Effects of Injecting GnRH 48 Hours after PGF2α on the Dynamic Follicular and Luteal Endocrine Cells in Post-Pubertal Holstein Heifers
Eighteen cycling Holstein heifers were allotted at random by weight and body condition score to one of two treatments to evaluate the effects of GnRH on luteal response when injected 48 hours (h) after the first injection in a 10 day interval between two injections of PGF2α . Heifers in the control group (n=9) received an injection of saline 48 h after the first injection of PGF2α; however, heifers in the µg) 48treatmenthafter group n=9) received an injection of GnRH (100 the first injection of PGF2α. Heifers were checked for estrus 3 times daily for 60 minutes each time. Blood samples were collected for analysis of progesterone on days 0 (first injection of PGF2α), 2(48 h after the first injection), 10(at the second injection of PGF2) and at day 17(7 days after the second injection of PGF2α). Plasma samples were analyzed for concentration of progesteronevia radioimmunoassay to evaluate luteal cell function. Concentrations of progesterone did not differ between the control heifers and treatment animals at any day of the study. However, heifers treated with GnRH showed a significant decline in concentration of progesterone from day 0 to day 2 in a luteal response to the injection; nevertheless, concentrations of progesterone increased significantly from day 2 to day 10. Thus, this data demonstrated that injecting GnRH 48 h after PGF2α either speeds up formation of new corpus luteum or prevents full regression of the corpus luteum present prior to the first injection of PGF2α.
Introduction
Prostaglandin F2α (PGF2α) is a hormone commonly used in numerous protocols designed to manipulate the follicular dynamic at the ovary due to its luteolytic activity. Prostaglandin is an endogenous uterine factor responsible for inducing regression of the corpus luteum (CL) in large domestic animals [1]. The ability of the CL to undergo regression in response to PGF2α is mostly dependent on the number of receptors in the bovine CL [2]. The CL is an endocrine gland that develops on the ovary after ovulation; its main function in reproduction of mammals is to secrete progesterone for maintenance of pregnancy. Progesterone exerts most of its effects by Open Access Journal of Veterinary Science & Research
closely regulating transcription of genes through certain receptors. In turn, the receptors help in the expression of genes by binding certain progesterone elements on DNA [3]. On the other hand, gonadotropin releasing hormone (GnRH) is also effectively used on research studies and estrous synchronization protocols to alter follicular growth dynamic [4]. Gonadotropin releasing hormone is a decapeptide released by the hypothalamus that induces ovulation and consequently increases the number of CL in cattle, as showed in a study conducted by Stevenson, et al. However, the ability of an injection of GnRH to induce ovulation is dependent on the stage of the estrous cycle [5]. In addition, GnRH has been previously demonstrated to alter follicular development, resulting in synchronization and emergence of new follicular waves [6, 7]. Ovulation normally occurs 24 to 30 h after the start of behavioral estrus. However, injecting GnRH at the time of breeding should result in a large surge of luteinizing hormone (LH), by inducing ovulation and consequently increasing chances of conception [8]. For cattle with a growing dominant follicle 10 mm in diameter, treatment with GnRH induces ovulation, with emergence of a new follicular wave approximately 2 days after treatment, but only when ovulation occurred [9]. Improvement of conception following GnRH treatment during estrus has been attributed to the prevention of ovulation failure or to reduced variation in the interval to ovulation [10]. Furthermore, many studies have shown the role of GnRH in advancing physiological activities responsible for luteal cell formation and function. Follicle size during ovulation has been associated as a good sign of fertile beef heifers. The growth, development and maturation of ovarian follicles are fundamental processes for high reproductive efficiency in farm animals. A fixed number of primordial follicles are established during fetal development with ovarian follicle growth taking 3-4 months period and categorized into gonadotropin independent and gonadotropin dependent stages. Gonadotropin dependent follicle growth in cattle occurs in waves with 2-3 waves per estrous cycle [11]. Several combinations of GnRH and PGF2α have been used to effectively synchronize estrus in cattle with acceptable pregnancy rates. A study conducted by Twagiramungu, et al. [12] revealed that administration of GnRH concurrently with PGF2α impairs total regression of the luteal tissue. This is in agreement with data previously reported by our laboratory. However, several studies have been conducted by extending or reducing the interval between the first and the second GnRH injections in reference to the PGF2α. Research trials have demonstrated the effectiveness of these protocols in improving pregnancy rates when PGF2α is injected 7 days after the first injection of GnRH [13, 14]. Nevertheless, a study conducted by a group of investigators showed that extending the interval between the first GnRH and the PGF2α does not reduce the variability in response to synchronization of ovulation in heifers [15].
However, others observed that extending the administration of the second GnRH to 48 h after the PGF2α improved pregnancy rate [16]. On the other hand, drastically decreasing the interval between the first GnRH and PGF2α may affect fertility. This is supported by findings indicating that the probability of pregnancy decreases substantially in dairy cows experiencing an incomplete CL regression [17]. The detrimental factor is the relatively high concentrations of progesterone secreted by luteal tissue as a consequence of an incomplete luteolysis which in turn, reduces the ability of endogenous estradiol to induce a pre-ovulatory surge of luteinizing hormone (LH) and ovulation [12, 18]. One study expressed that orderly progression of follicle– stimulating hormone (FSH) and LH release plays a crucial role in follicle maturation and ovulation as well as in the formation and maintenance of the CL [19]. Several studies have indicated that the lower efficacy of PGF2α in inducing regression of early CL may be related to differences in signal transduction due to differential expressions of genes associated with the PGF2α receptor at those two developmental stages. It has been suggested that the lack of luteolytic action by PGF2α in the developing bovine CL might be due to alterations in components of the signal transduction associated with the receptor by locally produced hormones [20]. Nonetheless, there is need for more studies in this line of research to develop new strategies and improve current synchronization protocols to achieve higher pregnancy rates. These strategies can only be fostered with better understanding of physiological mechanisms in control of follicle growth dynamic.
In a number of farm animals, PGF2α is recognized as the physiological luteolysin that is responsible for regression of the CL at the end of a non-fertile cycle [1]. Exogenous PGF2α causes regression to the bovine CL only between day 5 and day 16 after estrus. The lack of PGF2α response could be due to the deficiency in number of affinity of PGF2α receptors in the early CL [21]. Prostaglandin F2α released from the uterus in a pulsatile fashion is essential to induce regression of the CL in a cow. The CL has been recognized as site of PGF2α production. Overall results support the concept that the local release of PGF2α within the regressing CL amplifies the luteolytic action of PGF2α from the uterus.
Open Access Journal of Veterinary Science & Research
Materials and Methods
A study was conducted at the Dairy Production and Research Unit of Alcorn State University to evaluate the effects of GnRH on the dynamics of ovarian follicular and luteal development in post-pubertal Holstein heifers; and secondly, to evaluate the effects of GnRH on luteal response when injected 48 h after the first injection in a 10 day interval between two injections of PGF2α. Eighteen cycling Holstein heifers were allotted at random by weight, age and body condition score to one of two treatments. Heifers were conditioned to a body condition between 3 and 4 (BCS 1=Thin; BCS 5 = Obese) and checked for reproductive soundness before the trial. All heifers received two injections of PGF2α (25mg; i.m., Lutalyse; Pharmacia Upjohn Company) given 10 days apart. Heifers in the control group (n=9) received an injection of 5 cc saline i.m. 48 h after the first injection of PGF2α; however, heifers in the treatment group (n=9) received µang, iinjection.m., of GnRH (100 Cystorelin; Merial Limited,) 48 h after the first injection of PGF2α . Heifers were checked for estrus 3 times daily for 60 minutes each time. Blood samples were collected for analysis of progesterone on days 0 (first injection of PGF2α), 2 (48 h after the first injection), 10 (at the second injection of PGF2α) and at day 17 (7 days after the second injection of PGF2α). Plasma samples were analyzed for concentration of progesterone via radioimmunoassay to evaluate luteal cell function. Progesterone assays were performed using a commercial enzyme immunoassay kit provided by Oxford Biomedical Research (Oxford, Michigan). This is an enzyme-linked immunosorbent assay that operates on the basis of competition of solid- phases RIA system relying upon competitive binding between a radioactive and non-radioactive antigen for a fixed number of antibody sites coated to the assay tubes. The cross reactivity of the progesterone antiserum has been measured in various compounds. The percent cross- reactivity is expressed as the ratio of the concentration of progesterone the reacting compound concentration at 50% binding of the Progesterone Standard (ng/ml). The range of the progesterone assay used for this study was between 0 and 60 ng/ml. The assay displayed a sensitivity of 0.12 mg/ml and an average recovery rate of 97%. Average inters-and intra-assay coefficients of variability were 8.03 and 11.7 5% respectively. Data collected on concentration of progesterone in blood were analyzed using the GLM repeated measures Analysis SAS Institute, 1991. The correlation between concentrations of hormones at different days was also evaluated using the SAS CORR procedure. LSD was used to test differences among treatments at P>0.05.
Results and Discussion
Means and standard errors for body weight (Kg) of heifers in the control (382.3±36) and treatment group (383.2±31) were not significantly different. Similarly, no significant differences were observed in body condition score (BCS) between the control heifers (3.4±0.1) and heifers allocated to the treatment group (3.4±0.1; P>0.05). Several studies have demonstrated the effect of diet, body weight and body condition on reproductive patterns in cattle [22]. In this study, animals were kept on grass and pre-conditioned with supplemental grains to eliminate differences in body weight and body condition. Thus, no differences in body weight and body condition of heifers in the control and treatment group were observed. Consequently, these variables did not have an effect on the results reported in this study. Table 1 shows the means and standards errors for concentration of progesterone (ng/ml) on different days for heifers in the control and treatment groups. Concentration of progesterone did not differ between the control heifers and treatment animals at any day of the study (P>0.05).
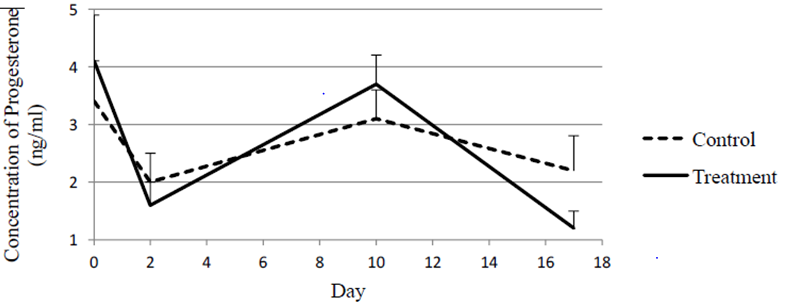
Figure 1 shows means and standard errors for concentrations of progesterone (ng/ml) at different days of the study within each treatment. Concentration of progesterone remained reasonably steady in the control group all across the study. No significant differences were observed in concentrations of progesterone between days in the control heifers. Progesterone was not affected by PGF2α in the control group animals. Nevertheless, heifers receiving an additional injection of GnRH two days after the first injection of PGF2a were administered, showed a significant decline in concentration of progesterone from day 0 to day 2 in a luteal response to the PGF2a injection. On the other hand, concentrations of progesterone increased significantly from day 2 to day 10. Furthermore, a significant decline was observed from day 10 to day 17.
The steady concentrations of progesterone observed in the control group of this study after receiving an injection of PGF2α are in contrast with findings reported by other investigators [2]. Nevertheless, Shipley, et al. Demonstrated that the number of receptor sites on the CL regulates its response to the luteolytic effect of PGF2α. On the other hand, the decline in progesterone concentrations experienced by heifers receiving GnRH two days after the first injection PGF2α seems to be an expression of a regressing CL. This group of treated heifers additionally experienced a significant increase in concentration of progesterone in response to GnRH. This effect can be attributed to either an accelerated formation Open Access Journal of Veterinary Science & Research
of a new CL [23] or due to preventing full regression of the CL present prior to the first injection of PGF2α [12].
very similar to that observed by Watts, et al. in dairy heifers treated with PGF2α on days 12 through 15 of the estrous cycle. Similarly, Harper, et al. [12] reported a failure of GnRH in altering the number of Holstein heifers expressing estrus during a hormonal protocol designed to synchronizeovulation.
The number of heifers showing estrus in the control and treated groups was not different. Estrus response of heifers in the control and treatment groups in this study is
| Variable | N* | Day 0 | Day 2 | Day 10 | Day 17 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 9 | 3.4±0.7a | 2.0±0.5a | 3.1±0.5a | 2.2±0.6a | ||||||||||||
| Treatment | 9 | 4.1±0.8a | 1.6±0.4a | 3.7±0.5a | 1.2±0.3a |
Table 1: Means and Standard Errors for Concentrations of Progesterone (ng/ml) at Different Days of the Study. N* = Number of Anim
Conclusion
Previously, our laboratory reported that injecting GnRH right after PGF2α alters the dynamics of follicular and luteal bovine cells. Thus, this data demonstrated that injecting GnRH 48 h after PGF2α either speeds up formation of new corpus luteum [23] or prevents full regression of the corpus luteum present prior to the first injection of PGF2α [12].
References
-
Jerome A, Shivastava N (2012) Prostaglandins Vis-à- vis bovine embryonic mortality: A review. Asian Pacific J of Reprod 1(3): 238.
-
Niswender G, Juengel J, Silva P, Rollyson M, McIntosh E (2000) Mechanisms controlling the function and life span of the corpus luteum. Physiol Rev 80(1): 1-29.
-
Jacobsen BM, Horwitz KB (2012) Progesterone receptors, their isoforms and progesterone regulated transcription. Mol Cell Endocrinol 357(1-2): 18-29.
-
Martinez MF, Adams GP, Bergfelt D, Kastelic JP, Mapletoft RJ (1999) Effect of LH or GnRH on the dominant follicle of the first follicular wave in heifers. Anim Reprod Sci 57(1-2): 23-33.
-
Perry GA, Perry BC, Krantz JH, Rodgers J (2011) Influence of inducing luteal regression before a modified fixed timed artificial insemination protocol in post-partum beef cows on pregnancy success. J Ani Sci 90(2): 489-494. Open Access Journal of Veterinary Science & Research
-
Clark BA, Rhodes FM, Burke CR, Morgan SR, Macmillan KL (1998) Manipulating patterns of ovarian follicle development in cattle with progesterone and gonadotrophin releasing hormone to produce oestrous cycles with two or three follicle waves. Proc NZ Soc Anim Prod 5: 85-87.
-
Marcos G Colazo, John P Kastelic, Julie A Small, Randy E Wilde, Douglas R Ward (2007) Resynchronization of estrus in beef cattle: Ovarian function, estrus and fertility following progestin treatment and treatments to synchronize ovarian follicular development and estrus. Can Vet J 48(1): 49-56.
-
Kaim M, Bloch A, Wolfenson D, Braw Tal R, Rosenberg M, et al. (2003) Effects of GnRH administered to cows at the onset of estrus on timing of ovulation, endocrine responses and conception. J Dairy Sci 86(6): 2012-2021.
-
Sa Filho OG, Patterson DJ, Vasconcelos JL (2011) Treatment to optimize the use of artificial insemination and reproductive efficiency in beef cattle under tropical environments. Vet Med Int 923053.
-
Gumen A, Keskin A, Yilmazbas Mecitoglu G, Karakaya E, Cevik S, et al. (2011) Effects of GnRH, PGF2α, and oxytocin treatments on conception rate at the time of artificial insemination on lactating dairy cows. Czech J Ani Sci 56(6): 279-283.
-
Crowe M, Mullen MP (2013) Regulation and differential secretion of gonadotropins during post partum recovery of reproductive function in beef and dairy cows, pp: 108-124.
-
Harper R, Bennett WA, Cuadra EJ, Vaughn CF, Watson CE, et al. (2008) Effects of GnRHα on the in dynamic combination of follicular with PGF 2and luteal endocrine cells in post-pubertal Holstein heifers. Livest Sci 117: 88.
-
Lamb GC, Stevenson JS, Kesler DJ, Garverick HA, Brown DR (2001) Inclusion of an intravaginal progesterone insert plus GnRH and prostaglandin F2alpha for ovulation control in postpartum suckled beef cows. J Anim Sci 79(9): 2253-2259.
-
Sa Filho OG, Patterson DJ, Vasconcelos JL (2009) Development of estrous synchronization protocols using melengestrol acetate in Bos indicus cattle. J Anim Sci 87(6): 1981-1990.
-
Rose CJ, Fuquay A, Moore S, Whisnant A, Williams S, et al. (2000) Evaluation of injection intervals in a modified ovulation-synchronization protocosl in dairy heifers. J Anim Sci, pp: 214.
-
Schmitt EP, Diaz T, Barros CM, De la Sota RL, Drost M, et al. (1996) Differential response of the luteal phase and fertility in cattle following ovulation of the first wave follicle with human chorionic gonadotropin or an agonist of Gonadotropin Releasing Hormone. J Anim Sci 74(5): 1074-1083.
-
Martins JPN, Policelli RK, Pursley JR (2009) Dynamicsαof luteolysis using two PGF2 analogs and subsequent differences in fertility. J Anim Sci 87: 268.
-
Hatler TB, Hayes SH, Ray DL, Reames PS, Silvia WJ (2008) Effect of subluteal concentrations of progesterone on luteinizing hormone and ovulation in lactating dairy cows. The Veterinary Journal 177(3): 360-368.
-
Menon KM, Munshi UM, Clouser CL, Nair AK (2004) Regulation of luteinizing hormone/human chorionic gonadotropin receptor expression: A Perspective. BioReprod 70(4): 861-866.
-
Goravanahally MP, Salem M, Yao J, Inskeep EK, Flores JA (2009) Differential gene expression in the bovine corpus luteum during transition from early phase to mid phase and its potential role in acquisition of luteolytic sensitivity to Prostaglandin F2 Alpha. Bio of Reprod 80(5): 980-988.
-
Parmar GA, Smith MF, Geary TW (2004) Ability of intravaginal progesterone inserts and melengestrol acetate to induce estrous cycles in postpartum beef cows. J Anim Sci 82(3): 695-704.
-
Cassady JM, Maddock TD, DiCostanzo A, Lamb GC (2014) Initial body condition score affects hormone and metabolic response to nutritional restriction and repletion in yearling postpubertal beef heifers. J Anim Sci 8 7(7): 2262-2273.
-
Mason MC, Copeland J, Cuadra EJ, Elsasser TH, Larson J, et al. (2014) Dynamics of progesterone, TNF-α and a metabolite of PGF2 in blood plasma of beef cows following embryo transfer. Vet Med Inter, pp: 8.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review