Are we any Close to Utilizing Targeting SIRT Signaling Pathway for Enhancing the Oocyte Quality in Women with Advanced Maternal age: Bringing from Bench to Bedside: A Narrative Review
Earlier we had reviewed the role of sirtuins regarding SIRTs in the form of epigenetic modifiers regarding postponement of Diabetic Kidney Disease (DKD) apart from in placental growth in pregnancy besides how resveratrol could be used in numerous chronic inflammatory diseases and Autoimmune Diseases to target SIRT1. Furthermore, we had reviewed the role of mitochondrial function in reproductive endocrinology as well as its significance in sustenance of oocyte quality. Recently lot of work has been conducted to improve the oocyte quality in women presenting with advanced age or those coming in with diminished ovarian reserve (DOR) with use of various anti-oxidant along with anti-inflammatory agents like melatonin, resveratrol. Thus we conducted a narrative review utilizing search engine PubMed, google scholar ;web of science; embase; Cochrane review library utilizing the MeSH terms like Sirtuins; maternal ageing; Oxidative stress (OS); organs ageing; oocytes quality; embryo quality; implantation; mitochondria function; Epigenetics; DNA methylation; Histone deacetylation; Histone crotonylation; melatonin; resveratrol; nicotinamide mononucleotide; nicotinamide riboside; from1990 to 2023 till date. We found a total of 300 articles out of which we selected 110 articles for this review. No meta-analysis was done. We observed Oxidative stress to be the mode implicated in mitigating oocyte quality. Escalated corroboration from animal model in addition to Clinical studies have validated that Sirtuins confer protection from OS aiding in improvement of oocyte quality through an anti-oxidant action .Thus escalating research has concluded that Sirtuins possess protective part to oocyte quality. Soon there might be a time with the rate at which this research in mounting women will not need to cryopreserve gametes only due to choosing professional carreers.
Introduction
Dysfunctional oocyte quality has been acknowledged to be a key factor influencing female infertility. The reduction of oocyte quality is usually generated in view of oxidative injury correlated with maternal ageing [1]. In view of alterationsin cultural as well as social milieu women tend to postpone fertility at a much later age in particular in west with women pursuing careers, even catching up here in India. Sequentially, a greater insight is required regarding age correlated oocyte quality dysfunction has evoked interest recently. Despite, lack of full insight regarding mode of dysfunctional oocyte quality variable constituents have been pointed to be correlated with this event [2, 3]. Oxidative stress (OS) that takes place secondary to Reactive oxygen species (ROS) at the time of ovarian ageing has been indicated in the form of a crucial factor [2], in the reduction of oocyte quality, possessing properties of mitochondrial impairment along with dysfunctional ovarian microenvironment [3].
Sirtuins, reflect a family of NAD+/ nicotinamide adenine dinucleotide (NAD+)-based protein deacetylases along with ADP ribosyl transferases. These have been revealed to be key controllers of genome sustenance, ageing in addition to chromosomal intactness of oocyte [4]. Escalating proof has validated that interplay amongst ROSas well as sirtuin family possesses crucial part in controlling the cellular ageing events. Sirtuins, further have been illustrated to confer protection from OS [5]. These proteins might be probable markers regarding ovarian ageing along with SIRT1, SIRT3, as well as SIRT6are acknowledged to be target molecule for postponement of organs ageing [6]. Earlier we had reviewed part of SIRTs in the form of epigenetic modifiers regarding postponement of Diabetic Kidney Disease (DKD) & in placental growth in pregnancy beside showed how resveratrol could be used in numerous chronic inflammatory diseases and autoimmune Diseases [7, 8, 9]. Here we have tried to review the significance of SIRT signaling pathway regarding improving oocyte quality.
Methods
Here we conducted a narrative review utilizing search engine PubMed, google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like Sirtuins; maternal ageing; Oxidative stress (OS); organs ageing; oocytes quality; embryo quality; implantation; mitochondria function; Epigenetics; DNA methylation; Histone deacetylation; Histone crotonylation; melatonin; resveratrol; nicotinamide mononucleotide; nicotinamide riboside; from1990 to 2023 till date.
Results
We found a total of 300 articles out of which we selected110 articles for this review. No meta-analysis was done.
Oocyte Quality Dysfunction Secondary to Ageing
With ageing slow but steady elimination of follicles along with reduction in ovarian oocyte quality possessing properties of incapacity of generating oocyte which possess competence for getting fertilized followed by embryonic formation. OS has been acknowledged to be the causative modulators impacting oocyte quality via 3 separate stages; i) escalated ROS formation, ii) antioxidant mobilization iii) Oxidative injury to main targets. The oocyte quality gets impacted by escalated ROS via telomere length shortening, impacting DNA injury as well as mutation induction. Mitochondria reflect the main formers of ROS with accrual of errors in mitochondria results in apoptosis that interferes with follicles generation in mammals [10]. Moreover, ROS might activate nuclear factor κB (NFκB), as well asstimulate inflammatory substances, intercom γ (IFN-γ), along with lipopolysaccharide (LPS), that escalate ROS formation in synergism [11].
The maximum acknowledged posit of oocyte quality dysfunction secondary to ageing is the inimical mitochondrial function secondary to ROS. Mitochondria have been believed to be the organelles possessing finite part in oocyte working. They reflect the major sources of energy generation as adenosine triphosphate (ATP), needed for various events, inclusive of assembling of meiotic spindle, chromosomal separation, fertilization along with following embryogenesis in mammals [12]. The lesser quantities of mitochondria/ dysfunctional mitochondrial working lead to the decreasing energy needed for meiosis at the time of oocyte maturation which in all probability results in chromosomal abnormalities in mice as well as humans [13]. Mitochondria further participate in various intracellular events needed for appropriate embryo generation inclusive ofCa2+homeostasis event, apoptosis control along with OS managing in mice as well as humans [2, 14]. At the time of aerobic metabolism mitochondria generate byproducts which can’t be avoided like ROS [15]. Moderate quantities of ROS are necessary for various intracellular events inclusive of ventilation, nerve transmission as well as immune controlling events [16]. Nevertheless, escalated ROS accrual secondary to reduction of ageing correlated cellular respiration functions can injure cellular lipids, proteins along with DNA impacting their normal working [15]. This OS impacts telomere function resulting in deletions or point mutations in them it chondral genome, reduction in ATP formation, meiotic spindleaberrations, genomic non stability, leading to the oocyte losing competence. It has been documented that mitochondrial DNA(mt DNA)copy numbers along with oocyte working in ageing mice as well as women are decreased correlated with greater chances ofaberrations in spindle assembly as well as chromosomal separation [13, 17]. Sequentially these result in chromosomal separation aberrations, maturation as well as fertilization failures, or oocyte/ embryo fragmentation, in mice [18] as well as humans [19]. In the case of human ageing oocytes, a reduction in the mitochondrial components along with escalated matrix density occur [20]. The reduction of mitochondrial respiration function correlated with ageing of granulosa cells (GC’s) stimulates the escalated ROS formation in mice [21]. Moreover, escalation of oxidatively injured cellular lipids, proteins along with DNA are present in variable ovarian partitions, inclusive of murine stromal tissue as well as changes of antioxidant - eenzymes expression [21].
The proposed etiology for OS is age correlated dysfunctional follicular vascularization [22]. A considerable important negative association among stage along with ovarian perifollicular blood flow was seen in the follicular phase in view of accrual of advanced glycation end- products (AGE) in follicular fluid at the time of ovarian ageing in mice as well as humans [22]. The granulosa (GC) and theca cells escalate the generation of vascular endothelial growth factors (VEGF) to adjust to this situation, that however have in capacity of finishing enough reaction. Probably this takes place in view of lesser reactivity of endothelial cells, inhibiting ovarian stroma vessels or escalating space amongst perifollicular blood vessels in the ageing ovary [22].
Reasoning offered for poor oocyte quality inhuman ovarian ageing is the interference of the connection amongst GC along with oocyte impact the follicular production. Oocytes liberate variable paracrine growth factors like Bone morphogenetic protein9 (BMP9) as well as Growth Differentiation factor-15 that are implicated in proliferation, differentiation along with possess crucial part in follicular generation as well as maturation [10, 23]. In the case of human oocyte senescence is further documented to be correlated with telomere length shortening in cumulus cells (CC), whose length has been acknowledged to be an embryo quality marker [24]. CC’s obtained from infertility patients having in vitro fertilization (IVF) have illustrated escalated apoptosis with correlated with considerably lesser fertilization in addition to pregnancy rates [10].
Part of SIRT Signaling Pathway Regarding Oocyte Quality
Cellular SIRT Signaling Pathway: Sirtuins, (or silent information regulator 2 family- SIRT family) represent proteins of the NAD+-based protein deacetylases family. They are part of Class IIIhistone deacetylates (HDACs) possessing the properties of catalytic actions of enzymes based on NAD+/NADH ratio [25]. The major effect of Sirtuins is protein acetylation, a significant post-translational modifications correlated with controlled various metabolic signaling pathways in mammals [4]. Sirtuins have been detailed in the form of crucial controllers innumerous cellular events inclusive of apoptosis [26], cell reprogramming [26], DNA healing [27], redox homeostasis [28], energy metabolism [29], tumorigenesis [29]. Moreover they possess a key part regarding intracellular OS sensing through modulating the NAD+/NADH ratio [30] (Figure1) [31].
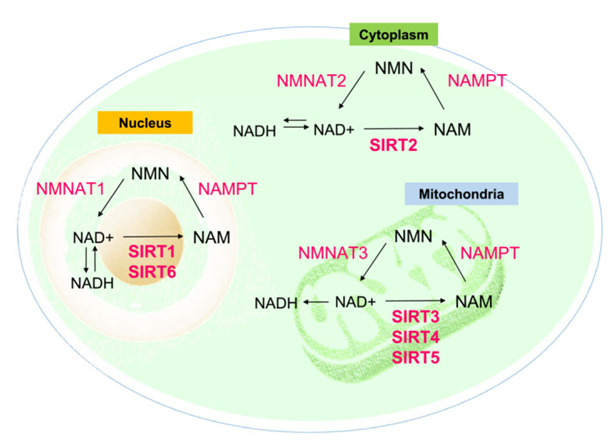
Figure 1: Courtesy ref no-31-Roles of SIRT1‐7 in sensing intracellular oxidative stress via modulating the NAD+/ NADH ratio in different cell organelles. NAD+, oxidative nicotinamide adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide; NAM, nicotinamide; NAMAT1,2,3, nicotinamide mononucleotide adenylyl transferase 1,2,3; NAMPT, nicotinamide phosphoribosyl transferase; NMN, nicotinamide mononucleotide.
In case of mammals there are 7Sirtuin family members observed in mammals (SIRT 1-7) [32]. Out of this SIRT 1 along with SIRT 2 exist in nucleus as well as cytoplasm. SIRT 3 is basically placed in mitochondria along with nucleus [4]. SIRT 4, SIRT5 along with SIRT 6, represent mitochondrial sirtuins, whereas SIRT 6, SIRT 7are located in nucleus] (Figure1) [4, 29]. Apart from protein acetylation, certain sirtuin said in various cellular events through other actions. Like the actions of SIRT 4, SIRT 6 does not implicate deacetylation, however ADP ribosylation [33], while SIRT5 possess the capacity of demalonylase along with desuccinylase proteins [34].
Of the members of sirtuin family, part of SIRT 1 is maximum detailed, in particular for its antiaging actions. On getting activated by stressful processes, binding of SIRT 1to various molecular targets along with deacetylases these molecules for controlling energy homeostasis, gene silencing, metabolism, genomic stability as well ascell survival [29, 32, 35]. Like SIRT 1 causes activation of AMP- activated protein kinase (AMPK), resulting in escalated mitochondrial working as well as transcriptional action [36]. SIRT1 deacetylates the Werner helicases along with NBS1that heal DNA injury [37]. Escalated chromosomal abnormalities as well as dysfunctional DNA injury healing takes place in the case of SIRT1-/- mice [38]. Furthermore SIRT1 changes gene injury at the time of mammalian ageing event aiding in the genomicintactness as well asage correlated alterations in gene expression [39]. SIRT1 further possesses conclusive part regarding cell fate by crosstalk with variable target constituents that modulate the apoptosis threshold like fork head box proteinO3a(FOXO3a),p53,Ku70,E2F1, NFκB complex, as well as transforming growth factor beta (TGF-β) signaling[29, 35]. Additionally, by controlling p53 action SIRT1 probably acts in the form of tumor promoteror repressor based on particular situations [40]. The other target of SIRT1 is Peroxisome Proliferator Activated Receptor γ coactivator -1α (PGC-1 α) which possesses a key part in gluconeogenesis as well as lipid metabolism in a cell line obtained from pheochromocytoma of the rat adrenal medulla along with hepatocytes [41].
The Part of SIRT Signaling Pathway Regarding Oocyte Quality
In the case of mammals this sirtuins family has got illustrated to attenuate oocyte quality by controlling the redox status [30]. In the meantime dysfunctional SIRT working leads to the fertility deficiencies [30, 32]. Figure 2 & 3 shows the probable part of SIRT signaling pathways regarding oocyte quality.
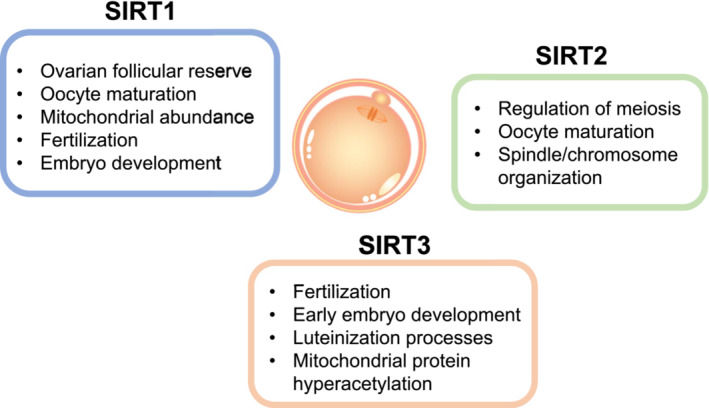
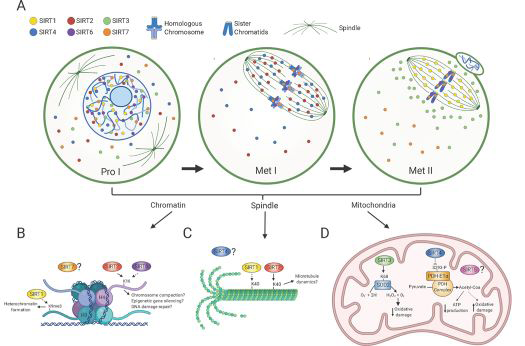
Figure 3: Courtesy ref no -5-Sirtuin distributions in oocytes during meiotic divisions and its associated molecular functions. A) Scheme denotes Sirtuin localization in oocytes at prophase I (Pro I) and at Metaphase I and II (Met I, Met II). SIRT1 is mostly nuclear in prophase I-arrested oocytes and on the spindle in Met II-arrested eggs. SIRT2 is found throughout the ooplasm and nucleus in prophase I and relocalizes to the spindle in Met I. SIRT6 and SIRT7 are chromatin-bound at all developmental stages, but SIRT7 is also present to a lesser extent in the ooplasm. SIRT3 and SIRT4 are predominantly nuclear in prophase I. SIRT3 redistributes around the spindle in Met II eggs and SIRT4 reallocates to the spindle in Met I. The cellular distribution of SIRT5 in oocytes is currently unknown. B-D) Molecular functions of Sirtuins during meiotic maturation in chromatin (B), the spindle (C) and mitochondria (D). B) SIRT1 promotes H3K9me3 deposition and heterochromatin formation and SIRT2 and SIRT6 regulates H4K16ac levels with potential implications in chromosome compaction, gene silencing and DNA damage repair signaling. C) SIRT1 and SIRT2 both regulate acetylation of α-tubulin suggesting a potential role in spindle dynamics. D) SIRT3 stimulates SOD2 activity and ROS balance and SIRT4 limits Pyruvate Dehydrogenase (PDH) complex activity leading to reduced ATP production. Solid and dashed arrows denote potential direct and indirect roles, respectively.
Role of SIRT1: The initial broadly detailed constituent that got acknowledged in the form of crucial transcription factor robustly responsible for ageing as well as lifespan is SIRT1.In mice expression of SIRT1 takes place at mRNA along with protein levels in ovarian tissue as well as luteinized granulosacells (GCs) assuming apart of meiosis chaperone [42]. It was pointed that SIRT1 decreased the ageing stimulated deficiencies inclusive of murine oocyte morphological alterations, ROS accrual, spindle morphology in addition tomato chondral working [43]. Outcomes obtained from mice experiments pointed apart of SIRT1 regarding adaptive reaction to OS along with confer protection against elimination of generational competence with reproductive along with post ovulatory ageing through SIRT1- FOXO3a [44] axis. Hampering SIRT1 escalatesROS quantities as well asinterferes with the spindle getting organized in murine oocytes [42]. SIRT1is further implicated in nutritional status along with controller of cell cycle at the time of folliculo genesis as well asluteinization events [38]. Moreover, SIRT1 modulates adaptive reaction to OS in murine oocytes by manipulating antioxidant enzymatic reaction [42]. Hampering of SIRT1 by a particular hampering agent, Ex527 leads to the upregulation of MnSod gene as well as avoid of ROS in various cell kinds [45]. Akin to that hampering of bovine oocytes by Ex527, avoided robust activation of primordial follicles (PMF’s), stimulated by cyclophosphamide. Ex527 therapy for 3 days possessed the capacity of decreasing the overexpression of SIRT1 stimulated by cyclophosphamide [46]. These outcomes pointed thatSIRT1 might be a probable molecule which might be targeted regarding avoidance of deletion of the dormant PMF’s secondary to cyclophosphamide. In case of ageing murine oocytes interference of SIRT1- FOXO3a- MnSod pathway in addition to reduction of miR132, a corroborated SIRT1modulator was seen [52]. While in vitro maturation assay outcomes pointed thatSIRT1 controlled redox status guaranteeing normal assembling of spindles at the time of oocyte maturation [42]. Study revealedSIRT1 activation by resveratrol decreased the piercing capacity of maternal ageing correlated deficienciesin murine oocytes. On adding resveratrol to maturation medium regulation SIRT1 protein expression in oocytes escalated, which escalated ATP quantities in matured oocytes, the total mitochondrial quantities as well as mitochondrial membrane potential, ratio of normal fertilization in addition to total blastocysts alterations in oocytes numbers [47]. Mice treated with rapamycin 5mg/kg/day for generating caloric limitations phenotypes right through mammalian target of rapamycin inhibitors (mTOR) repression depicted escalated follicle reserve along with enhanced SIRT1as well as SIRT6 [48]. In case of bovine hampering of SIRT1 lead to the greater ratio ofaberrant fertilization [49]. In contrast to young bovines oocytes obtained from ageing cows demonstrated greater quantities of aberrant fertilization along with blastocysts with lesser full cell numbers [49]. In case ofmurine oocytes which are aged, the quantities of NAD+as well as nicotinamide adenine mononucleotide adenylyl transferases (NMNAT2) were reduced along with the particular elimination of NMNAT2 in oocytes enhanced meiotic aberrations in addition to metabolism impairment. These abnormal phenotypes might be rescued by over expression or activation of SIRT1. It further displayed the significance of NAD+/ SIRT1 axis in modulating the actions of NMNAT2 on oocyte quality regulation [50]. Akin observations were documented in a different study which illustrated that the elimination of oocyte- SIRT1axis impacted ovarian follicular reserve, oocyte maturation, oocyte mitochondrial enrichment, OS, fertilization, embryo generation along with fertility at the time of ageing in mice. Noticeably, elimination of this crucial sirtuins from growing oocytes did not have impact in young ladies [51]. Role of SIRT2: Regarding SIRT2, it possesses significant part in metabolism by deacetylating cytoplasmic transcription factor along with retransferring them to nucleus [4]. A particular part of SIRT2 is peripheral myelination by facilitating arborization along with downstream expression of myelin particular genes, with demonstration of its part in neurological function[52].The acetylation status of tubulin is modulated by SIRT2in a NAD+-based way [53]. In case of mouse oocytes, more spindles along with chromosomal deficiencies as well as dysfunctional microtubule-kinetochore crosstalk were observed subsequent to elimination of SIRT2 action [54]. Additionally, lesser protein quantities of SIRT2 were seen in oocytes from ageing mice, pointing that reduction ofSIRT2 might be a factor aiding in the oocyte-age based deficiencies [54]. SIRT2 based BubR1 deacetylation was illustrated to control meiotic apparatus in normal oocytes along with modulate the action of maternal age on oocyte quality [55]. The elimination of SIRT2 interferes with maturational propagation as well asspindle/chromosome distribution in addition to disrupted kinetochore- microtubule connections [55]. The escalated SIRT1 along with SIRT2 induction taking place subsequent to the antioxidant N-acetylcysteine (NAC) might postpone oocyte ageing in mice [56]. Role of SIRT3: SIRT3, possessing maximum expression in metabolic tissues (like, liver, kidney, heart) occurs in lipidas well as glucose metabolism in mammals [4]. It controls the acetylation of various key metabolic enzymes inclusive of acetyl-CoA synthetase long chain acyl CoA dehydrogenase, 3 hydroxy, 3methyl glutaryl-coenzyme Asynthase-2 [8]. The hyper acetylation of mitochondrial protein resulting from knockout of SIRT3geneis correlated with a great risk of generation of metabolic syndrome (MetS) [57]. SIRT3 is believed to be germane to fertilization as well as embryo formation [30]. In vitro maturation (IVM) experiments of oocyte from mice and humans, supplementation of quercetin enhanced antioxidant action through decrease of SIRT3 expression leading to the greater rate of successful maturation [58]. In humans, knockout of SIRT3 considerably enhanced ROS formation in GCs [59]. Additionally, elimination of SIRT3 further results in reduction of expression of aromatase, 17βhydroxysteroid dehydrogenase1, steroid acute regulatory protein, cholesterol side chain cleavage,3 βhydroxysteroid dehydrogenase in GCs followed by reduction in progesteroneliberation, pointing to a part of SIRT3 in folliculo genesis as well asluteinization events [59]. Conversely, activating SIRT3 working might be aiding in the sustenance of human reproduction by sustenance of GC along with oocyte working [59]. Moreover, maternal obtained SIRT3 apparently confers protection in early embryo generation from stressful situations at the time of IVF along with culture in mice. SIRT3 is key for sustenance of mitochondrial homeostasis a part from avoidance of ROS-p53 pathway that is implicated in generational abnormalities [30]. In women presenting with advanced age the quantities of SIRT3 mRNA in GC in addition to CC reduces significantly in contrast to those in younger women [60]. In the case of ageing oocytes, a drastic reduction in SIRT1, SIRT2 as well as SIRT3 mRNA quantities correlated with enhanced intracellular ROS quantities were found. Additionally, delivering nicotinamide for hampering of SIRT1, SIRT2 as well as SIRT3 was documented to exaggerate postovulatory ageing in mice [61]. The hampered SIRT3 expression stimulated by escalated palmitic acid downregulated AMPK/ SIRT3 pathway stimulating palmitic acid ceramide accrual, mitochondrial protein hyperacetylation as well as impairment in porcine oocytes [62]. Role of SIRT4- SIRT7: The remaining members of sirtuin family inclusive of SIRT 4, SIRT5, SIRT 6, along with SIRT 7 have further been documented to abrogate oocyte quality. SIRT 4 has been acknowledged to be a deacetylase enzyme, possesses a crucial part in energy metabolism. Furthermore, it has been illustrated to be a tumor repressor in variable malignant situations inclusive of prostate cancer. It controls pancreatic β cells insulin liberation by glutamate dehydrogenase (GDH) or insulin breakdown enzymes [63]. Moreover SIRT3 in alliance with SIRT 4 functions for sustenance of mitochondrial NAD+- quantities subsequent to stress full conditions hampering apoptosis [64]. SIRT 4was illustrated to be a controller of energy metabolism in addition to meiotic equipment which possesses a key part regarding oocytes maturation. In expansion, SIRT 4 overexpression interferes with mitochondrial reorganization as well as stimulates meiotic aberrations at the time of oocytes maturation. In the mean while remarkably enhanced spindle/ chromosomal abnormalities on knockout of SIRT4 in oocytes of mice which were old [65]. In a different experiment knockout of SIRT4 possess the capacity of avoidance of ageing stimulated mitochondrial impairment in postovulatory phage [66]. Coenzyme Q10 supplementation was documented to decrease the mitochondrial impairment in aged oocytes by decreasing the strength of SIRT4 overexpression [66]. SIRT5 possesses an elemental part in sustenanceof energy homeostasis by desuccinylating metabolicc enzymes [4]. Noticeably, recent outcomes documented that repressionofSIRT5 action possessed the capacity of reduction of SARS COV2-viral quantities by crosstalk amongst associated non- structuralprotein Nsp14 [67]. Gene expression of SIRT5 in GC significantly reduced in GC in addition to CC in patients with diminished ovarian reserve (DOR) along with advanced maternal ageing in contrast to patients with normal reserve [61]. SIRT6 has been observed to aiding in the chromatin remodeling. DNA healing, gene transcription, OS as well as energy metabolism [68]. Mice possessing SIRT6 deficiencydisplay mitigation of NFκB signaling through H3K9 deacetylation at chromatin resulting in cellular senescence [69]. Furthermore, it is acknowledged to postpone chronic nephropathy propagation through its antioxidant as well as anti-inflammatory actions in mammals [68]. Porcine experiment illustrated that hampering of SIRT6 in cumulus free oocytes decreased the rate of 1st polar body expulsion [70]. Moreover, this repression hampered cumulus expansion in addition to gap junctional connection [86]. SIRT7 manipulates RNA polymerase 1 transcription which takes part in lipid metabolism as well as mitochondrial working [71]. Its particular knockout results in an upheavaled spindle/ chromosome in addition to elimination of cortical actin cap interfering with meiotic maturation, followed by formation of aneuploidy oocytes. Noticeably, in case of obese mice, reduction of SIRT7protein quantities takes place significantly, while overexpression of SIRT7 results in improvement of maternal obesity correlated meiotic aberrations in addition to OS in oocytes [72].
Treatment Implicating SIRT Signaling Pathway Regarding improvement of Oocyte Quality
Figure 4 illustrates the probable treatments implicating SIRT signaling pathways.
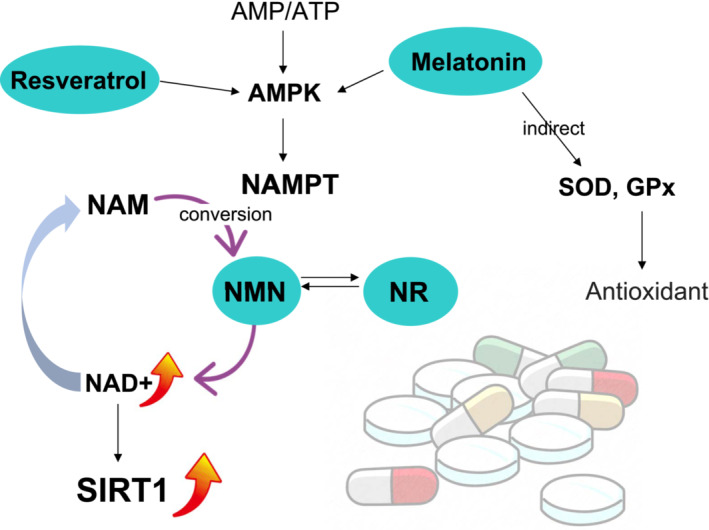
Figure 4: Courtesy ref no-31-Potential therapeutics involving the SIRT signaling pathway. Currently, supplementation of four different therapeutics, melatonin, resveratrol, nicotinamide mononucleotide (NMN), and nicotinamide riboside (NR) has demonstrated to restore oocyte quality through SIRT signaling pathways. The action of this therapeutics was integrated mainly in activation of SIRT1 induced by elevated NAD+. AMP, Adenosine monophosphate; AMPK, AMP‐activated protein kinase; ATP, adenosine triphosphate; GPx, glutathione peroxidase; NAD+, oxidative nicotinamide adenine dinucleotide; NAM, nicotinamide; NAMPT, nicotinamide phosphoribosyl transferase; SOD, super oxide dismutase.
Melatonin
Melatonin (alias N- acetyl 5-methoxy-tryptamine) represents an endogenous indole amine hormone basically generated by the pineal gland which modulates variable significant physiological processes [73]. Working in the form of an antioxidant, melatonin in addition to its products confers protection to cells from oxidation, inflammation as well as apoptosis [74]. Regarding reproductive health escalating outcomes pointed that it counters OS, apart from sustenance of mitochondrial working, avoided age- associated OS along with reproductive system conditions [73]. Its presence is there in all the stages of porcine oocytes, postpones ovarian ageing apart from improvement of oocyte maturation as well as embryo formation [75]. Melatonin escalates mitochondrial working as antioxidant by foraging free radicals which are toxic, confer protection tocell membranes apart from mitochondria, and postpones reduction of the human oocyte quality [76]. Experiments on animal models of variable species illustrated that melatonin promotes oocyte maturation by modulation of the production of luteinizing hormone (LH), cumuluscells expansion, oocyte maturation factors, DNA methylation along with histone acetylation [11]. Melatonin actions in mice is by generating antioxidant actions by direct activation of the sirtuin pathway, MT1/ AMPK pathway, or by direct escalation of the total antioxidant capacity (TAC)of antioxidant enzymes as well as superoxide dismutase (SOD) [77]. The ovarian SIRT1/ SIRT 3 mRNA expression quantities might be activated by melatonin [77]. Follicular GCs in addition to oocytes possessing the capacity of forming melatonin .Its inadequate quantities in follicular fluid is correlated with advanced maternal age correlated meiotic aberrations in mice [78]. In the meantime greater quantities of melatonin in case of murine follicular fluid fights the OS correlated with free oxygen radicals liberated at the time of ovulation [78]. Delivery of melaton in in ageing mice enhanced significantly the numbers apart from density of transzonal projections (TZP) in cumulus oocyte complexesas well as avoided TZP retracting in vitro, that are significant regarding sustenance of connection amongst GCs along with oocytes apart from promoting oocyte maturation [78]. Melatonin reverts the meiosis-deficiency phenotype through SIRT 1/ (SOD 2 that constitutes a main antioxidant enzyme in oocytes [79]. Greater work illustrated that melatonin decreased advanced maternal age correlated meiotic aberrations in ageing mice via the SIRT2 based H4K16 deacetylation pathway [80]. Additionally, melatonin confers protection to mitochondrial working as well as decreases OS injury in mice oocytes by reducing the quantities of 8 hydroxydeoxy guanosine (8OHDG) in mitochondria [81]. Moreover, melatonin escalates SIRT 3 in addition to SOD 2 action, hampering the autophagic demise of murine hepatocyte that confers protectiontoovarian cells along with decrease follicular atresia [82].
Furthermore, experiments on mice models illustrated that long term delivery of melatonin might decrease ovarian ageing pointed that significantly escalate numbers in addition to oocyte quality [81]. 1mm melatonin addition reduced ROS quantities, spindle aberrations as well as DNA degradation ratio in mice impacted by10- hydroxycamptothecin [74]. Akin to that in vivo supplementation of melatonin repressed mitochondrial impairment, reverting oocytes meiotic worsening in mice [83]. In a separate experiment it got illustrated that melatonin postponed postovulatory mouse oocyte ageing a SIRT1- MnSod based pathway pointing a molecular mode validate greater application of melatonin regarding treatment of infertility [84]. Delivery of oral melatonin in human sat the time of IVM significantly mitigated OS as well as meiotic aberrations in oocytes in obese females via SIRT 3 - SOD 2 based mode [85]. Clinical trials documented a considerable escalation in the numbers of in addition to mature oocytes attained subsequent to melatonin supplementation at the time of IVF cycle [86]. Furthermore, melatonin therapy under IVF-ET program was advocated to result in improvement of fertilization as well as pregnancy rates in case of women with lesser fertilization rates (≤50%) [87]. Nevertheless, as per a randomized controlled trial (RCT),conducted recently novariation in oocyte numbers of MII, fertilization rates, embryo quality, clinical pregnancy rates or LBR subsequent to melatonin supplementation was observed amongst treatment groupas well as control group [89]. However in a systematic review and meta-analysis melatonin therapy considerably escalated the numbers of oocytes retrieved, mature oocytes in addition to embryo quality, along with clinical pregnancy rates [90]. As per ameta-analysis inclusive of a RCT, oral melatonin supplementation at the time of IVF escalated the oocyte numbers, although not to a significant quantity [91]. Conversely, another study (non RCT), demonstrated that the percentage of mature oocytes (i.e MII stage oocytes /full oocytes obtained) was not altered subsequent to melatonin supplementation in addition to the biochemical pregnancy rates was not significantly greater in the group in receipt of melatonin [91]. Reiter, et al. further summarized the role of melatonin in rescuing age associated fertility [92] ((Figure 5 & 6) for mode of action of melatonin). Furthermore in view of remarkable reduction in melatonin in latter stages of life along with melatonin supplementation in latter part of life might postpone ageing in view of its greater quantities in follicular fluid in contrast to blood points to its correlation with generation in oocytes, GC’s in addition to luteal cells might aid in delaying oocyte deterioration and postpone and rescue fertility in women presenting later and further obviate the need for ART.
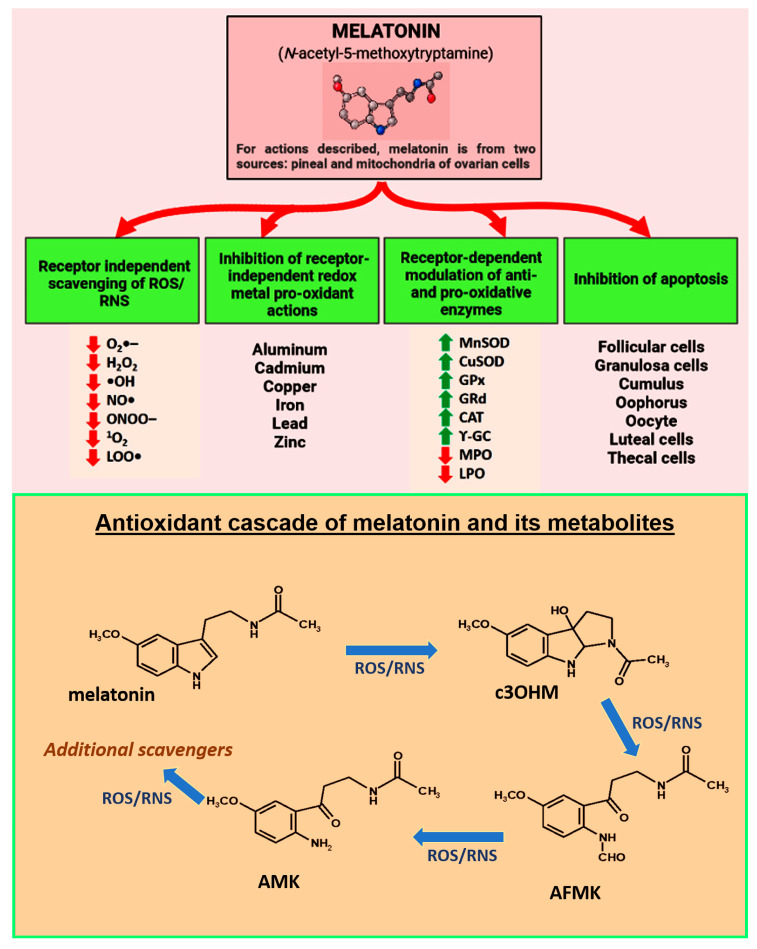
Figure 5: Courtesy ref no92-melatonin which is nocturnally generated by pineal gland and in a non-circadian way by mitochondria of other cells or constituents of ovary is posited to be a key factor in protecting against premature infertility and reproductive finishing. Furthermore publications pointed that melatonin represses or postpones ovarian ageing in animals and lowers the frequency of infertility in humans. Regarding the protective actions of melatonin against reproductive collapse this figure summarizes multiple receptor independent andreceptor dependent events which interfere with in particular Oxidative stress mediated ovarian deterioration and the key cells undergoing apoptosis along with directly foraging ROS/RNS and indirectly lowering Oxidative damage by upregulating antioxidant enzymes or along with down regulating prooxidant enzymes. Melatonin binds redox reactive transition metal ions to limit Heber Weiss and Fenton reaction a, thus decreasing the generation of v toxic hydroxyl (OH) radicals. Red arrow point’s inhibitor; green point’s stimulator. Bottom part represents what is acknowledged as melatonin ‘s antioxidant cascade as a radical forager .Thus not only is melatonin a direct radical forager but indirectly via its metabolites as well.-cyclic 3OHmol (3OHM);N-acetyl-Normyl 5 methoxy kynuramine (AFMK); N-acetyl-5 methoxy kynuramine (AMK); and probably others. Furthermore metabolites are more effective in contrast to melatonin itself regarding ROS/RNS.ROS/ RNS-Reactive oxygen species/ Reactive nitrogen species.
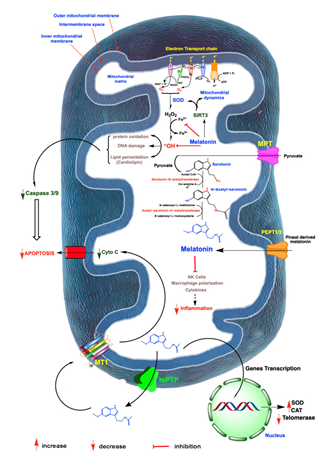
Figure 6: Courtesy ref no-92-A summary of free radicals which is generated in mitochondria and role of melatonin in Oxidative damage and ovarian ageing, radicals are generated partly to electron leakage from electron transport chain(ETC), in inner mitochondrial membraneand the rogue electron chemical reduces adjacent oxygen molecules to produce superoxide anion radical (O2-). This is quickly dismutated by superoxide dismutase (SOD), to hydrogen peroxide (H2O2) or it couples with nitric oxide (NO), to generated the markedly oxidizing peroxide anion (ONOD-)-not shown; H2O2 is converted to hydroxyl (OH) radicals via Heber Weiss reaction which is kinetically slow via Fenton reaction, both of which require a transition metal like ferrous (Fe2+), the OH and other oxidants damage molecules which initiate apoptosis. The antioxidant melatonin which is generated by a numbers of ovarian cells likely in mitochondria as well as pineal gland derived melatonin which enter these organelles and chelates iron and other redox reactive transition metals via act of SIRT3 melatonin furtherupregulatesSOD2 and impact mitochondrial dynamics in favour of remaining mitochondria. Finally melatonin indirectly neutralizes OH and ONOD-combined action. Melatonin acts as a robust protector of mitochondrial integrity and presents ideal cellular functions which delays ovarian ageing. Melatonin also works as an anti-inflammatory which in particular when chronic compromised mitochondrial physiology => ovarian cell including oocytes deterioration mitochondrial generated melatonin also escapes these organelles to a melatonin receptor (MT1), in the mitochondrial membrane which reduces the liberation of cytochrome c, thereby initiating programmed cell death (PCD) which could otherwise advance ovarian ageing. Lastly in ovarian cancer melatonin impacts generation of telomeres by decreasing telomerase activity to these strong cancer cell renewal I-IV mitochondrial complex.
Resveratrol
Resveratrol (alias(trans 3, 4, 5-trihydroxy stilbene) represents a polyphenolic substance acknowledged for possessing antioxidant, anti-inflammatory along with anti- ageing characteristic [32]. In view of its antioxidant as well as anti-apoptosis actions in mammals resveratrol has been illustrated to result in improvement of quality of ageing oocytes. Escalating outcomes have pointed resveratrol to confer protection to oocytes from age - based deficiencies in the form of a robust SIRT1stimulator [16, 32]. Resveratrol was observed to leadtoSIRT1activation indirectly through modulating the action of nicotinamide phosphoribosyl transferases along with AMPK [18, 93].
In case of mice resveratrol therapy for 1yr was documented to escalate SIRT1 mRNA quantities along with lead to improvement of numbers as well as oocyte quality in contrast to ageing mice which were age matched, as pointed by spindle morphology along with chromosomal alignment. Furthermore, resveratrol abrogated age correlated lesser embryo generation in a dose based fashion [94]. Moreover, the female fertility results were evaluation by Okamoto, et al. [95], subsequent to various time periods of resveratrol therapy in vivo by utilization of ageing mice models. Their outcomes illustrated that use in a v limited period (like 1 wk) of resveratrol therapy enhanced the quantities of Sirt1, Sirt3, Sirt4, Sirt5 as well as Sirt7 apart from improvement of mitochondrial working, however not copy numbers of mt DNA in ageing mice resulting in escalating implantation rates along with live birth rates (LBR) apart from reduction in miscarriage rates without leading to aberrationsin fetuses along with placenta. The greater serum resveratrol quantities lead to greater implantation rates apart from LBR [95]. These outcomes pointedthatresveratrol might be a probable anti- ageing treatment for women of advanced age.
In case of ageing bovines oocytes, resveratrol was documented to lead to improvement of oocyte quality by escalating mitochondrial production in addition to breakdown of oocytes as well as modulating genes in granulosa cells, whose expression quantities are correlated with the formation of oocyte as well as embryos [96]. In another akin experiment, maturation medium supplementation for oocytes obtained from ageing bovines oocytes resveratrol resulted in improvement of the fertilization ratio [49].
Akin observations were seen with porcine oocytes displaying that adding resveratrol therapy escalated SIRT1 activation along with leading to enhanced mitochondrial working at the time of in vitro ageing [97]. In agreement Polydatin addition (alias glyosidic resveratrol form) in case of in vitro maturation (IVM) media escalated SIRT1 protein as well as reduction of OS, leading to advantageous blastocyst formation in bovines [98]. This got validated by significant escalation of SIRT1-‘s gene expression in bovine GC, CC, oocytes as well as blastocysts apart from diminished ROS subsequent to resveratrol therapy [99]. GrzeczaA and Kordowistzki recently summarized how structurally resveratrol targets SIRT1 in oocytes in particular in women at latter stages of fertility &in other mammals [100] Figure 7 & 8.
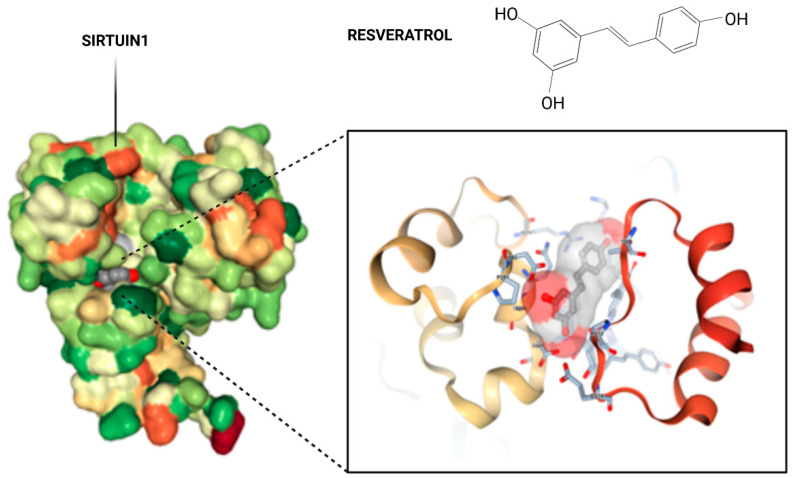
Figure 7: Courtesy ref no-100-Crystal structure of Sirt1 in complex with resveratrol. The overall structure of SIRT1 with the protein domains and the catalytical core of the protein in magnification in amino acid structure binding to resveratrol. A pair of SIRT1 residues involved in inter domain interactions are shown in a stick model, with hydrogen bonds.
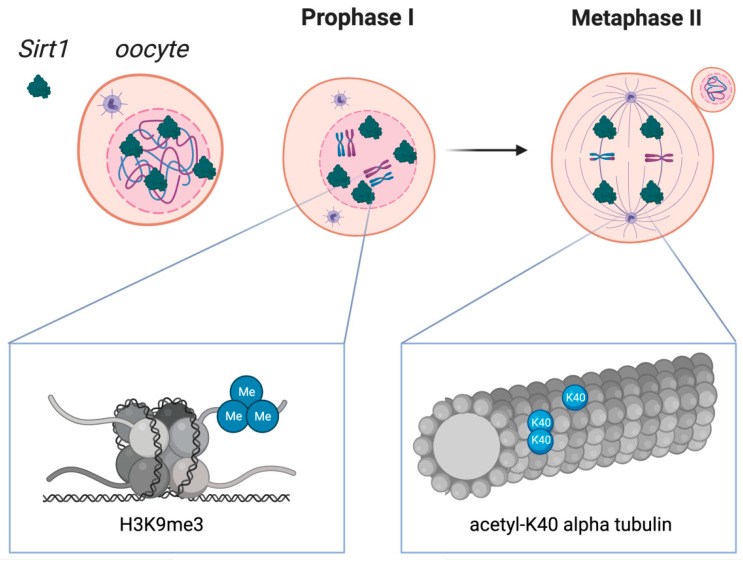
Figure 8: Courtesy ref no-100-Sirtuin 1 distribution in oocytes during crucial stages of the first and second meiotic diviScheme 1. Localization in oocytes at prophase I and at metaphase II. SIRT1 is mainly expressed in prophase I-arrested oocytes at nuclear localization and on the spindle in metaphase II oocytes. Molecular functions of Sirtuin 1 during meiotic divisions are shown at a chromatin level (left box) and at the spindle formation level (right box). Heterochromatin formation with possible impact on chromosome compaction, gene silencing, and DNA damage repair signaling have been reported upon SIRT1, which has shown to promote the H3K9me3 deposition, as depicted in the left zoomed-in box. As shown in the right zoomed-in box, SIRT1 also regulates the acetylation of α-tubulin, therefore influencing the formation of acetyl K-40 α-tubulin and suggesting an impact during spindle alignment.
Nicotinamide Mononucleotide (NMN) Along with Nicotinamide Riboside (NR)
Working in the form of NAD+ intermediate, NMN along with NR have been illustrated to mitigate age correlated conditions with effectiveness [101]. Utilization of NMN along with NR for therapy escalated NAD+ quantities followed by facilitation of SIRT1 activation in neurovascularunits [102, 103]. Additionally, molecular evaluation demonstrated that NMN actions of conferring neurovascular protection got modulated by stimulating genes implicated in rejuvenating mitochondria, pointing that such NAD+ intermediates might be utilized in mitochondrial targeted therapies [102]. NMN was illustrated to restore NAD+ quantities apart from replenishment of genes correlated with OS in numerous organs by activating SIRT1 in diet as well asage induced Diabetes in mice [103].
As far as oocyte quality is concerned, in vivo NMN supplementation was demonstrated to enhance oocyte ovulation apart from escalate meiotic competency along with fertilization capacity of ageing mice. Furthermore, transcriptome assessment demonstrated that NMN possessed the capacity of restoration of mitochondrial working along with deplete ROS accrual for apoptosis repression in oocytes of ageing mice [104]. In another skin experiment, conducted in ageing mice NMN had the capacity of abrogating the generational potential of oocytes modulated by a transgenicover expression of NAD+ - based deacetylases SIRT 2 [105]. NMN medium further resulted in enhancement of blastocyst generation in embryos obtained from oocytes from ageing women supplementation in the embryo culture [106]. Moreover, NMN therapy repressed the accrual of mit DNA mutations secondary to NADH/ NAD+ redox status in mice [107]. An akin outcome was documented in porcine oocytes regarding NMN supplementation corrected the meiotic abnormalities as well as mitochondrial working subsequent to exposure to ethylene glycol butyl ether through restoration of NAD+ - quantities as well as depletion of the enhanced ROS [108]. NR was observed to decrease quantities of ROS along with spindle abnormalities in ageing mouse oocytes. NR supplementation further enhanced ovarian mitochondrial energy metabolism, as well as reduced mitochondrial form clusters [109].
Gonzalez Fernandez, et al. [110], evaluated gene expression of sirtuins by quantitative Real Time- Polymerase Chain reaction (qRT-PCR), in human mural granulosa-lutein cells (hGL) from IVF patients in various infertility diagnostic groups as well as in oocyte donors (OD; control group). Study 1: sirtuins genes’ expression quantities along with association with age a part from IVF paradigms in women with no ovarian factor. They observed significantly greater expression quantities of SIRT1, SIRT2 and SIRT5 in patients ≥40 years old in contrast to OD as well asin women amongst 27 along with39 years old with tubal or male factor, apart from no ovarian factor (NOF). Only SIRT2, SIRT5 along with SIRT7 expression associated with age. Study 2: sirtuin genes’ expression in women poor responders (PR), endometriosis (EM) and polycystic ovarian syndrome. Compared to NOF controls, They observed greater SIRT2 gene expression in all diagnostic groups while SIRT3, SIRT5, SIRT6 as wellSIRT7 expression were greater only in PR. Associated with clinical paradigms SIRT1, SIRT6 as well as SIRT7 associate positively with FSH and LH doses delivered in EM patients. The number of mature oocytes retrieved in PR is positively correlated with the expression levels of SIRT3, SIRT4 and SIRT5. These outcomes pointed that cellular physiopathology in PR’s follicle may be correlated associated with cumulative DNA injury, pointing that further studies are warranted [110].
Conclusion
Escalating validation has been attained regarding sirtuins confer protection to the oocytes quality. Subsequent to basic apart from animal studies certain probable therapeutics implicating the SIRT signaling pathway have been seen to enhance oocyte quality in women with advanced age. Nevertheless, the biggest limitation currently is not adequate human randomized placebo controlled trials(RCT) are present to corroborate same animal studies in view of not all animal physiology corresponds to human physiology hence the skepticism if at all what apparently we observe as good results in certain humans who take such form of supplements without prescription as good results have to be substantiated by RCT’s. In any case melatonin has been seen to work in certain disease conditions where both resveratrol and melatonin work like metabolic diseases gives a ray of hope that akin SIRT targeting would prove to be of benefit in women who deliberately postpone fertility and might be in DOR as well. Despite, these therapies can be bought in view of their availability in the form of supplements as well asease of taking, easy availability without need for prescription, their utilization regarding infertility patients therapy are still contradictory. For assuming that these supplements as diets can be advocated as an ‘’add on therapy’’ for reproductive treatments requires well fashioned RCT’s before bringing them from bench to bedside.
References
-
Magnus MC, Wilcox AJ, Morken NH, Weinberg CR, Håberg SE, et al. (2019) Role of maternal age and risk of pregnancy history in risk of miscarriage: prospective register based study. BMJ 364: l869.
-
Musson R, Gąsior Ł, Bisogno S, Ptak GE (2022) DNA damage in preimplantation embryos and gametes: specification, Clinical relevance and repair strategies**.** Hum Reprod Update 28(3): 376-399.
-
Babayev E, Duncan FE (2022) Age associated changes in follicular fluid: the local oocyte microenvironment as a determinant of gamete quality. Biol Reprod 106(2): 351- 365.
-
Maissan P, Mooij EJ, Barberis M (2021) Sirtuins mediated system level **r**egulation of mammalian tissues at the interface between metabolism and cell cycle: a systematic review. Biology 10(3): 194.
-
Vazquez BN, Vaquero A, Schindler K (2020) Sirtuins in female meiosis and reproductive longevity. Mol Reprod Dev 87(12): 1175-1187.
-
Guo YM, Sun TC, Wang HP, Chen X (2021) Research progress of melatonin(s) in improving Ovarian function: a review of the current status. Aging 13(13): 17930- 17947.
-
Kochar Kaur K, Allahbadia GN, Singh M (2020) How does Epigenetics Regulate Development of Placenta and Placental Pathologies like Preeclampsia (PE), Intrauterine growth Restriction(IUGR)-With Main emphasis on PE. Advances in Bioengineering and Biomed Res.
-
Kochar Kaur K, Allahbadia GN, Singh M (2021) Potential role of Epigenetic Modulation in prevention or therapy for Diabetic Kidney Disease-still a dream or a reality –A Systematic Review. J Diab Nephro Diab Mgmt 1:1: 1-26.
-
Kochar Kaur K, Allahbadia GN, Singh M (2019) Will Utilization of Resveratrol’s Effects be Practical in Multiple Chronic Inflammatory Diseases and Autoimmune Diseases: A Detailed Review of its Immune Responses and Further Clinical Development in Humans in Future – A Systematic Review. Acta Scientific Microbiology Special Issue 1: 14-23.
-
Da Broi MG, Giorgi VSI, Wang F, Keefe DL, Albertini D, et al. (2018) Influence of follicular fluid and cumulus cells on oocyte quality: Clinical implications**.** J Assist Reprod Genet 35(5): 735-751.
-
Yong W, Ma H, Na M, Gao T, Zhang Y, et al. (2021) Roles of melatonin in the field of reproductive medicine. Biomed Pharmacother 144: 112001.
-
Rodríguez-Varela C, Herraiz S, Labarta E (2021) Mitochondrial enrichment in infertile patients: a review of different mitochondrial replacement therapies. Ther Adv Reprod Health 15: 26334941211023544.
-
Pasquariello R, Ermisch AF, Silva E, McCormick S, Logsdon D, et al. (2019) Alterations in oocytes mitochondrial numbers and functions are related to spindle defects and occur with maternal ageing in in mice and humans. Biol Reprod 100(4): 971-981.
-
Almansa-Ordonez A, Bellido R, Vassena R, Barragan M, Zambelli F, et al. (2020) Oxidative stress in reproduction :a mitochondrial perspective. Biology 9(9): 269.
-
Buffenstein R, Edrey YH, Yang T, Mele J (2008) The Oxidative stress theory of ageing: embattled or invincible? Insights from nontraditional model organisms. Age 30(2-3): 99-109.
-
Chung S, Yao H, Caito S, Hwang JW, Arunachalam G, et al. (2010) Regulation of SIRT1 in cellular functions**:** Roles of polyphenols. Arch Biochem Biophys 501(1): 79-90.
-
Konstantinidis M, Alfarawati S, Hurd D, Paolucci M, Shovelton J, et al. (2014) Simultaneous assessment of aneuploidy, polymorphisms and mitochondrial DNA in human polar bodies and embryos with the use of a novel microarray platforms. Fertil Steril 102(5): 1385-1392.
-
Ishii T, Miyazawa M, Takanashi Y, Tanigawa M, Yasuda K, et al. (2014) Genetically induced Oxidative stress in mice causes thrombocytosis, splenomegaly, and placental angiodysplasia that leads to recurrent abortion. Redox Biol 2: 679-685.
-
Quintero RA, Ishii K, Chmait RH, Bornick PW, Allen MH, et al. (2007) Sequential selective laser photocoagulation of communicating vessels in twin- twin transfusion syndrome. J Matern Fetal Neonatal Med 20(10): 763- 768.
-
de Bruin JP, Dorland M, Spek ER, Posthuma G, van Haaften M, et al. (2004) Age related changes in the ultrastructure of resting follicle pool in human ovary. Biol Reprod 70(2): 419-424.
-
Lim J, Luderer U (2011) Oxidative damage increases and antioxidant gene expression decreases with ageing in the mouse ovary. Biol Reprod 84(4): 775-782.
-
Tatone C, Amicarelli F, Carbone MC, Monteleone P, Caserta D, et al. (2008) cellular and molecular aspects of ovarian follicle ageing. Hum Reprod Update 14(2): 131- 142.
-
Gilchrist RB, Lane M, Thompson JG (2008) Oocyte secreted: regulators of cumulus cell function and oocyte quality. Hum Reprod Update 14(2): 159-177.
-
Cheng EH, Chen SU, Lee TH, Pai YP, Huang LS, et al. (2013) Evaluation of telomere length in cumulus cells as a potential biomarkers oocyte and embryo quality. Hum Reprod 28(4): 929-936.
-
Grabowska W, Sikora E, Bielak-Zmijewska A (2017) Sirtuins, a promising target in slowing down the ageing process. Biogerontology 18(4): 447-476.
-
Ong ALC, Ramasamy TS (2018) Role of Sirtuin- 1-p53 regulatory axis in ageing, cancer and cellular reprogramming. Ageing Res Rev 43: 64-80.
-
Lagunas-Rangel FA (2019) Current role of mammalian Sirtuins in DNA repair. DNA Repair 80: 85-92.
-
Singh CK, Chhabra G, Ndiaye MA, Garcia-Peterson LM, Mack NJ, et al. (2018) The Role of Sirtuins in antioxidant and redox signaling. Antioxid Redox Signal 28(8): 643- 661.
-
Finkel T, Deng CX, Mostoslavsky R (2009) Recent progress in the biology and physiology of Sirtuins. Nature 460(7255): 587-591.
-
Tatone C, Di Emidio G, Vitti M, Di Carlo M, Santini S Jr, et al. (2015) Sirtuin functions in female fertility: possible role in Oxidative stress and ageing. Oxid Med Cell Longev 2015: 659687.
-
Vo KCT, Sato Y, Kawamura K (2023) Improvement of oocyte quality the SIRT signaling. Reprod Med Biol 22(1): e12510.
-
Pasquariello R, Verdile N, Brevini TAL, Gandolfi F, Boiti C, et al. (2020) The Role of resveratrol, in mammalian reproduction. Molecules 25(19): 4554.
-
Morris BJ (2013) Seven Sirtuins for Seven deadly diseases of ageing**.** Free Radic Biol Med 56: 133-171.
-
Du J, Zhou Y, Su X, Yu JJ, Khan S, et al. (2011) Sirt5 is a NAD dependent protein lysine demalonylase and desuccinylase. Science 334(6057): 806-809.
-
Michan S, Sinclair D (2007) Sirtuins in mammals: insights into their biological functions. Biochem J 404(1): 1-13.
-
Price NL, Gomes AP, Ling AJ, Duarte FV, Martin-Montalvo A, et al. (2012) SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab 15(5): 675-690.
-
Yuan Z, Zhang X, Sengupta N, Lane WS, Seto E, et al. (2007) SIRT1 regulates the function of the Nijmegan breakage protein. Mol Cell 27(1): 149-162.
-
Wang RH, Sengupta K, Li C, Kim HS, Cao L, et al. (2007) Impaired DNA damage response, genomic instability and tumorigenesis in SIRT1 mutant mice. Cancer Cell 14(4): 312-323.
-
Oberdoerffer P, Michan S, McVay M, Mostoslavsky R, Vann J, et al. (2008) SIRT1 redistribution on chromatin promotes genomic stability but alters gene expression during ageing. Cell 135(5): 907-918.
-
Deng CX (2009) SIRT1 is it a tumor promoter or suppressor? Int J Biol Sci 5(2): 147-152.
-
Nemoto S, Fergusson MM, Finkel T (2005) SIRT1 functionally interacts with the metabolic regulator and transcriptional coactivator PGC-1 **α.** J Biol Chem 280(16): 16456-16460.
-
Di Emidio G, Falone S, Vitti M, D’Alessandro AM, Vento M, et al. (2006) SIRT1 signaling protects mouse oocyte against Oxidative stress and is deregulated during ageing. Hum Reprod 29(9): 2006-2017.
-
Xing X, Zhang J, Wu T, Zhang J, Wang Y, et al. (2021) SIRT1 reduces epigenetic and non-epigenetic changes to maintain the quality of postovulatory aged oocytes in mice. Exp Cell Res 399(2): 112421.
-
Lee D, Goldberg AL (2013) SIRT1 protein by blocking the activity of transcription factor FoxO1 and FoxO3, inhibits muscle atrophy and promotes muscle growth. J Biol Chem 288(42): 30515-30526.
-
Broussy S, Laaroussi H, Vidal M (2020) Biochemical mechanism and biological effects of silent information regulator 1(SIRT1) by EX-527(SEN00014196or selisistat). J Enzyme Inhib Med Chem 35(1): 1124-1136.
-
Di Emidio G, Tatone C, Barbato V, Genovese V, Placidi M, et al. Protective effects of a SIRT1 inhibitor on primordial follicles activation and growth induced by cyclophosphamide: insights from a bovine in vitro folliculogenesis system. J Assist Reprod Genet 39(4): 933-943.
-
Takeo S, Sato D, Kimura K, Monji Y, Kuwayama T, et al. (2014) Resveratrol improves the mitochondrial function and fertilization outcomes of bovine oocytes. J Reprod Dev 60(2): 92-99.
-
Zhang XM, Li L, Xu JJ, Wang N, Liu WJ, et al. (2013) Rapamycin preserves the follicle pool reserve and prolongs ovarian lifespan of female rats via modulating mTOR activation and sirtuin expression. Gene 523(1): 82-87.
-
Takeo S, Kawahara-Miki R, Goto H, Cao F, Kimura K, et al. (2013) Age associated changes in gene expression and developmental competence of bovines oocytes and a possible countermeasure against age associated events. Mol Reprod Dev 80(7): 508-521.
-
Wu X, Hu F, Zeng J, Han L, Qiu D, et al. (2019) NMNAT2 mediated NAD+ generation is essential for quality of aged oocytes. Aging Cell 18(3): e12955.
-
Iljas JD, Wei Z, Homer HA (2020) Sirt 1 sustains female fertility by slowing age - related decline in oocyte quality required for post fertilization. embryo development Ageing Cell 19(9): 13204.
-
Fourcade S, Morato L, Parameswaran J**,** Ruiz M, Ruiz Cortes T, et al. (2017) Loss of Sirt 2 leads toaxonal degeneration and locomotordisability associated with redox and energy imbalance. Ageing Cell 16(6): 1404- 1413.
-
North BJ, Verdin E Sirtuins (2004) Sir2 related NAD dependent protein deacetylases. Genome Biol 5(5): 224.
-
Zhang L, Hou X, Ma R, Molley K, Schedl T, et al. (2014) Sirt 2 functions in spindle organization and chromosomal alignment in mouse oocyt**e** meiosis. FASEB J 28(3): 1435-1445.
-
Qiu D, Hou X, Han L, Li X, Ge J, et al. (2018) Sirt2 BubR1 acetylation mediates the effects of advanced maternal age on oocyte quality. Ageing Cell 17(1): 12698
-
Liu J, Liu M, Ye X, Liu K, HuangJ J, et al. (2012) Delay in oocyte ageing in mice by the antioxidant N-acetyl Lcysteine NAC. Hum Reprod 27(5): 1411-1420.
-
Hirschey MD, Shimazu T, Jing E, Grueter CA, Collins AM, et al. (2011) SIRT3 deficiency and mitochondrial protein hyperacetylation accelerates the development of metabolic syndrome. Mol Cell 44(2): 177-90.
-
Cao Y, Zhao H, Wang Z, Zhang C, Bian Y, et al. (2020) Quercetin promotes in vitro maturation of oocytes from humans and aged mice. Cell Death Dis 11(11): 965.
-
Fu H, Wada Hiraike O, Hirano M, Kawamura Y, Sakurabashi A, et al. (2014) SIRT3 regulates the expression of folliculogenesis and luteinization related genes and progesterone secretion by manipulating Oxidative stress in human luteinized granulosa cells. Endocrinology 155(8): 3079-3087.
-
Pacella Ince L, Zander-Fox DL, Lan M (2014) Mitochondrial SIRT3 and its glutamate dehydrogenase GDH are altered infollicular cells of women with reduced ovarian reserveor advanced maternal age. Hum Reprod 29(7): 1490-1499.
-
Zhang T, Zhou Y, Li L, Wang HH, Ma XS, et al. (2016) SIRT 123 protect mouse oocytes from postovulatory ageing. Ageing Albany NY 8(4): 685-696.
-
Itami N, Shirasuna K, Kuwayama T, Iwata H (2018) Palmitic acid induces ceramide accumulation ,mitochondrial protein hyperacetylation and mitochondrial dysfunction in porcine oocytes**.** Biol Reprod 98(5): 644-653.
-
Ahuja N, Schwer B, Carrobio S, Waltregny D, North BJ, et al. (2007) Regulation of insulin secretion by SIRT 4 a mitochondrial ADP ribosyl transferase. J Biol Chem 282(46): 33583-33592.
-
Yang H, Yang T, Baur JA, Perez E, Matsui T, et al. (2007) Nutrient sensitive mitochondrial NAD levels dictate cell survival. Cell 130(6): 1095-107.
-
Zeng J, Jiang M, Wu X, Diao F, Qiu D, et al. (2018) SIRT 4 is essential for metabolic structure and meiotic mouse oocytes maturation. Ageing Cell 17(4): 12789.
-
Xing X, Zhang J, Zhang J, Wang Y, Wang J, et al. (2022) Coenzyme Q10 supplement rescues postovulatory oocyte ageing by regulating SIRT 4 expression. Curr Mol Pharmacol 15(1):190-203.
-
Walter M, Chen IP, Vallejo Gracia A, Kim IJ, Bielska O, et al. (2022) SIRT5 is a pro viral factor that interacts with SARS COV2 Nsp14 protein. BioRxiv.
-
Yang X, Feng J, Liang W, Zhu Z, Chen Z, et al. (2021) Role of SIRT6 in Kidney Disease a novel therapeutic target. Cell Mol Life Sci 79(1): 53.
-
Kawahara TL, Michishita E, Andler S, Damian M, Barber E, et al. (2009) SIRT6 links histones H3 to Lysine 9deacetylation to NF kappa B dependent gene expression and organismal lifespan. Cell 136(1): 62-74.
-
Cao Z, Zhang D, Tang X, Wang Y, Qi X, et al. (2020) Cumulus cell derived and maternal SIRT6 differentially regulates porcine oocytes meiotic maturation. Theriogenology 142: 158-68**.**
-
Yan WW, Liang YL, Zhang QS, Wang D, Lei MZ, et al. (2018) Arginine methylation of SIRT7 couples glucose sensing with mitochondrial biogenesis. EMBO Rep 19(12): 46377.
-
Gao J, Huang Y, Li M, Zhao H, Zhao Y, et al. (2015) Effect of local Fibroblast growth factor and vascular endothelial growth factor on subcutaneously allotransplanted ovarian tissue in ovariectomized mice. PLoS One 10(7): 0134035.
-
Jiang Y, Shi H, Liu Y, Zhao S, Zhao H (2021) Application of melatonin in female reproduction in the context of Oxidative stress. Oxid Med Cell Longev 2021.
-
Wang L, Zhang J, Zhao C, Jia Z, Feng X (2021) Melatonin reverses 10 hydroxycamptothecin induced **a**poptosis and autophagy in mouse oocytes. Reprod Sci 28(7): 1839-1849.
-
Kim EH, Ridio MR, Lee BC, Kim GA (2020) Melatonin Nrf2 signaling activates Peroxisomal activities in porcine oocytes cumulus oocyte complexes. Antioxidants Basel 9(11): 1080.
-
Sun TC, Liu XC, Yang SH, Song LL, Zhou SJ, et al. (2020) Melatonin inhibits Oxidative stress and apoptosis in cryopreserved ovarian tissue via Nrf2/HO-1 signaling pathway. Front Mol Biosci 29(7): 163.
-
Zhang L, Zhang Z, Wang J, Lv D, Zhu T, et al. (2019) Melatonin regulates the activities of ovary and delays the fertility decline in female via MT1 AMPK pathway. J Pineal Res 66(3): 12550.
-
Zhang H, Li C, Wen D, Li R, Lu S, et al. (2021) Melatonin improves the quality of maternally aged oocytes by maintaining intercellular communication and antioxidant metabolite supply. Redox Biol 49: 102215. **79.** Zhang M, Lu Y, Chen Y, Zhang Y, Xiong B (2020) Insufficiency of melatonin in follicular fluid is a reversible cause for advanced maternal age related aneuploidy in oocytes. Redox Biol 28: 101327.
-
Li C, He X, Huang Z, Han L, Wu X, et al. (2020) Melatonin ameliorates the advanced maternal age associated meiotic defects in oocytes through the SIRT2 dependent H4K16 deacetylation pathway. Ageing Albany NY 12(2): 1610-1623.
-
Song C, Peng W, Yin S, Zhao J, Beibei FU, et al. (2016) Melatonin improves age induced fertility decline and attenuates ovarian mitochondrial Oxidative stress in mice. Sci Rep 6: 35165.
-
Pi H, Xu S, Reiter RJ, Gou P, Zhang L, et al. (2015) SIRT 3 SOD 2ROS dependent autophagy in cadmium induced hepatotoxicity and its salvage by melatonin. Autophagy 11(7): 1037-1051.
-
Xing CH, Wang Y, Liu JC, Pan ZN, Zhang HL, et al. (2021) Melatonin reverses mitochondrial dysfunctionand Oxidative stress induced apoptosis of Sudan1 exposed mouse oocytes**.** Ecotoxicol Environ Saf 225: 112783.
-
Yang Q, Dai S, Luo X, Zh J, Li F, et al. (2018) Melatonin postovulatory oocyte dysfunction by regulating SIRT14 expression. Reproduction 156(1): 81-92.
-
Han L, Wang H, Li L, Li X, Ge J, et al. (2017) Melatonin protects against maternal obesity associated ion Oxidative stress and meiotic defects in oocytes via SIRT 3 SOD 2 dependent pathway. J Pineal Res 63(3).
-
Batioglu AS, Sahin U, Gurlek B, Ozturk N, Unsal E (2012) The efficacy of melatonin administration on oocyte quality. Gynaecol Endocrinol 28(2): 91-93.
-
Tamura H, Takasaki A, Miwa I, Taniguchi K, Maekawa R, et al. (2008) Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radicals damage and improves fertilization rate. J Pineal Res 44(3): 280- 287.
-
Fernando S, Wallace EM, Vollenhoven B, Lolatgis N, Hope N, et al. (2018) Melatonin in assisted reproductive technology a pilot double blind randomized placebo controlled trial. Front Endocrinol Lausanne 19(9): 545.
-
Hu KL, Ye X, Wang S, Zhang D (2020) Melatonin application in assisted reproductive technology a systematic review and meta analysis of randomized controlled trials. Front Endocrinol Lausanne 11(160).
-
Mejlhede MAB, Jepsen JB, Knudson UB (2021) Oral melatonin supplementation during invitro fertilization treatment a systematic PRISMA review and meta analysis of randomized controlled trials. Gynaecol Endocrinol 37(12): 1079-85.
-
Hosseini FS, Shamsipour M, Yazdekhasti H, Akbari Asbagh F, Shahraki Z, et al. (2021) The effect of oral melatonin supplementation on MT ATP6 gene expression and IVF outcomes in Iranian infertile couples:a non randomized controlled trial. Naunyn Schmiedebergs Arch Pharmacol 394(7): 1487-1495.
-
Reiter R, Sharma R, Romero A, Manucha W, Xian Tan D, et al. (2023) Age related ovarian failure and fertility melatonin to the recue. Antioxidants Basel 12(3): 695.
-
Um JH, Park SJ, Kang S, Yang S, Foretz M, et al. (2010) AMP activated protein kinase deficient mice are resistant to themetabolic effects of resveratrol. Diabetes 59(3): 554-563.
-
Liu M, Yin Y, Ye X, Zeng M, Zhao Q, et al. (2013) Resveratrol protects against age associated infertility in mice. Hum Reprod 28(3): 707-717.
-
Okamoto N, Sato Y, Kawagoe Y, Shimizu T, Kawamura K (2022) Short term resveratrol treatmentrestored the quality of oocyte in ageing mice. Ageing Albany NY 14(14): 5628-5640.
-
Sugiyama M, Kawahara Miki R, Kawana T, Shirasuna K, Kuwayama T, et al. (2015) Resveratrol induced mitochondrial synthesis andautophagy in oocytes derived from early antral follicles of aged cows. J Reprod Dev 61(4): 251-259.
-
Ma R, Zhang Y, Zhang L, Han J, Rui R (2015) Sirt1 protects pig oocyte against invitro ageing. Anim Sci J 86(9): 826- 832.
-
Khan I, Kim EH, Lee BC, Song SH, Maslam A, et al. (2017) Polydatin improves the developmental competence of bovines oocytes invitro via induction of Sirtuin1 Sirt1. Reprod Fertil Dev 29(10): 2011-2020.
-
Wang F, Tian X, Zhang L, He CJ, Ji PY, et al. (2014) Beneficial effects of resveratrol on bovines oocytes maturation and subsequent embryo development after invitro fertilization. Fertil Steril 101(2): 577-586 .
-
Grzecza A, Kordowistzki P (2022) Resveratrol and SIRT1 anti ageing cornerstone for oocytes. Nutrients 14(23): 5101.
-
Yoshino J, Baur JA, Imai S (2018) NAD+ Intermediates: The Biology and Therapeutic Potential of NMN and NR. Cell Metab 27(3): 513-528.
-
Kiss T, Nyul Toth A, Balasubramaniam P, Tarantini S, Ahire C, et al. (2020) Nicotinamide mononucleotide NAM supplementation promotes neurovascular rejevunation in aged mice transcriptional foot prints of SIRT1 activation mitochondrial protection, anti inflammatory and along with anti-apoptotic effects. Gero Science 42(2): 527-546.
-
Chen X, Amorim JA, Moustafa GA, Lee JJ, Yu Z, et al. (2020) Neuroprotective effects and mechanism of action of Nicotinamide mononucleotide NAM in a photoreceptor degenerative model of retinal detachment. Ageing Albany NY 12(24): 24504-24521.
-
Yoshino J, Mills KF, Yoon MJ, Imai S (2011) Nicotinamide mononucleotide,a key NAD intermediate treats the pathophysiology of diet andage induced Diabetes in mice. Cell Metab 14(14): 528-536.
-
Miao Y, Cui Z, Gao Q, Rui R, Xiong B (2020) Nicotinamide mononucleotide supplementation reverses the declining quality of maternally aged oocytes. Cell Rep 32(5): 107987.
-
Bertoldo MJ, Listijono DR, Ho WJ, Riepsamen AH, Goss DM, et al. (2020) NAD repletion rescues female fertility during reproductive ageing. Cell Rep 30(6): 1670-1681.
-
Yang L, Lin X, Tang H, Fan Y, Zeng S, et al. (2020) Mitochondrial DNA mutation exacerbates female reproductive ageing via impairment of NADH NAD. Ageing Cell 19(9): 13206.
-
Miao Y, Li X, Shi X, Gao Q, Chen J, et al. (2021) Nicotinamide mononucleotide restores the meiotic competency of porcine oocytesexposed to ethylene glycol butyl ether. Front Cell Dev Biol 9: 628580.
-
Yang Q, Cang L, Wang Y, Luo X, Li H, et al. (2020) Increasing ovarian NAD levels improve mitochondrial functions and reverse ovarian aging. Free Radical Biol Med 156: 1-10.
-
Gonzalez Fernandez R, Martin Ramirez R, Rotoli D, Hernandez J, Naftolin F, et al. (2020) Granulosalutein cells Sirtuins gene expression profile differ between normal donors and infertilewomen. Int J Mol Sci 21(1): 295.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’