fetal trophoblasts to maternal immune cells may contribute to the immune tolerance in pregnancy [134]. Besides other authors have shown how ECV’s might be used in other disorders [135].
Moreover Selective intrauterine growth restriction (sIUGR) is a disorder of monochorionic (MC) twin pregnancies. However, the underlying mechanism remains largely unknown. Trophoblast cells are the major component of the placenta. Dysfunction of trophoblast cells is associated with placental dysfunction. In their earlier study, Wenetal [136] identified miR-338-5p is down regulated in placenta tissues sharing larger twins of sIUGR. In the present study, they aimed to investigate the role of miR-338-5p in trophoblast cells and explored its target. Their results further indicated that miR-338-5p was down regulated in placental tissues supporting larger twins of sIUGR, whereas epidermal growth factor-containing fibulin-like extracellular matrix protein 1 (EFEMP1) was upregulated. Moreover, miR- 338-5p overexpression suppressed the growth and invasion of trophoblast cells. Importantly, results from luciferase reporter assay demonstrated that miR-338-5p bound on the 3′-UTR of EFEMP1. miR-338-5p suppressed the growth and invasion of trophoblast cells via targeting EFEMP1. Further, miR-338-5p/EFEMP1 might disrupt the function of trophoblast cells via inhibiting the phosphorylation of AKT [136].
Earlier Li et al. had evaluated the differential expression of same miR-338-5p of the sa miR-338-5p in gliomas and the role of miR-338-5p in glioma cell invasion via its potential target gene TSHZ3 encoding Teashirt zinc finger homobox 3, predicted by bioinformatics, and matrix metallopeptidase 2 (MMP2), the key pro-invasive protease overexpressed in gliomas. Quantitative real-time reverse transcription PCR (qRT-PCR) and Spearman correlation analysis were used to determine differential expressions of miR-338-5p and TSHZ3 in astrocytic gliomas of different grades (n = 35) and glioblastoma cell lines (U87 and U251) in comparison to non-neoplastic brain (NNB) tissues (n = 6). Western blotting was used to determine the protein levels of TSHZ3 and MMP2 in glioblastoma cell lines and Matrigel invasion assay to examine the role of miR-338-5p in cell invasiveness. The results demonstrated that the expression of miR-338-5p, normalized to hsnRNA U6, was significantly higher in grade III and IV gliomas and glioblastoma cell lines in contrast to that in NNB and grade II gliomas, whereas TSHZ3 expression, normalized to GAPDH, was inversely related to miR-338-5p (R = -0.636, P < 0.01). Luciferase assays showed TSHZ3 to be a target gene of miR-338-5p. In both U87 and U251 cells, miR-338-5p mimics increased MMP2 and invasiveness of the cells. Overexpression of ectopic TSHZ3 suppressed the cell invasiveness and attenuated the pro-invasive effect of miR- 338-5p mimics. Overall, their results demonstrated that miR- 338-5p has a function in promoting glioma cell invasion by targeting TSHZ3 suppression on MMP2. In conclusion, miR- 338-5p is a possible potential biomarker for the diagnosis and target for therapy of high-grade glioma [137].
This intricate insight has resulted in not only treating tumours, utilizing ECV’s , but intricate knowledge of molecular modes like MMPs with micro RNA interaction might aid in developing therapies by delivering ECV’s from particular microRNA implicated in the pathogenesis of diseases like preeclampsia, IUGR, besides lot of malignancies [138].
Moreover how curcumin might be participating in endometriosis treatment was by targeting MMP2 as well as 9, besides further melatonin might target by attacking MMP3 along with apoptotic pathways [139, 140, 141, 142].
Further Grzechocinska et al. demonstrated how there was an absence of association among lack of correlation between proMMP-2 along with MMP-2 amounts point that separate pathway of their activation in AUB. ProMMP-2 is up regulated by estradiol and down regulated by progesterone while MMP-2 levels increase with the length of bleeding [143]. This way with knowledge given by Johanns et al with regards to Cellular uptake of proMMP-2:TIMP-2 complexes by the endocytic receptor megalin/LRP-1 ,with earlier knowledge on how low-density lipoprotein receptor-related protein-1 (LRP-1) mediates the clearance of a complex between the zymogen form of MMP-2 (proMMP-2) and tissue inhibitor of metalloproteinases, TIMP-2, in HT1080 human fibrosarcoma cells . Here they demonstrated that, in BN16 rat yolk sac cells, proMMP-2: TIMP-2 complex is endocytosed through a distinct LRP member, megalin/LRP-2. Addition of receptor-associated protein (RAP), a natural LRP antagonist, caused accumulation of endogenous proMMP-2 and TIMP- 2 in conditioned They e concluded that megalin/LRP-2 can efficiently mediate cell-surface binding and endocytosis of proMMP-2: TIMP-2 complex. Therefore megalin/LRP- 2 can be considered as a new actor in regulation of MMP-2 activity, an enzyme crucially involved in many pathological processes. Thus more and more insight is being developed regarding targeting DUB with various MMP pathways [144]. Further Yohan et al showed how targeting the MMP-14/ MMP-2/integrin αvβ3 axis with multispecific N-TIMP2– based antagonists might be utilized for cancer therapy thus utilizing the newer MMP-14or membrane type 1(MT-1- MMP) axis [145].
References
-
Cui N, Hu M, Khalil RA (2017) Biochemical and Biologi- cal Attributes to Matrix Metalloproteinase. Prog Mol Biol Transl Sci 147: 1-73.
-
Griselda ACP, Idalia GV, Claudia CDR, Ramirez AJM, Guerrero RJF, et al. (2020) The roles of Matrix metalloproteinases and their inhibitors in Human Disease. Int J Mol Sci 21(24): 9739.
-
Roth GA, Johnson C, Abajobir A, Abd Allah F, Abera SF, et al. (2017) Global, regional and national burden of cardiovascular diseases for 10 causes 1990-2015. J Am Coll Cardiol 70(1): 1-25.
-
Cork BA, Li TC, Warren MA, Laird SM (2001) Interleukin- 11(IL-11) in human endometrium: expression throughout the menstrual cycle and the effects of cytokines on endometrial IL-11production in vitro. J Reprod Immunol 50(1): 3-17.
-
Godbole G, Suman P, Gupta SK, Modi D (2011) Decidualized endometrial stromal cells derived factors promote trophoblast invasion . Fertil Steril 95(4): 1278- 1283.
-
Licht P, Fluhr H, Newwinger J, Wallwiener D, Wildt L (2007) Is human chorionic gonadotropin directly involved in the regulation of human implantation? Mol Cell Endocrinol 269(1-2): 85-92.
-
Karthikeyan VJ, Lane DA, Beevers DG, Lip GY, Blann AD (2013) Matrix Metalloproteinases and their Tissue inhibitors in hypertension –related pregnancycomplication. J Hum Hypertens 27: 72-78.
-
Fauser BC, Tartalazis BC, Rebar RW, Legro RS, Balen AH, et al. (2012) Consensus on women’s health aspects of Polycystic Ovarian Syndrome (PCOS): The Amsterdam ESHRE/ASRMSponsored 3rd PCOS Consensus Workshop Group. Fertil Steril 97(1): 28-38.
-
Abraham Suzanne OJ (2015) Fundamentals of Obstetrics and Gynaecology. 10th (Edn.), Elsevier: Amsterdam. The Netherlands, pp: 384.
-
McCartney CR, Marshall JC (2016) Polycystic Ovarian Syndrome. N Engl J Med 375: 54-64.
-
Orio F, Muscogiuri G, Nese C, Palomba S, Savastano S, et al. (2016) Obesity, type 2 diabetes mellitusand cardiovascular disease risk:an update in the management of Polycystic Ovarian Syndrome. Eur J Obstet Gynaecol Reprod Biol 207: 214-219.
-
Hulboy DL, Rudolph LA, Matrisian LM (1997) Matrix Metalloproteinases as mediators of reproductive function. Mol Hum Reprod 3(1): 27-45.
-
Dambala K, Vavilis D, Bili E, Goulis DG, Tartalazis BC (2017) Serum visfatin , vascular endothelial growth factor and Matrix Metalloproteinase-9 in women with Polycystic Ovarian Syndrome. Gynaecol Endocrinol 33(7): 529-533.
-
Shalev E, Goldman S, Ben-Shlomo I (2001) The balance between MMP-9 and MMP-2 and their Tissue inhibitors (TIMP-1)in luteinzed granulose cells: comparison between women with PCOS and normal ovulatory women. Mol Hum Reprod 7(4): 325-331.
-
Liu B, Cai LY, Lv HM, Xia L, Zhang YJ, et al. (2008) Raised serum levels of Matrix Metalloproteinase-9 in women with Polycystic Ovarian Syndrome and its association with insulin like growth factors binding protein. Gynaecol Endocrinol 24(5): 285-288.
-
Gomes VA, Viera CS, Jacob Ferreira AL, Belo VA, Soares GM, et al. (2011) Imbalanced circulating Matrix Metalloproteinase in Polycystic Ovarian Syndrome. Mol Cell Biochem 353(1-2): 251-257.
-
Ackali A, Bostanci N, Ozcaka O, Ozturk Ceyhan B, Gumus P, et al. (2015) Elevated Matrix Metalloproteinase-8 in saliva and serum in Polycystic Ovarian Syndrome and its association with gingival inflammation. Innate Immune 21(6): 619-625.
-
Lewandowski KC, Komorowski J, O Callaghan CJ, Tan BK, Chen J Prelevic GM, et al. (2006) Increased circulating levels of Matrix Metalloproteinase2 and -9 in women with Polycystic Ovarian Syndrome. J Clin Endocrinol Metab 91: 1173-177.
-
Zhou F, Shi LB, Zhang SY (2017) Ovarian fibrosis: a phenomenon of concern. Clin Med J 130(3): 365-371.
-
Ben Shlomo I, Goldman S, Shalev E (2003) Regulation of Matrix Metalloproteinase -9(MMP-9), Tissue inhibitor of MMP, and progesterone secretion in luteinzed granulosa cells from normally ovulating women with Polycystic Ovarian Syndrome. Fertil Steril 79(1): 694-701.
-
Oksjoki S, Rahkonen O, Haarala M, Vuorio E, Antilla l (2004) Differences in connective tissue gene expression between normally functionining Polycystic and post- menopausal ovaries. Mol Hum Reprod 10(1): 7-14.
-
Baka S, Zouria K, Kouskoni E, Makrakis E, Demedriou S, et al. (2010) Matrix Metalloproteinase 2 and -9(MMP-9), and their Tissue inhibitors in follicular fluid in patients with Polycystic Ovarian Syndrome undergoing in vitro fertilization. In Vivo 24: 293-96.
-
Barisic A, Devic Pavlic S, Ostojic S, Pereza N (2018) Matrix Metalloproteinases and, Tissue inhibitors of Metalloproteinases gene polymorphisms in disorders that influence fertility and pregnancy complications. A Systematic review and meta analysis. Gene 647: 48-60.
-
Rouse CE, Eckert LO, Barbarinsa L, Fay E, Gupta M, et al. (2017) Spontaneous Abortion and ectopic pregnancy:Case definition and guidelines for the datacollection, analysis, and presentation of maternal immunization safety data . Vaccine 35(48): 6563-6574.
-
Meaney S, Corcoran P, Spillane N, O Donoghue K. (2017) Experience of miscarriage: an interpretive phenomenological analysis. BMJ Open 7(3): e011382.
-
Franasiak JM, Scott RT (2017) Contribution of immunology to implantation failure of euploid embryos. Fertil Steril 107(6): 1279-1283.
-
Pereza N, Ostojic S, Kapovic M, Peterlin B (2017) Systematic review and meta-analysis of genetic association studies in idiopathic recurrent Spontaneous Abortion. Fertil Steril 107(1): 150-159.
-
Schatz F, Guzeloglu KO, Arlier S, Kayisli UA, Lockwood CJ (2016) The role of Decidual cells in uterine hemostasis, menstruation, inflammation, adverse pregnancy outcomes and abnormal uterine bleeding. Hum Reprod Update 22(4): 497-515.
-
Moore CS, Crocker SJ (2012) An alternate perspective on the roles of TIMPs and MMP’s in pathology. Am J Pathol 180(1): 12-16.
-
Jiang G, Qi Y (2015) Detection of MMP-9 and TIMP-3 mRNA expression in the villi of the patients undergoing early Spontaneous Abortion: a report of 30 cases. Exp Med 9(5): 1939-1943.
-
Onogi A, Naruse K, Sado T, Tsunemi T, Shigomi H, et al. (2011) Hypoxia inhibits invasion of extravillous trophoblast cells through reduction of Matrix Metalloproteinase(MMP)-2 activation in the early first trimester of human pregnancy. Placenta 32(9): 665-670.
-
Nissi R, Talvensaari MA, Kotila V, Ninimaki M, Javela I, et al. (2013) Circulating Matrix Metalloproteinase MMP-9 and MMP-2/TIMP-2 complex are associated with Spontaneous early pregnancy failure. Reprod Biol Endocrinol 11: 2.
-
Anumba DO, El Gelany S, Elliott SL, Li TC (2010) Circulating levels of Matrix proteinases and their inhibitorsin pregnant women with and without history of recurrent pregnancy loss. Reprod Biol Endocrinol 8: 62.
-
Rahat B, Sharma R, Bagga R, Hamid A, Kaur J (2016) Imbalance between matrix metalloproteinases and their tissue inhibitors in preeclampsia and gestational trophpblastic diseases. Reproduction 152(1): 11-22.
-
Skrzypczak J, Wirstein P, Mikolajczyk M, Ludwikowski G, Zak T (2007) TGF superfamily and MMP, MMP-9, TIMP-1 Genes expression in the endometrium of women with impaired Reproduction. Folia Histochem Cytobiol 45(Suppl1): S143-S148.
-
Li MQ, Hou XF, Shao J, Tang CL, Li DJ (2010) The DSC- expressed CD82 controls the invasiveness of trophpblast cells via integrinβ1/MAPK/ MAPK3/1 signalling pathway in human first trimester of pregnancy. Biol Reprod 82(5): 968-979.
-
ACOG Committee on Obstetrics Practice (2002) ACOG Practice Bulletin. Diagnosis and management of Preeclampsia and eclampsia. Number 33, January 2002. American College of and Obstetricians and Gynecologists. Int J Gynaecol Obstet 77(1): 67- 75.
-
Klein T, Bischoff R (2011) Physiology and patho Physiology of matrix metalloproteinases. Amino Acid 41(2): 271-290.
-
Staun Ram E, Goldman S, Gabarin D, Shalev E (2004) Expression and importance of matrix metalloproteinases-2 and 9 (MMP-2 and MMP-9) in human trophpblast invasion. Reprod Biol Endocrinol 2: 59.
-
Zhu JY, Pang ZH, Yu YH (2012) Regulation of trophoblast invasion: the role of matrix metalloproteinases. Rev Obs Gynaecol 15(3-4): e137-e143.
-
Plaks V, Rinkenberger J, Dai J, Flannery M, Sund M, et al. (2013) Matrix metalloproteinases-9 deficiency phonocopies features of Preeclampsia and intrauterine growth restriction. Proc Natl Acad Sci USA 110(27): 11109-11114.
-
Chen J, Khalil R (2017) Matrix metalloproteinases in normal Pregnancy and Preeclampsia. Prog MolBiol Transl Sci 148: 87-165.
-
Myers JE, Merchant SJ, Macleod M, Mires GJ, Baker PN, et al. (2005) MMP2 levels are elevated in the plasmaof women who subsequently develop Preeclampsia. Hypertens Pregnancy 2: 103-115.
-
Estrada GG, Cappello RE, Mishra N, Romero R, Strauss JF 3rd, et al. (2011) Increased expression of Matrix metalloproteinases-1 in systemic vessels of Preeclamptic women: a critical mediator of vascular dysfunction. Am J Pathol 178(1): 451-460.
-
Martinez FML, Perez FA, Garza VI, Espinosa JMA, Avila CL, et al. (2018) Matrix metalloproteinase multiplex screening identifies increased MMP2 urine concentrations in women predicted to develop Preeclampsia. Biomarkers 23(1): 18-24.
-
Staun RE, Shalev E (2005) Human trophoblast function during the implantation process. Reprod Biol Endocrinol 3: 56.
-
Luizon MR, Palei AC, Sandrim VC, Amaral LM, Machado JS, et al. (2014) Tissue inhibitors of Metalloproteinases-1 polymorphism, plasma TIMP1 levels, and anti- Hypertensive therapy responsiveness in Hypertensive Disorders of pregnancy. Pharm J 14(6): 535-541.
-
Montagnana M, Lippi G, Albiero A, Scevarolli GL, Franchi M, et al. (2009) Evaluation of Matrix metalloproteinases-2 and 9and their inhibitors in physiologic and Pre- eclamptic pregnancy. J Clin Lab Anal 23(2): 88-92.
-
Zhang H, Lin QD, Qiao C (2006) Expression of trophoblast invasion related genes mRNA and protein in placenta in Preeclampsia. Zhonghua Fu Chan Ke Za Zhi 41: 509-513.
-
Deng CL, Ling ST, Liu XQ, Zhao YJ, Lv YF (2015) De- creased Expression of Matrix metalloproteinases-1 in the maternal umbilical serum, trophoblasts and deciduas leads to Preeclampsia. Exp Med 9(3): 992-998.
-
Lavee M, Goldman S, Daniel Spiegel E, Shalev E (2009) Matrix metalloproteinases-2 is elevated inmid trimester amniotic fluid prior to the development of Preeclampsia. Reprod Biol Endocrinol 7: 85.
-
Ma R, Gu B, Gu Y, Groome LJ, Wang Y (2014) Downregulation of TIMP-3 leads to increase in TACE expression and TNF-αproduction by placental trophoblast cells. Am J Reprod Immunol 71(5): 427-433.
-
Xie D, Zhu J, Liu Q, Li J, Song M, et al. (2019) Dysregulation of HDAC9 represses trophoblastcells migration and invasion through TIMP-3 activation in Preeclampsia. Am J Hypertens 32(5): 515-523.
-
Xiang Y, Zhang X, Li Q, Xu J, Zhou Y, et al. (2013) Promoter Hypomethylation of TIMP-3 is associated with Preeclampsia in a Chinese population. Mol Hum Reprod 19: 153-159.
-
Sandrim VC, Diniz S, Eleuterio NM, Gomes KB, Dusse LMS, et al. (2018) Higher levels of circulating TIMP- 4 in Preeclampsia is strongly associated with clinical parameters and microRNA. Clin Exp Hypertens 40(7): 609-612.
-
Sandrim VC, Luizon MR, Machado JS, Cavalli RC (2019) C>T (RS17035945) polymorphisms in TIMP-4 protects against Preeclampsia.J Obs Gynaecol 39(1): 135-137.
-
Drewio S, Czikk M, Baczyk D, Lye S, King- dom J (2011) Glial cell missing 1 mediates overexpres- sion of Tissue inhibitors of Metalloproteinases-4 in severe Pre-eclamptic placental villi. Hum Reprod 126(5):1025- 1034.
-
Baker AH, Edwards DH, Murphy G (2002) Matrix metalloproteinase inhibitors :biological actions and therapeutic opportunities. J Cell Sci 115(Pt-19): 3719- 3727.
-
Dufour A, Overall CM (2013) Missing the target: Matrix metalloproteinase anti targets in inflammation and cancer. Trends Pharm Sci 34(4): 233-242.
-
Lopez OC, Matrisian LM (2007) Emerging roles of proteases in tumor suppression. Nat Rev Cancer 7(10): 800-808.
-
Zhang X, Huang S, Guo J, Zhou L, You L, et al. (2016) Insights into the distinct roles of MMP-11 in tumor biology and future therapeutics (Review). Int J Oncol 48(5): 1783-1793.
-
Kaplan RN, Riba RD, Zacharoulis S, Bramley AH, Vincent L, et al. (2005) VEGFR1 Positive haematopoietic Bone marrow obtained progenitor initiate the pre metastatic niches. Nature 438(7069): 820-827.
-
Erler JT, Bennewith KL, Cox TR, Lang G, Bird D, et al. (2009) Hypoxia –induced lysyl oxidase is a critical mediator of Bone marrow cell recruitment to form the pre metastatic niches. Cancer Cell 15(1): 35-44.
-
Pavlaki M, Zucker S (2003) Matrix metalloproteinase inhibitors (MMPIs): the beginning of phase Ior the termination of phase III clinical trials. Cancer Metastais Rev 22(2-3): 177-203.
-
Zeng ZS, Cohen AM, Zhang ZF, Stetlier SW, Guillem JG (1995) Elevated Tissue inhibitors of Metalloproteinases-1 RNA in colorectal cancer stroma correlates with lymph node and distant Metastais. Clin Cancer Res 1(8): 899- 906.
-
Troeberg L, Lazenbatt C, Anower EKMF, Freeman C, Federov O, et al. (2014) Sulphated glycosaminoglycans control the extracellular trafficking and the activity of the metalloproteinase inhibitors TIMP-3. Chem Biol 21(10): 1300-1309.
-
Bachman KE, Herman JG, Corn PG, Merlo A, Costello JF, et al. (1999) Methylation associated silencing of Tissue inhibitors of Metalloproteinases-3 gene suggests a suppressor role in kidney, brain, and other human Cancers. Cancer Res 59(4): 798-802.
-
Baker AH, George SJ, Zaltsman AB, Murphy G, Newby AC (1999) Inhibition of invasion and induction of apoptotic cell death of cancer lines by overexpression of TIMP-3. Br J Cancer 79(9-10): 1347-1355.
-
Anand AB, Bao L, Smith R, Iwata K, Olsen BR, et al. (1996) A review of Tissue inhibitors of Metalloproteinases -3 and experimental trafficking and its effect of the primary tumor growth. Biochem Cell Biol 74(6): 853-862.
-
Sharma S, Free A, Mei Y, Pelper SC, Wang Z, et al. (2010) Distinct Molecular signatures in pediatric infratentorial glioblastomas defined by aCGH. Exp Mol Pathol 89(9): 169-174.
-
Gu P, Xing X, Tanzer M, Rocken C, Weichert W, et al. (2008) Frequent loss of TIMP-3 expression of in progression of esophageal and gastric adenocarcinomas. Neoplasia 10(6): 563-572.
-
Wang Y, Zhang D, Zheng W, Luo J, Bai Y, et al. (2008) Multiple gene methylation of nonsmall lung cell cancers evaluated with 3–dimensional microarray. Cancer 112(6): 1325-1336.
-
Guan Z, Zhang J, Song S, Dai D (2013) Promoter methylation and expression of TIMP-3 gene in gastric cancer. Diagn Pathol 8: 110.
-
Voland P, Besig S, Rad R, Braun T, Baur DM, et al (2009) Correlation of Matrix metalloproteinase and Tissue inhibitors of Metalloproteinase expression in ileal carcinoids, lymph nodes and liver metastases with progress and survival. Neuroendocrinology 89(1): 66- 78.
-
Jackson HW, Defamie W, Waterhouse P, Khokha R (2017) TIMPs: Versatile regulators in cancer. Nat Rev Cancer 17(1): 38-53.
-
Pulukuri SM, Pratibandla S, Patel J, Es- tes N, Rao JS (2007) Epigenetic in activation of the ex- pression Tissue inhibitors of Metalloproteinase-2 (TIMP-2) gene in human prostate tumors. Oncogene 26: 5229-5237.
-
Chowdhary A, Brinson R, Wei B, Stetler SWG (2017) Tissue inhibitors of Metalloproteinase-2 (TIMP-2): Bioprocess, Development, Physicochemical, Biochemical, and biologic characteristization of highly expressed recombinant protein. Biochemistry 56(49): 6423-433.
-
Zhu L, YuH, Liu SY, Xiao XS, Ding WH, et al. (2015) Prognostic value of Tissue inhibitors of Metalloproteinase-2 expression in patients with non- small cell lung Cancer: Systematic review and meta analysis. PLoS ONE 10(4): e0124230.
-
Kubben FJG, Sier CFM, Meijer MJW, Van den Berg M, Van den Reijden JJ, et al. (2006) Clinical impact of MMP and TIMP gene polymorphisms in gastric cancer. Br J Cancer 95(6): 744-751.
-
Hagemann T, Gunawan B, Schulz M, Fuzesi l, Binder C (2001) mRNA expression of Matrix metalloproteinases and their inhibitors differ in subtypes of renal cell carcinomas. Eur J Cancer 37(15): 1839-1846.
-
Liss M, Sreedhar N, Kesh Gegian A, Sauter G, Chernick MR, et al. (2009) Tissue inhibitors of Metalloproteinase-4 is elevated in early stage breast Cancers with accelerated progression and poor clinical course. Am J Pathol 175(3): 940-946.
-
Rorive S, Lopez XM, Maris C, Trepant AL, Sauvage S, et al. (2010) TIMP4 and CD63:New prognostic Biomarkers in human astrocytoma. Mod Pathol 23(10): 1418-1428.
-
Hanahan D, Weinberg RA (2011) Hallmarks of Cancer: the next generation. Cell 144(5): 646-674.
-
Egeblad M, Werb Z (2002) New functions for the Matrix metalloproteinases in Cancer progresion. Nat Rev Cancer 2(3): 161-174.
-
Peschon JJ, Slack JL, Reddy P, Stocking KL, Sunnarborg SW, et al. (1998) An essential role for ectodomain shedding in mammalian Development. Science 282(5392): 1281- 1284.
-
Rajasinghe LD, Pindiprolu RH, Gupta SV (2018) Delta– trocotrienol inhibits non-small-cell lung cancer cell invasion via the inhibition of NFκB, UPA Activator and MMP-9. Oncotergets Ther 11: 4301-4314.
-
Yamashita K, Suzuki M, Iwata K, Koike T, Hamaguchi M, et al. (1996) Tyrosine phosphorylation is crucial for growth signalling by Tissue inhibitors of Metalloproteinase (TIMP-1 and TIMP-2). FEBS Lett 396(1): 103-107.
-
D’Alessio S, Ferrari G, Cinnante K, Scheerer W, Galloway AC, et al. (2008) Tissue inhibitors of Metalloproteinase- 2(TIMP-2) binding to membrane type1 Matrix metalloproteinases induces MAPK activation and cell growth by a non proteinolytic mechanism. J Biol Chem 283(1): 87-99.
-
Hoegy SE, Oh HR, Corcoran ML, Stetler SWG, Liotta LA, et al. (2008) Tissue inhibitors of Metalloproteinase-2 (TIMP-2) suppresses TKR-growth factor signalling independent of Matrix metalloproteinases inhibition. J Biol Chem 276(5): 3203-3214.
-
Gioleli C, Theocharis AD, Karamanos NK (2011) Roles of Matrix metalloproteinases in Cancer progression and their pharmacological targeting. FEBS J 278(1): 16-27.
-
Waldhauer I, Goehlsdorf D, Gieseke E, Weinschenk T, Wittenbrink M, et al. (2008) Tumor–associated MICA is shed by ADAM proteases. Cancer Res 68(15): 6368- 6376.
-
Jung KK, Liu XW, Chirco R, Fridman R, Kim HR (2006) Identification of CD63 as Tissue inhibitors of Metalloproteinase-1 interacting cell surface protein. EMBO J 25(17): 3934-3942.
-
Stetler SWG (2008) Tissue inhibitors of Metalloproteinases in cell signalling: metalloproteinase- independent Biological activities. Sci signal 1(27): 6.
-
Li H, Nishio K, Yamashita K, Hayakawa Y, Hoshino T (1995) Cell Cycle–dependent localization of Tissue inhibitors of Metalloproteinase-1 immunoreactivity in cultured human gingival fibroblasts. Nagoya J Med Sci 58(3-4): 133-142.
-
Groft KK, Muzik H, Rewcastle NB, Johnston RN, Knauper V, et al. (2001) Differential expression and localization of TIMP-1 and TIMP-4 in human gliomas. Br J Cancer 85(1): 55-63.
-
Guo YH, Gao W, Li Q, Li PF, Yao PY, et al. (2004) Tissue inhibitors of Metalloproteinase-4 suppresses vascular smooth muscle cell migration and induces cell apoptosis. Life Sci 75(20): 2483-2493.
-
Smith MR, Kung H, Durum SK, Colburn NH, Sun Y (1997) TIMP-3 induces cell death by stabilizing TN Falpha receptors on the surface of human colon carcinoma cells. Cytokine 9(10): 770-780.
-
Ahonen M, Poukkula M, Baker AH, Kashwagi M, Nagase H, et al. (2003) Tissue inhibitors of Metalloproteinase-3 induces apoptosis in melanoma cells by stabilization of death receptors. Oncogene 22(14): 2121-2134.
-
Alexius Lindgren M, Andersson E, Lindstedt L, Engstrom W (2014) RECK gene and Biological malignancy-–its significance in angiogenesis and inhibition of Matrix metalloproteinases induces. AntiCancer Res 34(8): 3867-3873.
-
Eiro N, Gonzalez L, Martinez Ordonez A, Fernandez Garcia B, Gonzalez LA, et al. (2018) Cancer associated fibroblasts affect breast Cancer gene expression, invasion and angiogenesis. Cell Oncol 41(4): 369-378.
-
Koolwijk P, Sidenius N, Peters E, Sier CF, Hanemaaijer R, et al. (2001) Proteolysis of the urokinase type plasminogen activator receptor by metalloproteinase-12: implication for angiogenesisin fibrin matrices. Blood 97(1): 3123-3131.
-
Handsley MM, Edwards D (2015) Metalloproteinases and their inhibitors in tumor angiogenesis. Int J Cancer 115(6): 849-860.
-
Wurtz SO, Schrohl AS, Sorensen NM, Lademann U, Christensen IJ, et al. (2005) Tissue inhibitors of Metalloproteinase-1 in breast Cancer. Endocr Relat Cancer 12(2): 215-227.
-
Feldman AL, Stetler Stevenson WG, Costouros NG, Knezevic V, Baibakov G, et al. (2004) Modulation of tumor-host inter actions, angiogenesis, and tumor growth by Tissue inhibitors of Metalloproteinase-2 via a novel mechanism. Cancer Res 64(13): 4481-4486.
-
Oh HR, Takahashi R, Kondo S, Mizoguchi A, Adashi E, et al. (2001) The membrane anchored MMP inhibitor RECK is a key regulator of extra cellular Matrix integrity and angiogenesis. Cell 107(6): 789-800.
-
Fernandez CA, Butterfield C, Jackson G, Moses MA (2003) Structure and functional uncoupling of the enzymatic and angiogenic inhibitoryactivities of Tissue inhibitors of Metalloproteinase-2(TIMP-2): Loop 6 is a novel angiogenic inhibitor. J Biol Chem 278(42): 40989- 40995.
-
Qi JH, Ebrahem Q, Moore N, Murphy G, Claesson Welsh L, et al. (2003) A novel function for Tissue inhibitors of Metalloproteinase-3(TIMP-3): inhibition of angiogenesis by blockage of VEGF binding to VEGF- receptor2.Nat Med 9(4): 407-415.
-
Kang KH, Park SY, Rho SB, Lee JH (2008) Tissue inhibitors of Metalloproteinase-3 interacts with angiotensin II type 2 receptor and additively inhibits angiogenesis. Cardiovascular Res 79(1): 150-160.
-
Saunders WB, Bohnsack BL, Faske JB, Anthis NJ, Bayless KJ, et al. (2006) Coregulation of vascular tube stabilization by endothelial cell TIMP-2 and pericyte TIMP-3. J Cell Biol 175(1): 179-191.
-
Radisky ES, Raeeszadeh Sarmazadeh M, Radisky DC (2017) Therapeutic potential of Matrix metalloproteinase: inhibition in breast Cancer. J Cell BioChem 118(11): 3531-3548.
-
Bosch R, Philips N, Suarez Perez JA, Juarranz A, Devmurari A, et al. (2015) Mechanisms of photoaging and cutaneous photocarcinogenesis, and photoprottective strategies with phytochemicals. Antioxidants 4(2): 248- 268.
-
Radisky ES, Radisky DC (2010) Matrix metalloproteinase- epithelial-mesenchymal transition in breast Cancer. J Mammary Gland Biol Neoplasia 15(2): 201-212.
-
D Angelo RC, Liu XW, Najy AJ, Jung YS, Won J, et al. (2014) TIMP-1 via TWIST 1 induces EMT phenotypes in human breast epithelial-cells. Mol Cancer Res 12(9): 1324-1333.
-
Ho AT, Voura EB, Soloway PD, Watson KL, Khokha R (2001) MMP inhibitors augment fibroblasts adhesion through stabilization of focal adhesion contacts and upregulation of cadherin function. J C Biol Chem 276(43): 40215-40224.
-
Hojilla CV, Kim I, Kassiri Z, Fata JE, Fang H, et al. (2007) Metalloproteinase Axes Increases beta-catenin signalling in primary Mammary epithelial cells lacking TIMP-3. J Cell Sci 120(Pt-6): 1050-1060.
-
Barker HE, Chang J, Cox TR, Lang G, Bird D, et al. (2011) LOXL2 mediated Matrix remodeling in metastases and Mammary gland involution. Cancer Res 71(5): 1561-1572.
-
Malanchi I, Huelsken J (2009) Cancer Stem cells: never Wnt away from the niche. Curr Opin Oncol 21(1): 41-46.
-
Hancox RA, Allen MD, Holliday DL, Edwards DR, Pennington CJ, et al. (2009) Tumor-associated tenascin-C isoforms promote breast Cancer cell invasion and growth by Matrix metalloproteinase dependent and in dependent mechanisms. Breast Cancer Res 11(2): 24.
-
Nissinen L, Kahri VM (2014) Matrix metalloproteinase in inflammation. Biochim Biophys Acta 1840(8): 2571-2580.
-
Brew K, Nagase H (2010) The Tissue inhibitors of Metalloproteinase (TIMPs): An ancient family with structural and functional diversity. Biochim Biophys Acta 1803(1): 55-71.
-
Wu DW, Tsai LH, Chen PM, Lee MC, Wang L, et al. (2012) Loss of TIMP-3 promotes tumor invasion via elevated IL-6 production and predicts relapse in HPV infected non-small cell lung Cancer. Am J Pathol 181(5): 1796-1806.
-
Folgueras AR, Pendas AM, Sanchez LM, Lopez Ontin C (2004) Matrix metalloproteinase in Cancer: from new functions to improved inhibition strategies. Int J Dev Biol 48(5-6): 411-424.
-
Trojanek JB (2015) Role of Matrix metalloproteinase in hypertension: pathogenesis of obesity and hypertension. Postepy Biochem 61(4): 356-363.
-
Nagao Y, Hisaoka M, Matsu yama A, Kanemitsu S, Hamada T, et al. (2012) Association of miR-21 expression with its targets .PDCD4 and TIMP-3 in pancreatic ductal adenocarcinoma. Mod Pathol 25: 112-121.
-
Overall C (2002) Molecular determinants of Matrix metalloproteinase substrate specificity: Matrix metalloproteinase substrate binding domains, nodules and exodsites. Mol Biotechnol 22(1): 51-86.
-
Vandenbroucke RE, Libert C (2014) Is there new hope for therapeutic Matrix metalloproteinases inhibition. Nat Rev Drug Discov 13(12): 904-927.
-
Timokhina E, Strizhakov A, Ibragimova S, Gitel E, Ignatvo I, et al. (2020) Matrix metalloproteinases MMP2 and MMP9 occupy a new role in severe Preeclampsia. J Pregnancy 8369645.
-
Kaur KK, Allahbadia GN, Singh M (2013) An update on role of human prokineticins in human reproduction- potential therapeutic implications. Published in Open J Genet 3: 201-15.
-
Su MT, Tsai PY, Tsai HL, Chen Y, Kuo PL (2017) miR- 346 and miR-582-3p-regulated EG-VEGF expression and trophoblast invasion via matrix metalloproteinases 2 and 9. BioFactors. 43(2):210–219.
-
Kaur KK, Allahbadia GN, Singh M (2020) How does Epigenetics Regulate Development of Placenta and Placental Pathologies like Preeclampsia (PE), Intrauterine growth Restriction (IUGR)-With Main emphasis on PE’’. Advances in Bioengineering and Biomedical Science Research.
-
Kaur KK, Allahbadia GN, Singh M (2020) Utilization of Extracellular Vesicles for Treatment of Type 1 Diabetes Mellitus (T1DM) Along with Type 2 Diabetes Mellitus (T2DM) besides Complications Associated with Diabetes- A Systematic Review. J Clin Diabetes Obes 1: 001-013.
-
Kaur KK, Allahbadia GN, Singh M (2020) An Update on Future Utilization of Extracellular Vesicles in Evaluation Part of These in Maturation of Gametes, Fertilization along with Embryo Implantation for Escalating the Success of Artificial Reproductive Technology: A Systematic Review. Open Access Journal of Gynecology and Obstetrics 3(2): 10-29.
-
Ding J, Huang F, Wu G, Han T, Xu F, et al. (2015) miR-519-d-3p suppresses invasion and migration of trophoblast cells via targeting MMP2. PLoS 10(3): e0120321.
-
Chaiwangyen W, Murrieta CJM, Favaro RR, Photini SM, Gutierrez SRN, et al. (2020) MiR-519d-3p in Trophoblastic Cells: Effects, Targets and Transfer to Allogeneic Immune Cells via Extracellular Vesicles. Int J Mol Sci 21(10): 3458.
-
Choi EJ, Kil IS, Cho EG (2020) Extracellular Vesicles Derived from Senescent Fibroblasts Attenuate the Dermal Effect on Keratinocyte Differentiation. Int J Mol Sci 21(3):1022.
-
Wen H, Hu Y, Chen L, Zhao L, Yang XY (2020) miR- 338-5-p targets Epidermal growth factor containing fibulin like extracellular matrix protein 1 to inhibit the growth and invasion of trophoblast cells in intrauterine growth restriction. Reprod SCI 27(6): 1357-1364.
-
Li Y, Huang Y, Qi Z, Sun T, Zou Y (2018) miR-338- 5-promotes glia cell invasion by regulating TSHZ3and MMP2. Cell Mol Neurobiol 38(3): 669-677.
-
Giessler F, Ender F (2020) Extracellular Vesicles and cell-cell communication: New insights and new therapeutic strategies not only in Oncology. Int J Mol Sci 21(12): 4331.
-
Awamleh Z, Han VKM (2020) Identification of miR-210-5p in human placentae from pregnancies complicated by preeclampsia and intrauterine growth restriction, and its potential role in the pregnancy complications. Pregnancy Hypertens, pp: 159-168.
-
Swarnakar S, Paul S (2009) Curcumin arrests endometriosis by downregulation of matrix metalloproteinase-9 activity. Indian J Biochem Biophys 46(1): 59-65.
-
Jana S, Rudra DS, Paul S, Snehasikta S (2012) Curcumin delays endometriosis development by inhibiting MMP-2 activity. Indian J Biochem Biophys 49(5):342-348.
-
Paul S, Bhattacharya P, Das Mahapatra P, Swarnakar S (2010) Melatonin protects against endometriosis via regulation of matrix metalloproteinase-3 and an apoptotic pathway. J Pineal Res 49(2): 156-68.
-
Grzechocinska B, Dabrowski FA, Chlebus M, Gondeka A, Czarzasta K, et al. (2018) Expression of matrix metalloproteinase enzymes in endometrium of women with abnormal uterine bleeding. Neuro Endocrinol Lett 38(8): 537-543.
-
Johans M, Lemoine P, Janssens V, Giuseppina G, Nielsen R, et al. (2017) Cellular uptake of proMMP-2: TIMP-2 complexes by the endocytic receptor megalin/ LRP-2. Sci Rep 7: 4328.
-
Yosef G, Arkadash V, Papo N (2020) Targeting the MMP-14/MMP-2/integrin αvβ3 axis with multispecific N-TIMP2–based antagonists for cancer therapy. J Biol Chem 293(34): 13310-13326.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’