An Update on Primary Adult Hypogonadism in Elderly Males Secondary to Inflammaging: A Narrative Review
With the escalation of life expectancy, it is obvious that greater prevalence of conditions like late-onset hypogonadism (LOH) are bound to be encountered in an escalating manner in elderly males. LOH the properties of reduction in testosterone quantities in addition to possess significant influence in physical along with mental health. Whereas, the etiological factor of LOH continue to uncharted, an escalating interest is existent in the evaluation of part of inflammageing in its production. Inflammageing portrays theposit of chronic low grade systemic inflammation which takes place at the time of ageing. Such inflammatory status has been held responsible in the generation of variable age associated diseases. Separate cellular along with molecular mechanistic modes have been isolated to aid in inflammageing inclusive of cellular senescence, mitochondrial impairment, autophagy abnormalities in addition to immune senescence alias immunosenescence. Having reviewed earlier the part of inflammageing in addition to immunosenescence in ovarian ageing & variable infectious diseases we provide a a narrative review in reference to LOH. Further we deal with the, recent advance in its therapies inclusive of adrenomedullin, SS-31,amitochondria targeting peptide Leydig cells transplantation with use of CRISPR/Cas or use of variable stem cells.
Abbreviations
LOH: Late Onset Hypogonadism; LH: Luteinizing Hormone; ROS: Reactive Oxygen Species; OS: Oxidative Stress; CRP: C Reactive Protein; MAPK: Mitogen Activated Protein Kinase; SASP: Senescence- Associated Secretory Phenotype.
Introduction
Ageing portrays a complicated biological event which possesses the capacity of physiological as well as biochemical alteration in the human body. Regarding males, a significant result of ageing is the decrease in the generation of testosterone (T), a crucial hormone which possesses variable physiological functionality [1]. The basic region of T generation is the Leydig cells located in the testicular interstitial tissue. Nevertheless, such cells are functionally in addition to structurally changed as ageing takes place that result in diminished T molecules generation [2]. Such diminished T quantities are parallel with a disorder known as whose labelling has been as late-onset hypogonadism (LOH) that presents with a variety of symptoms inclusive of i)reduced libido ,ii)fatigue ,iii)sarcopenia as well as iv)mood variations [3]. Studies suggest that luteinizing hormone (LH) quantities continue to be germanely stable in elderly males whereas T liberation considerably gets diminished [4]. Contrasting evaluation of Leydig cell populations of young from elderly males displayed significantly diminished number of Leydig cells in the older subjects [5]. Furthermore, a disturbed redox harmony is observed in ageing Leydig cells that results in accrual of reactive oxygen species (ROS) which is followed by generation of Oxidative stress (OS). OS disturbs signaling pathways critical for T generation leading to diminished expression of crucial T generation molecules along with hampering the Leydig cells working [6].
Simultaneously, the ageing event results in chronic low grade inflammation termed inflammageing [7]. C Reactive Protein (CRP), tumor necrosis factor alpha(TNF-α), Interleukin (IL-6),IL-1β, as well as other correlated biomarkers utility is performed in a routine fashion in the form of serum biomarkers for evaluating inflammageing [8]. Other strategies for evaluating systemic inflammation are determination of circulating miRNA sequencing in addition to quantification of circulating mitochondrial DNA copy [9].
Apart from acute inflammation continuation of inflammageing takes place for longer duration in addition to is intricately correlated with variable ageing associated disorders [8]. A plethora of studies illustrated, escalation of quantities of proinflammatory factors for instance TNF-α, as well as IL-1β, in the circulation of older subjects [10]. The activation of inflammatory signaling pathway are pronouncedly regulated by nuclear factor κB(NFκB) along with p38 mitogen activated protein kinase (MAPK),which possess a central part in cellular ageing. The activation of such pathway stimulates the liberation of inflammatory mediators as well as proteasis, which terminates in the generation of senescence- associated secretory phenotype (SASP) [11]. In view of absence of blood test is barrier which confers protection, such proinflammatory cytokines might gain entry into testicular interstitial tissue via circulation, inimically impacting Leydig cell working [12]. Furthermore, the ageing event might stimulate the switching of testicular M2 macrophages in to M1 kind macrophages which stimulates the proinflammatory cytokines that might aggravate testicular inflammation [13]. Earlier we had reviewed growth in pregnancy besides how resveratrol could be used in numerous chronic inflammatory diseases and autoimmune diseases, role of NLRP3 inflammasome, Telomeres Dynamics in Reproduction and the significance of SIRT signaling pathway with NAD metabolism regarding improving oocyte quality and detailed NAD metabolism regarding ageing, obesity and tackling age correlated diseases by buttressing NAD+ for tackling ovarian ageing related to infertility and on the part of how immunosenescence and inflammageing participate in escalating generation of viral and bacterial diseases or pathological ageing with poor response to challenges as confronted during the COVID pandemic [14, 15, 16, 17, 18, 19, 20]. Here we provide an exhaustive narrative review with emphasis on recent update on how immunosenescence and inflammageing result in hypogonadism, mitochondrial impairment, pathophysiological propagation of late-onset hypogonadism (LOH) in elderly males.
Therapeutic interventions for inflammageing with idea of decreasing rate /halting propagation of age associated LOH is further detailed.
Methods
Here we conducted a narrative review utilizing search engine pubmed, google scholar ;web of science ;Embase; Cochrane review library utilizing the MeSH terms like immunosenescence; inflammageing; mitochondrial impairment; late-onset hypogonadism (LOH); therapeutic interventions for inflammageing from 1990’s till date in 2024 November .
Results
We found a total of 600 articles out of which we selected 125 articles for this review. No meta-analysis was done.
Controlling of Testosterone Generation by Leydig Cells
The testosterone (T) generation by Leydig cells is basically controlled by hypothalamic-pituitary- gonadal (H-P-G) axis [21]. Liberation of gonadotropin releasing hormone (GnRH) from the hypothalamus takes place in a pulsatile fashion [22]. GnRH cross talks with pituitary gonadotroph receptors, which stimulates the liberation of LH from the pituitary [23]. Binding of LH with Leydig cell receptors takes place which results in activation of adenylyl cyclase which gets followed by transformation of adenosine triphosphate (ATP)to cyclic adenosine monophosphate(AMP) with subsequent activation of downstream protein kinase A [24]. Subsequently cholesterol gains entry in to Leydig cells, where transportation of free cholesterol to mitochondria is attained by enzyme steroidogenic acute regulatory protein (StAR).
Mitochondrial P450 scc enzyme performs cleavage of free cholesterol to pregnenolone [25]. Pregnenolone shifts to endoplasmic reticulum (ER) which is transformed to progesterone (P) along with dehydroepiandrosteronesulphate (DHEAS), through enzyme 3β hydroxysteroid dehydrogenase(3β HSD) 2 along with cytochrome P450 17A1(CYP17A1) [26]. Following that P as well as DHEAS get converted to androstenedione. Finally T generation takes place via getting catalyzed by the enzyme 17 βhydroxysteroid dehydrogenase (17β HSD) [27]. At the time of ageing event alterations in the body might influence T generation by impacting the appropriate working of H-P-G) axis.
Immunesenescence
Immunesenescence, portrays the gradual reduction in the working of immune system with escalation of age [28]. It implicates diminished immune cells enrichment, jeopardized effectiveness, changed reaction design, escalated inflammation in addition to decreased tolerance [29]. The ageing event influencing peripheral blood immune cells, that possess the capacity of stimulating systemic inflammation, plausibly disturbing the working of H-P-G )axis leading to dysfunctional controlling of T generation . Additionally, ageing testicular macrophages might further aid in testicular inflammation, which directly hampers the working of Leydig cells.
Alterations in Peripheral Blood Immune Cells
Ageing immune cells aid in generation of systemic inflammation as well as age associated conditions [30]. Single cell sequencing study performed by Huang Z, et al. [30], in reference to human immune profiling over ageing time period along with gender illustrated considerable switching in peripheral blood immune cells that aided in systemic inflammation [31].
T Cells Changes in Ageing
The maximum significant changes observed in ageing males have been observed in T cells which possess the properties of considerable diminished quantities, significant characteristics of cellular ageing in addition to up regulation of inflammatory gene expression.
First single cell sequencing studies point to a significant decrease in CD8+T cells along with CD4+naïve T cells in peripheral blood of ageing males [31]. Ii) Martinez Zamudio et al. [31], observed senescent characteristics in CD8+T cells inclusive of a)senescence- associated β- galactosidase (sa β-gal) actions, b))telomere impairment as well as c) dysfunctional mitochondrial transcription factors in aged males [32]. iii) Furthermore, aged males display) escalated expression of particular genes for instance dual specific phosphatase -8(DUSP2), C-X-C motif chemokine ligand (CXCL4),DNA damage inducible transcript 4(DDIT4),NF- KBIA(encodes NF-κB1 transcription factor TF]-involved in immune &antiviral action)along with JUNB(JunB, protooncogene AP1 TF) in CD8+T cells along with CD4+ T cellsb) in addition to activation of MAPK signaling pathways [31]. Such alterations result in dysfunctional working of T cells, leading to diminished proliferation, escalated lysosomes quantities in addition to escalated proinflammatory cytokines [33].
Natural Killer Cells Dynamics in Elderly Males
Natural killer cells (NK cells), portray critical constituents in the context of innate immunity, which express CD 56 along with CD16 [34]. Dependent on CD 56 expression, their classification gets done as CD 56bright/CD16- phenotype, as well as CD 56dim/ CD16+ cells subpopulations. CD 56bright/CD16- NK cells have the capacity of liberating cytokines in addition to controlling immunity, whereas CD 56dim/ CD16+ cells display greater cytotoxicity along with implicated in innate immunity [35]. Ageing’s impact on such NK cells subpopulations in peripheral blood is a reduction in CD 56bright/CD16- subsets, with escalation of absolute number as well as percentage of CD 56dim/ CD16+ NK cells [36]. Furthermore, a subset of CD 56dim/ CD16+ NK cells have been found to express escalated quantities of CD 57,a marker correlated with substantially differentiated cells [37]. Certain CD 56dim/ CD16+ CD 57+ cells illustrate considerably escalated expression of senescence- associated genes for instance zinc finger protein36 (ZFP36) as well as DUSP2 [31], escalating liberation of cytokines [38].
Alterations in Monocyte in Elderly Males
The manner ageing impacts peripheral blood monocyte quantities is controversial. The study performed by Seidler et al. [38], illustrated practically negligible influence [39]. Nevertheless, recently sequencing studies displayed escalated quantities in older subjects [31]. Nevertheless, a plethora of studies have been in line that ageing results in escalated quantities of CD16+ monocytes, suggesting phenotypic changes in the monocytes at the time of ageing event [31, 39]. Apart from canonical CD14+ monocytes, CD16+ monocytes possess escalated inflammatory probability which facilitates the generation of the SASP [40].
Concurrently, the NFκB ,IL-1β signaling pathway in addition to inflammatory reactions signaling pathway in monocytes are substantially activated with concomitant significant up regulation of expression of proinflammatory genes for instance TNF, DDIT4 JUNB in elderly males [31].
Dendritic Cells(DCs) dynamics in Elderly Males
Dendritic cells (DCs) possess a crucial part in immune reactions in view of their working in the form of antigen presenting cell (APC).They incorporate plasmacytoid DCs (pDCs) as well as myeloid DCs (mDCs) [41]. Maximum
scientific researchers evaluating have drawn conclusions that peripheral blood DCs quantities in healthy elderly males does not illustrate noticeable alterations on contrasting with their younger compatriots [31, 42]. However, switching in pDCs as well as mDCs percentage at the time of senescence continues to be controversial [43]. Irrespective of mDCs enrichment alterations, they illustrate prominent proinflammatory properties in contrast to pDCs. Diminished phosphatidyl inositol 3 – kinase (PI3K) action which accelerates activation of NFκB, which is followed by formation of proinflammatory cytokines [42]. Senescent mDCs further revealed escalated expression of CD 68 pointing to sustenance of activation, which results in escalated proinflammatory cytokines generation despite the absence of external stimuli [44].
Alterations of Testicular Macrophages in Elderly
A Working of Testicular Macrophages: Testicular macrophages are crucial for sustenance of testicular immune prerogative, which aids in spermatogenesis as well as manipulating T generation [45]. Their division is performed into i) interstitial along withii) peritubular macrophages depending on phenotypic traits [46]. Origination of such cells takes place from embryonic in addition to bone marrow(BM) progenitors, which assume an antiinflammatory status with liberation of IL-10 as well as transforming growth factor beta( TGF –β), which assist in T generation in addition to spermatogenesis [45, 47]. Additionally, such macrophages possess the capacity of liberating 25 hydroxycholesterol [48], which promote T generation in Leydig cells.
Impact of Ageing in Testicular Macrophages: Transmission electron microscopy utilization was done in the earlier studies for evaluation of age associated changes in testicular macrophages, which displayed abberations in cellular structure for instance mitochondria as well as golgi apparatus [49]. Nevertheless, the mechanistic modes behind alterations in ageing continue to be uncharted in view of technological restrictions. Once sequencing methodologies have been introduced, a greater exhaustive insight has been achieved in reference to genetic, phenotypic , quantitative alterations in ageing macrophages. An escalated hyperactivated M1 kind macrophages in addition to considerable escalated liberation of the cytokines in older human testis have been seen [50]. Mouse studies conducted by Nie et al. [50], corroborated such observations, illustrating existence of three subpopulations. Significant escalation of subpopulation3 (senescence- particular) occurred in older mice illustrating hyperactivated as well as inflammation associated gene expression. Ageing enrichment of genes associated with subpopulation 1 along with2 had an association with type1 interferon liberation in addition to toll like receptor 4 (TLR4) signaling [13].
Immunesenescence Actions on Hypothalamic- Pituitary-Gonadal (H-P-G) Axis
Changes in peripheral blood immune cells at the time of immunesenescence significantly aids in considerable proinflammatory status in males. Such inflammatory status substantially influences working of H-P-G Axis [51]. The ageing event of hypothalamus is closely correlated with the activation of NFκB signaling pathway. Such activation of NFκB signaling pathway influences the working of GnRH neurons, which intercepts GnRH gene transcription, that terminates in aberrant GnRH liberation amongst the hypothalamus [52]. Different studies have isolated the existence of proinflammatory cytokines specifically IL-6 in the pituitary tissue in the ageing mice. Such inflammatory situation hampers the tissue healing in the mechanistic modes of the pituitary stem cells [53]. Ageing in a pronounced fashion results in expansion of proinflammatory testicular macrophages, escalating liberation of proinflammatory cytokines as well as gene expression. Such switching of macrophages has a profound impact on the testicular proinflammatory microenvironment at the time of ageing [54]. Systemic in addition to restricted inflammation might aggravate Leydig cell ageing which result in mitochondrial impairment as well as hampering of autophagy (Figure 1) [55].
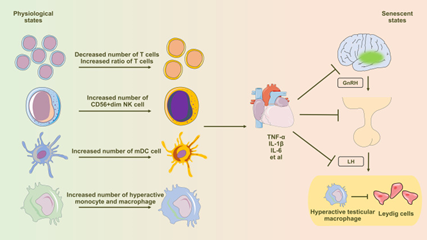
Figure 1: Courtesy Reference No-55-Diagram Illustrating the Influence of Immune Senescence-Induced Inflammaging on Testosterone Synthesis in Leydig Cells. Aging Profoundly Affects Peripheral Blood Immune Cells, Inducing Notable Alterations in T Cells, NK Cells, Dendritic Cells, Monocytes, and Macrophages. This Shift Leads to an Elevated Presence of Pro-Inflammatory Immune Cells and an Excessive Production of Pro-Inflammatory Cytokines, Subsequently Influencing the Regular Functionality of the Hypothalamic-Pituitary-Gonadal (HPG) Axis Via the Circulatory System. Testicular Macrophages, Under the Influence of Senescence, Transition Towards the M1 Subtype (Hyperactivated), Characterized by Heightened Secretion of Abundant Pro-Inflammatory Factors. These Factors are then Released Through Paracrine Signaling, Directly Impacting the Functionality of Leydig Cells.
Cellular Senescence
Cellular senescence possesses the properties of culmination of cell proliferation along with working at the time of particular situations, associated with morphological, physiological as well as molecular changes [56]. Such close biological event is closely associated with canonical propagation of organism, the ageing directions along with initiation of different diseases.
Primary Alterations in Senescent Leydig cells
Unraveling of senescent cells incapacity of going through usual cell division is imperative for cell cycles was performed by Hayflick and Moorhead’s founder work [57]. Following that evaluations have incorporated telomere length as well as working of telomerase in the form of fundamental etiological factors that aids in cell cycles arrest [58, 59]. Morphological, in addition to structural alterations usually are the properties of senescent cells whose presentation is in the form of an escalated, flattening of morphology as well as irregularity of organelles [60]. Sa β-gal works in the form of a commonly utilized biomarker which yields understanding of cellular senescence to certain magnitude despite not working in the form of a mandatory pointer [61].
Concurrently, the expression of particular cell cycle hampering proteins prominently get significantly up regulated in senescent cells [62]. Transmission electron microscopy utilization was done by Jin Y, et al. [63], where they observed illdistributed mitochondria in senescent Leydig cells [63]. Jeong MG, et al. [64], found noticeable escalated Sa β-gal quantities in addition to escalated expression of p16, p19 as well as p21 in ageing Leydig cells [64].
SASP in Senescent Leydig cells
SASP is critical cellular ageing event where senescent cells liberate different molecules influencing the microenvironment along with adjacent cells [65]. Continuation of SASP possesses the capacity of stimulating inflammation in addition to age associated conditions [66].
NFκB Signaling Pathway Activation
The NFκB signaling pathway is crucial in immune reactions, inflammation, proliferation in addition to apoptosis [67]. The activation pathways canonically implicate cell surface receptors, viral infections, bacterial constituents, as well as cytokines [68]. Translocation of NFκB takes place from cytoplasm to nucleus binding SASP associated gene promoters, which stimulate transcription [69]. Additionally, NFκB possesses the capacity of indirectly escalating the expression of the transcription factor C/EBPβ-homologous protein (Chop) followed by promoting direct controlling of SASP gene expression by C/EBPβ [70]. Amongst Leydig cells, Shang L, et al. [71] illustrated significantly activated NFκB signaling pathway [71]. Subsequent to exposure to inflammatory stimulation, mouse Leydig progenitors cells along with TM3 cell line illustrated considerable escalation of proinflammatory cytokines in the supernatant of cell cultures.
B.MAPK Signaling Pathway Activation
The MAPK signaling pathway inclusive of extracellular signal –regulated kinase (ERK1/2), c-Jun-N-terminal kinase (JNK) as well as p38 MAPK is crucial in cell events for instance proliferation, differentiation ,inflammation, along with apoptosis [72]. Freund A, et al. [73] pointed that p38 MAPK possesses an independent part in SASP, escalating NFκB’s transcriptional actions for SASP activation [73].
Concurrently p38 MAPK imparts an indirect impact by heightening the activation of transcription factors for instance activating transcription factor2 (ATF2), Activator protein 1(AP1), CREB1 [74]. Sequentially, such activated transcription factors crosstalk in the controlling of SASP. Activation of p38 MAPK was observed in ageing Leydig cells, whereas activation of ERK1/2 / JNK was not pronounced [75]. This represses crucial T generation, finally diminishing serum T quantities.
The constituents of SASP liberated by senescent Leydig cells possess the capacity of imparting paracrine actions on adjacent cells [76]. Noticeably, C/EBPβ, ATF2, AP1, in addition to CREB1 molecules are part of basic leucine zipper transcription factors [77]. Actually such molecules are closely implicated in the controlling of SASP [78]. Significantly amongst Leydig cells such molecules possess a crucial part in the controlling of T signaling pathways [79].
Mitochondrial Impairment
Leydig cellular senescence is intricately correlated with mitochondrial impairment. Ageing stimulates generation of OS in cells, therefore impacting expression quantities of crucial molecules implicated in T generation on the mitochondria. In turn this starts events for instance apoptosis in addition to necroptosis with termination in mitochondrial impairment.
Part of Mitochondria in T Generation
Mitochondria portray intracellular organelles that possesses critical part in T generation. First they possess main part in generation of ATP, the imperative energy substrates needed for T generation [80]. Secondly mitochondria portray imperative part of close signaling stepwise patterns crucial in controlling T generation in the form of molecules for instance StAR as well as CYP11A1 which reside amongst such organelles [81]. Furthermore, mitochondria, control OS along with apoptosis of Leydig cells, emphasizing their composite implications [82].
Mitochondrial Impairment in Senescent Leydig Cells
There is a significant part of mitochondrial impairment in dysfunctional T generation in senescent Leydig cell [83]. StAR expression on the outer mitochondrial membrane (OMM) of senescent Leydig cell illustrates a pronounced decrease [84], which result in structural disturbance in Leydig cell mitochondria followed by dysfunctional mitochondrial respiratory chain [electron transport chain (ETC)] working [85].
Mitochondrial impairment in Leydig cells might result in diminished generation of ATP in addition to intracellular ROS accrual [86]. Escalated ROS stimulates activation of inflammatory vesicles starting cellular pyroptosis [87]. For this event nucleotide-binding domain, leucine-rich- repeat containing family, pyrin domain-containing (NLRP3) inflammasome is responsible, where in reaction to NFκB activation inflammatory stimuli accelerates the transcription of inflammasome associated genes [88]. Activation of NLRP3 enrolls the adapter molecule ASC leading to caspase1 activation. Caspase1 results in cleavage of pro IL-1β into IL-1β as well as pro IL-18 into IL-18 leading to their liberation along with followed by apoptosis [89]. In reaction to inflammatory stimuli Leydig cell illustrates escalated intracellular NLRP3 in addition to ASC expression, cleavage of caspase1, transformation IL-1β as well as IL-18, activation of apoptosis in addition to diminished expression of T generating molecules [90]. Taken together such alterations terminate in the hampering of T generation.
Autophagy Insufficiency
Part of Autophagy in Leydig cells: Autophagy portrays an inherent catabolic event which sustains intracellular stability by breaking down dysfunctional cellular constituents [91]. In case of eukaryotic cells3 Kinds of Autophagy have been detected towards the lysosome; i) macroautophagy ii) microautophagy besides ii) chaperone modulated autophagy (CMA) [92]. Based on the organelles whose targeting takes place the produced autophagic vesicles are labelled by particular names .Like for mitochondria, peroxisomes, lipids, as well as RNA the labelling is mitophagy, peroxophagy, lipophagy as well as ribophagy respectively [93]. In Leydig cells autophagy controls OS, cholesterol uptake, apoptosis pyroptosis which impacts T generating capability [90, 94].
Autophagy Insufficiency Along with its Sequelae in Leydig Cells: Dynamic behaviour is illustrated amongst cells by mitochondria which persistently go through fusion in addition to division events [95]. The expression of prefission proteins get stimulated by mitochondrial fission therefore inducing a synonymous changes in the magnitude of mitophagy [96]. Recent, work on Leydig cells emphasized escalated N6 methyladenosine (m6A) quantities in primary Leydig cells obtained from senescent mice .Such escalated m6A intercepts intracellular autophagy, impacts the working capacity of T generation [97]. Redox dysequilibrium gets stimulated by diminished autophagy in Leydig cells, leading to intracellular ROS accrual [94]. Mitochondrial fusion balance got disrupted subsequent to decontrolling of ROS quantities influencing autophagy. Yi L, et al. [98] held cadmium stimulated apoptosis in Leydig cells responsible for mitochondrial fragmentation, resulting in dysfunctional working as well as escalated superoxide in addition to ROS quantities, hampering mitochondrial autophagy [98]. Reduced autophagy in Leydig cells further starts pyroptosis. Inflammatory stimuli decrease autophagy, which activation of inflammasome along with starts pyroptosis [90] (Figure 2).
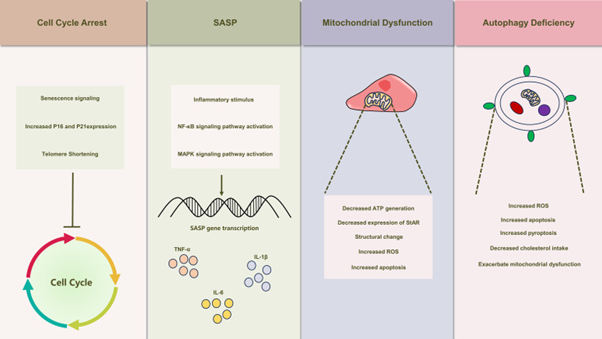
Figure 2: Courtesy Reference No-55-Illustrates the Schematic Depiction of Senescence-Induced Alterations in Leydig Cells. Leydig Cell Senescence Manifests a Spectrum of Modifications, Encompassing Cell Cycle Arrest, The Emergence of Senescence-Associated Secretory Phenotypes (SASP), Mitochondrial Dysfunction, and Autophagy Deficiency. These Alterations Also Play a Crucial Role in the Development of Testicular Inflammaging, Leading to Substantial Inhibition of Leydig Cell Function.
Plausible Therapeutic Approaches
The consensus on management of LOH is testosterone replacement’ therapy (TRT). However, TRT is associated with inimical sequelae whose avoidance is not feasible [99]. The alternate innovative approaches variable from TRT are described here which possess the capacity of attenuating LOH by abrogating Inflammageing (Figure 3 for pathophysiological alteration necessitating therapies.
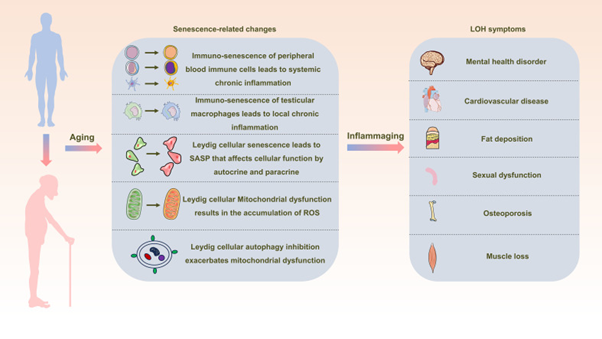
Figure 3: Courtesy Reference No-55-Schematic Representation Depicting the Interrelation between Inflammaging and Late-Onset Hypogonadotropic Hypogonadism (LOH). As Aging Progresses, Alterations in Peripheral Blood Immune Cells, Testicular Macrophages, and Leydig Cells Become Pronounced, Creating a Milieu of Widespread and Localized Inflammation. This State, Termed Inflammaging, Disrupts the Intricate Process of Testosterone Synthesis within Leydig Cells, Consequently Initiating and Advancing the Development of LOH in Aging Males.
Physical Exercise
Physical exercise has come out to be natural in addition to exhaustive strategy in reference to management of health, illustrating an obvious influence on clinical symptomatology correlated with different diseases [100]. In reference to LOH appropriate physical exercise possesses the probability of attenuating systemic inflammation, escalate cellular antioxidant stress capability, as well as escalate T quantities in cases. Various randomized controlled trial implicating subjects amongst 40-75yrs those taking part in attempting interval aerobic exercises at the magnitude reaching 90% of the maximum heart rates three times/week resulted in considerable decrease in the circulating CRP quantities along with TNF-α in the subjects enrolled in the study [101]. Clinical trials have corroborated that elderly subjects taking part in routine physical exercise have presentation of potentiating quantities of antioxidant capability markers in the blood inclusive of glutathione peroxidase (GPx), full nitrite/ nitrate in addition to full oxyradicals foraging capability in contrast to their sedentary compatriots [102]. Furthermore moderate magnitude of aerobic exercises have illustrated pronounced capability of considerably escalating T quantities in older male subjects [103].
Antiinflammatory along with Antioxidant Therapies
Adrenomedullin (ADM), portrays a peptide hormone generated mainly by adrenal medulla possesses a wider existence testis [104]. ADM confers robust protection in inflammatory events [105]. Primary Leydig cells in rats generate ADM as well as express its receptors pointing to its autocrine, along with paracrine part in guaranteeing T generation [106]. ADM plausibly confers protection at the time of inflammation by intercepting ROS generation, provision of stability to mitochondria, hampering NFκB’s and MAPK signaling, in addition to escalating autophagy which counteracts on apoptosis along with pyroptosis [107]. Antioxidants which are targeted against mitochondria directly crosstalk with mitochondria for counteracting OS as well as injury [108]. SS peptides particularly synthetic tetrapeptide, elamipretide (SS-31) have been substantially worked on in reference to their influence on mitochondrial working[109]. As per reports, SS-31 basically crosstalk with the phospholipid cardiolipin in the inner mitochondrial membrane (IMM). Chavez ID, et al. [109] utilized chemical cross-linking with mass spectrometry to isolate protein crosstalkers of SS-31 in mitochondria. The SS-31- crosstalking proteins, all acknowledged cardiolipin binders, placed into two groups, those implicated in ATP generation production via the oxidative phosphorylation pathway as well as and those implicated involved in 2-oxoglutarate metabolic processes crosstalkers Residues cross-linked with SS-31 binding regions display that in plethora of instance, are proximal to cardiolipin-protein crosstalking areas [109]. Working of SS31 takes place at the level of the cells decreasing the neutralizing ROS production, which attenuates OS amongst cells as well as mitochondria in an efficacious manner [110]. It possesses a critical part in the sustenance of mitochondrial membrane intactness therefore sustenance of equilibrium amongst fission along with fusion, critical for cellular energy metabolism along with working [111]. Additionally, it is substantiated that SS31 possesses antiinflammatory characteristics, manipulating inflammatory mediator generation cellular reactions to inflammation [112]. Its therapeutic capability encompasses different inflammatory in addition to age associated situations inclusive of cardiovascular disease (CVD), Kidney disorders as well as Alzheimer’s disease [113]. At present ADM along with SS31in particular diseases have propagated to phase II clinical trial [114]. Greater evaluations are necessary for illustrating the safety of such agents in clinical scenario as well as substantiate its effectiveness for tackling LOH.
Transplantation of Leydig cells
Currently cell transplantation has attracted considerable interest in the form of therapeutic strategy for variety of diseases. Utilization of methodologies for addressing cell Transplantation for tackling LOH incorporates i) Leydig cells transplantation ii) transplantation of Leydig stem cells in addition to iii) stem cells Transplantation. Luo P, et al.
[115] performed a study by utilization of autofluorescence dependent identification as well as purification of Leydig cells. This was followed by transplantation of identified Leydig cells subcutaneously into denuded mice leading to escalated serum testosterone quantities [115]. Arora H, et al. [116] illustrated that simultaneous subcutaneous transplantation of Leydig stem cells, assisting cells along with myoid cells promoted the differentiation in addition to maturation of Leydig cells resulting in liberation of testosterone [116]. Administration of bone marrow (BM), mesenchymal stem cells (MSC’s) in the rats tail vein has been found to escalate serum testosterone quantities. For such actions diminished OS along with senescence phenotype is implicated in Leydig cells [117].
Part of Herbal Medicine
Different plant based medicines which are constituents of Chinese canonical medicine with the idea escalating sex hormones quantities in addition to sexual working have been found to be favourable in LOH. Yangying capsules have been formed by the group of Sun, Jin et al illustrating a positive influence on testosterone formation [118]. Complicated herbal constitutions are bothersome in indicating the precise constituents as well as gaining insight in reference to clinical effectiveness. Present work has concentrated on separate herbs along with their extracts corroborating direct actions on Leydig cells on T generation.
Icariin obtained from Epimedium simulates sex hormones, escalate immunity in addition to decrease inflammation [119]. Different studies have validated that icariin stimulated testosterone generation factor which implicated the upregulation of T generation correlated molecules for instance Esr1/Src/Akt/Creb/Sf1 pathways, conservation of mitochondrial working as well as other mechanistic modes behind [120]. Polysaccharides obtained from Polygonatum sibricum recognized for escalating sexual along with kidney working, illustrate antiinflammatory in addition to antioxidant actions [121].
Such polysaccharides guarantee T generation in mice diminishing Oxidative stress, conservation of mitochondrial working as well as hampering mitochondria modulated apoptosis [122]. Furthermore, variety herbs have been evaluated for their capability of endorsing as well as protecting T generation [123], which makes it mandatory to perform exhaustive assessment of different experimental avenues.
Ashwagandha alias Withania somnifera gets substantially used in Indian canonical medicine. It works favorably in symptoms correlated with LOH via plausible advantages inclusive of diminished stress, antioxidant characteristics, antiinflammatory along with immunomodulatory actions, promotion of T generation apart from other mechanistic modes [124].
Conclusions
As Figure3 summarizes separate pathophysiological alteration that necessitate therapies in ageing men.In a nutshell like in obesity chronic low grade inflammation is a part and parcel of ageing as emphasized by us in ovarian ageing and need for use of correcting NAD+ metabolism in ageing with precursor substances Nicotinamide riboside (NR) as well as nicotinamide mononucleotide (NMN) lead to escalated SASP resulting in escalated chronic inflammationand alters inflammageing, inflammation Cellular Senescence ,immunity, DNA metabolism, here we have summarized the role of chronic low grade inflammation alias inflammageing and immunosenescence(alteration in innate along with adaptive immunity in LOH [rev by us in ref 19 & 20).
Testosterone generation by Leydig cells (LCs) possess a critical part in male reproduction. The working of LCs that have degenerated possess the capacity of testosterone insufficiency, which finally might result in primary male hypogonadism.
Transplantation of exogenous LCs with the capability of generating testosterone in reaction to the controlling of the H-P-G axis might be a favourable alternative methodology for treating male primary hypogonadism. Recent studies have illustrated the probability of producing Leydig-like cells from stem cells by different strategies. Additionally, somatic cells, for instance embryonic or adult fibroblasts, have further got reprogrammed into Leydig-like cells with success. Recently Li Z, et al. [125] reviewed the recent advances in the production of Leydig-like cells, highlighting on contrasting the efficacy along with safety of different protocols used in addition to the cells generated.
By further evaluating the properties of Leydig-like cells generated from fibroblasts dependent on small signaling molecules as well as controlling factors, they observed that despite the cells might generate testosterone, they are significantly separate from real LCs.
For future in vivo applications, it is significant that the steroidogenic cells generated be analyzed apart from their steroidogenic working however further for their full cell metabolic state by proteomics or transcriptomic tools (Figures 4-6).
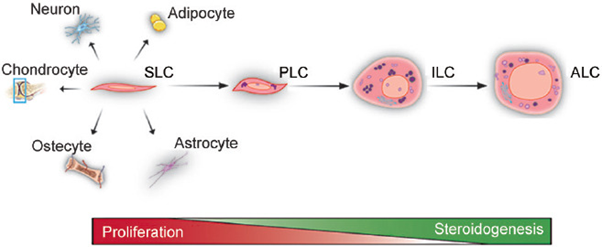
Figure 4: Courtesy Reference No-125-The Differentiation and Steroidogenic Processes of ALCS. In Rat Testes, the Differentiation of Adult Leydig Cells Includes Four Stages: SLCs, PLCs, ILCs, and ALCs. SLCs are Capable of Self-Renewal but not of Producing Androgens. PLCs Can Proliferate 4 Rounds and Synthesize Androsterone as A Major Product. ILCs Proliferate Only Once and Produce 5α-Androstane-3α, 17β-Diol as a Major Product. ALCs Lose The Ability to Proliferate and Produce Testosterone as a Final Product. The Wedge Icons Represent the Proliferative or Steroid-Producing Ability of Leydig Cells. SLC: Stem Leydig Cell; PLC: Progenitor Leydig Cell; ILC: Immature Leydig Cell; ALC: Adult Leydig Cell.
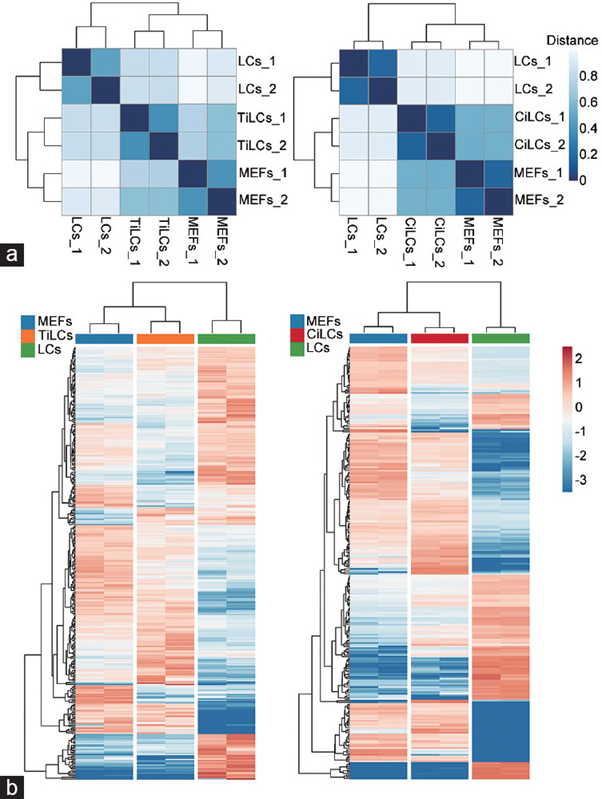
Figure 5: Courtesy reference no-125-The similarity analysis of gene expression profiles and transcription factors between MEFs, Leydig-like cells, and LCs. (a) The similarity (Jaccard distance) of global gene expression profiles. The expression data of samples were extracted from 2 datasets GSE87020 and GSE145797 deposited in public databases. The data were log- transformed normalized and computed. The color difference of the distance represents the degree of similarity in expression profiles. The darker color means a higher similarity between samples. Left: comparisons among MEFs, LCs and TiLCs. Right: comparisons among MEFs, LCs and CiLCs. (b) Hierarchical clustering analysis of differentially expressed TFs. A total of 353 differentially expressed TFs among MEFs, LCs and Leydig-like cells were identified (fold change >1.5 and P < 0.05). The expression level of each transcriptional factor was normalized based on a global z-score transformation. The hierarchical tree on the left and on the top of the heatmap represents the clustering of TFs and the samples, respectively. Each column shows 2 cell replicates, and the color scales range from dark red to bright blue, corresponding to the up- or downregulation of TF expression, respectively. Left: comparisons among MEFs, LCs, and TiLCs. Right: comparisons among MEFs, LCs, and CiLCs. MEFs: mouse embryonic fibroblasts; TiLCs: transcription factor-based induced Leydig-like cells; CiLCs: compound-based induced Leydig-like cells; LCs: Leydig cells; TF: transcription factor.
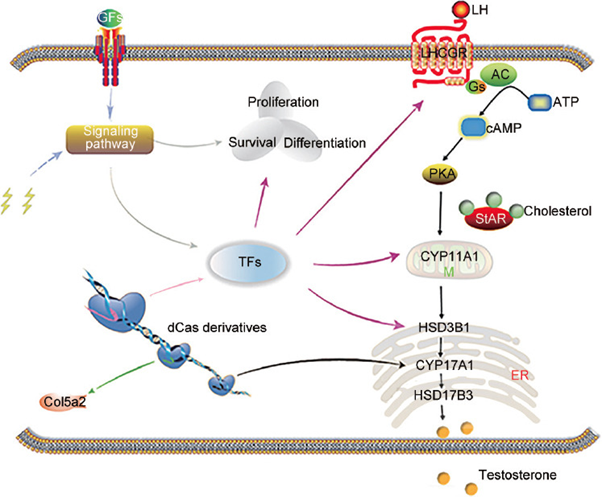
Figure 6: Courtesy reference no-125-Diagram of Leydig-like cells generated from host cells using chemicals, growth factors, and CRISPR/Cas9 technologies. The CRISPR/Cas derivatives are capable of simultaneously activating or repressing targeted genes. Growth factors or chemicals can regulate the expression of genes and biological process by mediating signaling pathways. Growth factors or chemicals (agonists or antagonists) can regulate the biological function and gene expression by mediating signaling pathways. The CRISPR/Cas9 system, growth factor, and chemical promote synergistically the host cell reprogramming into steroid-producing Leydig-like cells. Lightning icon represents the agonists or antagonists of signaling pathways. CRISPR/Cas: clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9; Col5a2: collagen type V alpha 2 chain; GFs: growth factors; TFs: transcription factors; M: mitochondria; ER: endoplasmic reticulum; LH: luteinizing hormone; LHCGR: luteinizing hormone/choriogonadotropin receptor; ATP: adenosine triphosphate; Gs: G proteins; AC: adenylyl cyclase; cAMP: cyclic adenosine monophosphate; PKA: protein kinase A; StAR: steroidogenic acute regulatory protein.
References
-
Yeap BB, Manning L, Chubb SAP, Handelsman DJ, Almeida OP, et al. (2018) Progressive impairment of testicular endocrine function in ageing men: testosterone and dihydrotestosterone decrease, and luteinizing hormone increases in men transitioning from 8th to 9th decade of life. Clin Endocrinol (Oxf) 88(1): 88-95.
-
Zirkin BR, Papadopolous V (2018) Leydig cells formation, function and regulation. Biol Reprod 99(1): 101-111.
-
Nieschlag E (2020) Late-onset hypogonadism: a concept comes of age. Andrology 8(6): 1506-1511.
-
Camacho EM, Huhtaniemi IT, O’Neill TW, Finn JD, Pye SR, et al. (2013) Age associated changes in hypothalamic- pituitary- testicular function in middle aged and older men are modified by weight changes and lifestyle factors : longitudinal results from the European male ageing study. Eur J Endocrinol 168(3): 445-455.
-
Mularoni V, Esposito V, Di Persio S, Vicini E, Spadetto G, et al. (2020) Age related changes in human Leydig cell status. Hum Reprod 35(12): 2663-2676.
-
Chung JY, Chen H, Zircan B (2021) Sirt1 and Nrf2: regulation of Leydig cell oxidant/antioxidant intracellular environment and steroid formation. Biol Reprod 105(5): 1307-1316.
-
Fulop T, Larbi A, Pawelek G, Khalil A, Cohen AA, et al. (2023) Immunology of ageing: The birth of Inflammaging. Clin Rev Allergy Immunol 64(2): 109-122.
-
Franceschi C, Campisi J (2014) Chronic inflammation (inflammageing) and its potential contribution to age associated diseases. J Gerontol A Biol Sci Med Sci 69(S1): S4-S9.
-
Oliveiri F, Rippo MR, Procopio AD, Fazioli F (2013) Circulating inflamma-miRs in ageing and age related diseases. Front Genet 4: 121.
-
Abd El-Kader SM, Al-Shareef FM (2018) Inflammatory cytokines and immune system modulation by aerobic versus resisted exercise training for elderly. Afr Health Sci 18(1): 120-131.
-
Lopes-Paciencia S, Saint-Germain E, Rowell MC, Ruiz AF, Kalegiri P, et al. (2019) The senescence- associated secretory phenotype and its regulation. Cytokine 117: 15-22.
-
Pelletier RM (2011) The blood testis barrier: the junctional permeability proteins and thelipids. Prog Histochem Cytochem 46(2): 49-127.
-
Zhang W, Xia S, Xiao W, Song Y, Tang L, et al. (2022) A single cell transcriptomic landscape of mouse testicular ageing. J Adv Res 53: 219-34.
-
Kaur KK, Allahbadia GN, Singh M (2020) An update on future utilization of Extracellular Vesicles in evaluation part of these in maturation of gametes ,fertilization along with embryo implantation for escalating the success of Artificial Reproductive Technology: A Systematic Review. Open Access Journal of Gynecology and Obstetrics 3(2): 10-29 .
-
Kaur KK, Allahbadia GN, Singh M (2022) An Update on the Innovative Part of NLRP3 Inflammasome Regarding Newer Strategies for Treatment of Reproductive Conditions Possessing Greater Risk: A Systematic Review. J Gynecol 7(3): 1-13.
-
Kaur KK, Allahbadia GN, Singh M (2023) Are we any Close to Utilizing Targeting SIRT Signaling Pathway for Enhancingthe Oocyte Quality in Women with Advanced Maternal age: Bringing from Bench to Bedside: A Narrative Review. J Gynecol 8(2): 1-17.
-
Kaur KK, Allahbadia GN, Singh M (2024) An update on Telomeres Dynamics in Reproduction; Specifically in Females &Ageing: A Narrative Review. Open J of Gyneocol 9(2): 1-13.
-
Kaur KK, Allahbadia GN, Singh M (2024) Exciting hope for Professional Women marrying Late - manipulating NAD+ Metabolism -A-Bright Future Prospect-A narrative review. Chronicles of Clinical Reviews and Case Reports The Geek Chronicles 1: 1-30.
-
Kaur KK, Allahbadia GN, Singh M (2024) An Update on Ageing Associated Immunosenescence and Inflammageing in Infectious Diseases: A Narrative Review.
-
Kaprara A, Huhtaniemi IT (2018) The hypothalamic- pituitary- gonadal axis: tales of mice and men. Metabolism 86: 3-17.
-
Xie Q, Kang Y, Zhang C, Xie Y, Wang C, et al. (2022) The role of kisspeptins in the hypothalamic-pituitary- gonadal axis and reproduction. Front Endocrinol(Lausanne) 13: 925206.
-
McCann SM, Arubayashi U, Sun HQ, Yu WH (1993) Control of follicle stimulating hormone and luteinizing hormone release by hypothalamic peptides. Ann N Y Acad Sci 687: 55-59.
-
Choi J, Smitz J (2014) Luteinizing hormone and human chorionic Gonadotropins: of differences. Mol Cell Endocrinol 383: 203-213.
-
Miller WL (2013) The steroid hormone synthesis in mitochondria. Mol Cell Endocrinol 379: 62-73.
-
Hebert Schuster M, Rotta BE, Kirkpatrick B, Guibourdenche J, Cohen M (2018) The interplay between glucose regulated protein78(GRP78) and steroids in the reproductive system. Int J Mol Sci 19(7): 1842.
-
Payne AH, Hales DB (2004) Overview steroidogenic enzymes in the VDAC pathway from cholesterol to active steroid hormones. Endocrin Rev 25: 947-970.
-
Wang Y, Dong C, Han Y, Gu Z, Sun C (2022) Immunesenescence, ageing and successful ageing. Front Immunol 13: 942796.
-
Pawelek G (2018) Age and Immunity: what is immunosenescence. Exp Gerontol 105: 4-9.
-
Franceschi C, Bonafe M, Valensin S, Oliveiri F, DeLuca M, et al. (2000) Inflamm-ageing: an Evolutionary perspective on immunosenescence. Ann N Y Acad Sci 908: 244-254.
-
Huang Z, Chen B, Liu X, Li H, Xie L,Gao Y, et al. (2021) Effect of sex and ageing on the immune landscape as assessed by single cell transcriptomic analysis. Proc NatlAcad Sci USA 118(33): e2023216118.
-
Martinez Zamudio RJ, D HK, Vasilopolous T, Gitter Williams L, Herbig U (2021) senescence- associated β- galactosidase the abundance of senescent CD8+T cells inageing humans. Ageing Cell 20: e13344.
-
DesdnMico G, Soto-Herendero G, Aranda JF, Offer J, Carrasac E, et al. (2020) T cells with dysfunctional mitochondria induce multi-morbidity and premature senescence. Science 368: 1371-1376.
-
Poznanski SM, Ashkar AA (2019) What defines NK cells functional fate: phenotype or metabolism. Front Immunol 10: 1414.
-
Wendt K, Wilk E, Buyney S, Buer J, Schmit RE, et al. (2006) Gene and protein characteristics reflect functional diversity of CD 56dim and CD 56bright NK cells. J Leukocyt Biol 806: 1529-1541.
-
Khummuang S, Chuensirikulchai K, Pata S, Laopajan W, Chruekmlow N, et al. (2019) Characterization and functional analysis of novel circulating NK cells sub- populations. Int Immunol 31: 515-530.
-
Bjorkstrom NK, Riese P, Heuts F, Andersson S, Fauriat C, et al. (2010) Expression patterns of NKG2A,KIR and CD57 define a process of CD 56dim NK cell differentiation uncoupled from NK cell education. Blood 116: 3853- 3864.
-
Liu M, Meng Y, Zhang L, Han Z, Feng X (2021) High efficient generation of n Natural killer cells from peripheral blood with preferred cell vitality and enhanced cytotoxicity by combination of IL-2, IL-15, and IL-18. Biochim Biophys Res Commun 534: 149-150.
-
Seidler S, Zimmermann HW, Bartneck M, Trautwein C, Tacke F (2010) Age dependent alterations of monocyte subsets and age related monocyte chemokine pathways in healthy adults. BMC Immunol 10: 30.
-
Ong SM, H E, Dang TM, Yeap WH, Tan CT, et al. (2018) The pro-inflammatory phenotype of the non-classical monocytes subsets is attributed to senescence. Cell Death Dis 9: 266.
-
Macri C, Pang M, Patton T, O’Keefe M (2018) Dendritic cells subsets. Semin Cell Dev Biol 84: 11-21.
-
Agrawal A, Agrawal S, Cao JN, Su H, Gupta S (2007) Altered innate immune functioning of dendritic cells in elderly humans: a role of phosphatidyl inositol 3 - kinase signaling pathway. J Immunol 178(11): 6912-6922.
-
Agrawal A, Sridharan A, Prakash S, Agrawal H (2012) Dendritic cells and ageing: consequences for autoimmuninty. Exp Rev Clin Immunol 8(1): 73-80.
-
Agrawal A, Tay J, Agrawal S, Ton S, Gupta S (2009) Increased reactivity of Dendritic cells from aged subjects to self-antigen, the human DNA. J Immunol 182(2): 1138-1145.
-
Schechter R, London A, Schwartz M (2013) Orchestrated leukocyte recruitment to immune privileged sites: absolute barriers versus educational gates. Nat Rev Immunol 13(3): 206-218.
-
Keller NM, Gentek R, Giminez G, Bigot S, Mailfert S, et al. (2017) Developmental origin and maintenance of distinct testicular macrophages populations. J Exp Med 214(10): 2829-2841.
-
Fijak M, Meinhart A (2006) The testis in immune privilege. Immunol Rev 213: 66-81.
-
Lukyanenco YO, Chen JJ, Hutson JC (2001) Production of 25 hydroxycholesterol by testicular macrophages and its effects on Leydig cells. Biol Reprod 64(3): 790-796.
-
Giannessi F, Giambelluca MA, Scavuzzo MF, Ruffoli R (2005) Ultrastructure of testicular macrophages in ageing mice. J Morphol 263(1): 39-46.
-
Nie X, Munyoki SK, Sukhwani M, Schmidt N, Missel A, et al. (2022) Single cell analysis human testis ageing and correlation with elevated body mass index. Dev Cell 57(9): 1160-1176.
-
Cai D, Khor S (2019) Hypothalamic microinflammation paradigm in ageing and age related diseases. Cell Metab 30(1): 19-35.
-
Zhang G, Li J, Purkayastha S, Tang Y, Zhang H, et al. (2013) Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nature 497(7448): 211-216.
-
Vennekens A, Laporte E, Hermans F, Cox B, Janiskowski A, et al. (2021) Interleukin-6 is an activator pituitary stem cells upon local damage: a competence quenched in the ageing gland. Proc Natl Acad Sci, USA, 118(25): e2100052118.
-
Hales DB (2002) Testicular macrophage modulation Leydig cell steroidogenesis. J Reprod Immunol 57(1-2): 3-18.
-
Xing D, Jin Y, Jin B (2024) A Narrative Review on inflammageing and late more onset hypogonadism. Front Endocrinol (Lausanne) 15: 1291389.
-
Roger L, Tomas F, Gire V (2021) Mechanisms and regulation of cellular senescence. Int J Mol Sci 22(23): 13173.
-
Hayflick L, Moorhead PS (1961) The serial cultivation of human diploid cell strains. Exp Cell Res 25(3): 585-621.
-
Giardini MA, Segatto M, da Silva MS, Nunes VS, Cano M (2014) Telomere and telomerase biology. Prog Mol Biol Trans Sci 125: 1-40.
-
Rssiello F, Jurk D, Passos JF, D’Adda DFF (2022) Telomere dysfunction in ageing andage related diseases. Nat Cell Biol 24: 135-147.
-
Habiballa I, Salmonowicz H, Passos JF (2019) Mitochondria and cellular senescence: implications for musculoskeletal ageing. Free Radical Res 132: 3-10.
-
Lee BY, Han JA, Im JS, Morrone A, Johung K, et al. (2006) Senescence- associated β- galactosidase is lysosomal β- galactosidase. Ageing Cell 5(2): 187-195.
-
Xie J, Wang Y, Lu L, Liu L, Yu X, et al. (2021) Cellular senescence in knee osteoarthritis: molecular mechanisms and therapeutic implications. Ageing Res Rev 70: 101413.
-
Jeong MG, Song H, Shin JH, Jeong H, Kim HK, et al. (2017) Transcriptional co activator with PDZ binding proteins is required to testicular function. Ageing Cell 16: 1035- 1042.
-
Birsch J, Gil J (2020) Senescence and the SASP: many therapeutic avenues. Genes Dev 34(23-24): 1565-1576.
-
Midha A, Pan H, Abarca C, Andle J, Carapeto P, et al (2021) Unique human and mouse beta senescence- associated secretory phenotype(SASP) reveal conserved signaling pathways and heterogenous factors. Diabetes 70: 1098- 1116.
-
Oeckinghaus A, Hayden MS, Ghosh S (2011) Crosstalk in NFκappaB signaling pathways. Nat Immunol 12: 695- 708.
-
Vallabhpurapu S, KarinM (2009) Regulation and function of NFκappaB transcription factor in the immune system. Annu Rev Immunol 27: 693-733.
-
Salminen A, Kauppinen A, Kaarniranta K (2012) Emerging role of NFκappaB signaling in the induction of senescence- associated secretory phenotype (SASP). Cell Signal 24: 835-845.
-
Sebastian T, Malik R, Thomas S, Sage J, Johnson PF (2005) C/EBP- beta cooperates with RB:E2F to implement Ras (V12) induced cellular senescence. EMBO J 24(18): 3301-3312.
-
Shang T, Zhang W, Wang T, Sun B, Deng T, et al. (2011) T toll like receptor initiated testicular innate immune response in mouse Leydig cells. Endocrinology 152(7): 2827-2836.
-
Kim EK, Choi EJ (2015) Compromised MAPK signaling in human diseases: an update. Arch Toxicol 89: 867-882.
-
Freund A, Patil CK, Campisi J (2011) p38 MAPK signaling is a novel DNA damage respone independent regulator of senescence- associated secretory phenotype. EMBOJ 30: 1536-1548.
-
Liu P, Gao Q, Guan L, Sheng W, Hu Y, et al. (2020) Atorvastatin attenuates isoflurane induced activation of ROS- p38 MAPK / ATF2 pathway, neuronal degeneration and cognitive impairment. Front Aging Neurosci 12: 620946.
-
Luo D, QiX, Xu X, Yang L, Yu C, et al. (2023) Involvement of p38 MAPK in Leydig cell ageing in age related decline in testosterone. Front Endocrinol (Lausanne) 14: 10188249.
-
Acosta JC, Banito A, Wuestefeld T, Georgilis A, Janich P, et al. (2013) A complex secretory programme orchesterated by the inflammasome controls paracrine senescence. Nat Cell Biol 15: 978-990.
-
Deppmann CD, Alvania RS, Taparowski EJ (2006) Crossspecies annotation of basic leucine zipper transcription factor interactions: insight into the evolution of closed interaction networks. Mol Biol Evol 23(8): 1480-1492.
-
Salotti J, Johnson PF (2019) Regulation of senescence and senescence- associated secretory phenotype transcription factor by the C/EBP- beta. Exp Gerontol 128: 110752.
-
Martin LJ, Nguyen HT (2022) Basic leucine zipper transcription factors as regulator of Leydig cell functions. Int J Mol Sci 23(21): 12887.
-
Roger AJ, Muñoz-Gomez SA, Kamikawa R (2017) The origin and diversification of mitochondria. Curr Biol 27(21): R1177-R1192.
-
Galano M,Li Y, LiL, Sottas C, Papadopolous V (2021) Role of constitutive StAR in Leydig cells. Int J Mol Sci 22(4): 3.
-
Midzak AS, Chen H, Aon MA, Papadopolous V (2011) ATP synthesis, mitochondrial function and steroid biosynthesis in rodent and tumor Leydig cells. Biol Reprod 84: 976-985.
-
Midzak AS, Chen H, Aon MA, Papadopolous V, Zircan BB (2009) Leydig cell ageing and the mechanisms of reduced testosterone synthesis. Mol Cell Endocrinol 299: 23-31.
-
Culty M, Luo L, Yao ZX, Chen H, Papadopolous V, et al. (2012) Cholesterol transport, peripheral Benzodiazepine receptor and steroidogenesis in ageing Leydig cells. J Androl 23: 439-447.
-
Galano M, Papadopolous V (2022) Role of constitutive StAR in mitochondrial structure and function in MA Leydig cells. Endocrinology 163(8): bqac091.
-
Beattle MC, Adekola L Papadopolous V, Chen H, Zircan BB. (2015) Leydig cell ageing and Hypogonadism. Exp Gerontol 68: 87-91.
-
Zhou R, Yazdi ASMPTJ (2011) A Role for mitochondria in NLRP3 inflammasome activation. Nature 469: 221-255.
-
Afonina IS, Zhong Z, Karin M, Beyaert R (2017) Limiting inflammation, the negative regulator of NFκappaB and the NLRP3 inflammasome. Nat Immunol 18(8): 861-869.
-
Chen X, Liu G, Yuan Y, Wu G, Wang S, et al. (2019) NEK7 interacts with NLRP3 to modulate pyroptosis in inflammatory bowel disease via NFκappaB signaling. Cell Death Dis 10(12): 906.
-
Li MY, Zhu XI, Zhao BX, ShiI, Wang W, et al. (2019) Adrenomedullin alleviates the pyroptosis of Leydig cells by promoting autophagy via the ROS AMPK mTOR axis. Cell Death Dis 10(7): 489.
-
Mizushima N, Komatsu M (2011) Autophagy: renovation of cells and tissues. Cell 147(4): 728-741.
-
Duraes FV, Niven J, Dubrot J, Hugues S, Gannage M (2015) Macroautophagy in endogenous processing of self and pathogen derived antigens for MHC class II presentation. Front Immunol 6: 459.
-
Wang L, Klionski DJ, Shen HM (2023) The emerging mechanisms and functions of microautophagy . Nat Rev Mol Cell Biol 24(3): 186-203.
-
Yahyavy S, Valizadeh A, Saki G, Khormandi L (2020) Taurine inducesautophagy and inhibits Oxidative stress in mice Leydig cells. Ibra Assist Reprod 24(3): 250-256.
-
Tandler B, Haopell CL, MearsJA (2018) Morphological pathways of mitochondrial division. Antioxidants( Basel) 7(2): 30.
-
TwigG, Elorza A, Molina A, Mohamed H, Wikstrom JD, et al. (2008) Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J 27(2): 433-446.
-
Chen Y, Wang J, Xu D, Xiang Z, Ding J, et al. (2021) A mRNA methylation regulates testosterone synthesis through modulating autophagy. Autophagy 17(2): 457-475.
-
Yi L, Shang XJ, Lv L, Wang Y, Zhang J, et al. (2022) Cadmium induced apoptosis of Leydig cells is mediated by mitochondrial fission and inhibition of mitophagy. Cell Death Dis 13: 928.
-
Barbonetti A, D’Andrea S, Francavilla S (2020) Testosterone replacement’ therapy. Andrology 8: 1551- 1566.
-
Collado MD, Lavin PAM, Penacoba C, Del CJ, Leyton RM, et al. (2021) Key factors associated with adherence to Physical exercise in patients with chronic disease and older adults: anumbrella review. Int J Environ Res Public Health 18(4): 2023.
-
Pedersen LR, Olsen RH, Anholm C, Astrup A, Eugen- Olsen J, et al. (2019) Effects of 1year exercise training versus combined exercise training and weightloss on body composition, low grade inflammation and lipids in overweight patients with coronary artery disease: a randomized trial. Cardiovasc Diabetol 18(1): 127.
-
Franzoni F, Ghidiaoni I, Galetta F, Plantinga Y, Lubrano Y, et al. (2005) Physical activity , plasma antioxidant capacity and endothelium dependent vasodilation in young and older men. Am J Hypertens 18(4): 510-516.
-
Leitao AE, Souza Vieira MC, Pelegrini A, Da Silva EL, Azevedo Guimarães AC (2021) A 6-month, double- blind, placebo-controlled, randomized trial to evaluate the effect of Eurycoma longifolia (Tongkat Ali) and concurrent training on erectile function and testosterone levels in androgen deficiency of aging males (ADAM). Mauritas 145: 78-85.
-
Li YY, Hwang ISS, Wai-Sum O, Tang F (2006) Adrenomedullin peptide: gene expression of adrenomedullin, its receptors and receptor activity modifying proteins, and receptor binding in rat testis- -actions on testosterone secretion. Biol Reprod 75(2): 183-188.
-
Xian X, Sakurai T, Kamiyoshi A, Ichikawa Shindo Y, Tanaka M, et al. (2017) Vasoprotective activities of the adrenomedullin-RAMP2 system in endothelial cells. Endocrinology 158(5): 1359-1372.
-
Chan YF, Wai-Sum O, Tang F (2008) Adrenomedullin in the rats testis its production actions on testosterone secretion, regulation by human chorionic Gonadotropin and its interactions endothelin in Leydig cells. Biol Reprod 78(4): 773-779.
-
Hu W, Shi L, Li MY, Zhou PH, Qiu B, et al. (2017) Adrenomedullin protect Leydig cells against lipopolysaccharide induced Oxidative stress and inflammatory reactions viaMAPK /NFκappaB signaling pathways. Sci Rep 7(1): 16479.
-
Szeto HH, Schiller PW (2011) Novel therapies targeting inner mitochondrial membrane- from discovery to Clinical development. Pharma Res 28(11): 2669-2679.
-
Chavez ID, Tang X, Campbell MD, Reyes G, Kramer PA, et al. (2020) Mitochondrial protein interaction landscape of SS-31. Proc NatlAcad Sci USA 117(26): 15363-15373.
-
Shang L, Ren H, Wang S, Liu H, Hu A, et al. (2021) SS-31 protects liver from ischaemia reperfusion injury via modulating macrophage polarization. Oxid Med CellLongev 2021: 6662156.
-
Peng X, Wang K, Zhang C, Bao JP, Vlf C, et al. (2021) The mitochondrial antioxidant SS-31 attenuated lipopolysaccharide-induced apoptosis and pyroptosis of nucleus pulposus cells via scavenging mitochondrial ROS and maintaining the stability of mitochondrial dynamics. Free Radical Res 55(11-12): 1080-1093.
-
Mo Y, Deng X, Zhang L, Huang Y, Li W, et al. (2019) SS- 31 reduces inflammation and oxidative stress through the inhibition of Fis1 expression in lipopolysaccharide- stimulated microglia. Biochim Biophys Res Commun 520(1): 171-178.
-
Zhu Y, Luo M, Bai X, Li J, Nie P, et al. (2022) SS-31, a Mitochondria-Targeting Peptide, Ameliorates Kidney Disease. Oxid Med CellLongev 2022: 1291509.
-
Kita T, Ashizuka S, Takeda T, Matsumoto T, Ohmiya N, et al. (2022) Adrenomedullin for biologic-resistant Crohn’s disease: A randomized, double-blind, placebo- controlled phase 2a clinical trial. J Gastroenterol Hepatol 37(11): 2051-2059.
-
Luo P, Feng X, Deng R, Wang F, Zhang Y, et al. (2021) An autofluorescence-based isolation of Leydig cells for testosterone deficiency treatment. Mol Cell Endocrinol 535: 111389.
-
Arora H, Zuttion MSSR, Nahar D, Lamb B, Hare JM, et al. (2019) Subcutaneous Leydig cells autograft: a promising strategy to increase serum testosterone. Stem Cells Transl Med 8(1): 58-65.
-
Wang Z, Tang T, Liu S, Chen Y (2020) Effect of bone marrow mesenchymal stem cells on ovarian and testicular function in ageing Sprague-Dawley rats induced by D-galactose. Cell Cycle 19(18): 2340-2350.
-
Sun D, Dong W, Jin B, Chen G, Cai B, et al. (2018) Mechanisms of Yangying capsules in Leydig cells apoptosis and testicular synthesis via promoting StAR expression. Biol Pharm Bull 41(9): 1401-1405.
-
Bi Z, Zhang W, Yan X (2022) Antiinflammatory and immunoregulatory effects of icariin and icaritin. Biomed Pharmacother 151: 113180.
-
Sun J, Xu W, Zheng S, Lv C, Lin J, et al. (2022) Icariin promotes mouse Leydig cell testosterone synthesis via the Esr1/Src/Akt/Creb/Sf-1 pathway. Toxicol Appl Pharmacol 441: 115969.
-
Shen F, Song Z, Xie P, Li L, Wang B, et al. (2021) Polygonatum sibiricum polysaccharide prevents depression-like behaviors by reducing oxidative stress, inflammation, and cellular and synaptic damage. J Ethnopharmacol 275:114164.
-
Han C, Zhu Y, Yang Z, Fu S, Zhang W, et al. (2020) Protective effect of Polygonatum sibiricum against cadmium-induced testicular injury in mice through inhibiting oxidative stress and mitochondria-mediated apoptosis. J Ethnopharmacol 261:113060.
-
Zhang Q, Yang C, Zhang M, Lu X, Cao W, et al. (2021) Protective effects of ginseng stem-leaf saponins on D-galactose-induced reproductive injury in male mice. Ageing (Albany NY) 13(6): 8916-8928.
-
Alanazi HH, Elfaki E (2023) The immunomodulatory role of Withania somnifera (L.) dunal in inflammatory diseases. Front Immunol 14: 1084757.
-
Li ZH, Lu JD, Li SJ, Chen HL, Su ZJ (2022) Generation of Leydig-like cells: approaches, characterization and challenges. Asian J Androl 24(4): 335-344.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’