Melatonin Effect on Cryopreserved Sperm Cells of Crioulo Stallions
The freezing/thawing process of spermatozoa can cause cellular damage to the male gamete, decreasing the fertilization potential due to the increase in the production of reactive oxygen species (ROS). Melatonin is a potent endogenous antioxidant that protects the body against the damage caused by ROS. This study has evaluated different melatonin concentrations on the sperm viability of cryopreserved semen of Crioulo stallions. For that, three ejaculates were collected from five stallions diluted in a commercial extender followed by centrifugation and resuspension in a commercial freezing extender supplemented with 0; 1.25; 2.5. 5mM of Melatonin before the cryopreservation process. After thawing, the evaluation was performed assessing motility and flow cytometry evaluations: the plasma membrane integrity (PI), the integrity of the acrosomal membrane (FITC-PNA), mitochondrial membrane potential (JC1), and ROS generation (DCF-DA). Our results showed that sperm motility in the group without Melatonin and the 1.25mM group did not show the difference; however, the groups 2.5mM and 5mM presented a reduction in sperm motility. The 1.25 mM concentration was able to protect the plasma membrane during the cryopreservation process, in addition to showing a significant reduction in the production of ROS and increasing the percentage of sperm with integral acrosome. It can also be seen that high concentrations of Melatonin did not show beneficial effects. In conclusion, the addition of 1.25 mM of the Melatonin in Crioulo sperm cells showed to have a protective effect on the sperm cell during cryopreservation.
Introduction
Brazil has several species of domestic animals that were introduced by the colonizers. The Crioulo horse also known as the Spanish horse, originating from Andalusian and Jaca Navarra horses, brought from the Iberian Peninsula by settlers in the sixteenth century [1]. These, after a long process of natural selection, have characteristics adapted to specific environmental conditions, being known as “criollas” breeds [2]. After long period of adaptation the Crioulo horse is now considered a local adapted breed of the Pampas (a natural region between Uruguay, Argentina, Brazil, and Paraguay, located in South America) with a reputation for long-distance endurance linked to low basal metabolism [1].
In the past years, the equine industry has faced ascension in their production, generating thousands of direct and indirect jobs across the world [3, 4]. Horses started to be used not only for sports but also for recreation, therapy, and physical activities, raising interest in the equine industry [5, 6]. These factors have pushed the implementation in the use of reproductive biotechnologies in the horse industry, allowing the majority of the breed associations to approve the use of artificial insemination (AI) and embryo transfer (ET) on regular bases[7, 8]. Horses can be inseminated with fresh, cooled, or frozen/thawed semen, facilitating the dissemination of the genetic material, consequently, increasing the possibilities of choice for the stallions [7]. The cryopreservation of semen is an important tool used in the animal industry since it can reduce the spread of diseases, eliminating the geographical barriers and allowing for the storage of the semen for an indeterminate period, with reproductive [9], besides being able to provide the formation of germplasm banks of naturalized breeds. During the preparation of equine ejaculate for the cryopreservation process, it is necessary to remove the seminal plasma and, consequently, the predominant source of the antioxidant protection for the sperm cells [10, 11]. This process can cause damage to the sperm cells affecting the stallion fertility results [9, 12, 13, 14].
The process of freezing and thawing the sperm cells can reduce their fertilization potential due to an increase in the production of reactive oxygen species (ROS) caused by the process of lipid peroxidation in the sperm membranes [15]. In addition to that, the process of cryopreservation and the generation of ROS can also cause DNA fragmentation resulting in decreased fertility [15, 16]. However, the physiological concentration of ROS is needed to increase the fluidity of the membranes, preserving the fertilization capacity of sperm cells, and also has some beneficial effects on sperm hyperactivation, capacitation, and acrosomal reaction [17, 18].
Therefore, lower concentrations of ROS play an essential role in sperm physiology, while high concentrations can cause damage. Thus, protection mechanisms are required to limit the production of ROS to maintain normal cellular functions [17, 18].
Several studies have investigated the effect of different antioxidants in the supplementation of stallions to improve the quality of sperm quality [19, 20]. The use of primary antioxidants and fatty acids (L-carnitine, selenium, vitamin E, omega-3, and omega-6) by oral supplementation had a positive effect on sperm kinetics, increasing the plasma and acrosomal membrane integrity, due to the increase in the concentrations on poly-unsaturated fatty acids combined with the decrease of lipid peroxidation by the antioxidants [21]. However, it requires constant administration of this supplementation for an extended period. Thus, the use of those antioxidants directly in the ejaculate can help to minimize the losses in sperm quality during cryopreservation and storage of spermatozoa [17, 22].
Melatonin is a hormone produced by the pineal gland, which maintains circadian rhythm. It is also a potent endogenous antioxidant that helps to protect the body against damage from free radicals (ROS) [23, 24, 25]. González- Arto, et al. [26] identified the presence of MT1 and MT2 Melatonin receptors in spermatozoa of different domestic and wild species, regardless of their seasonality. The presence of these receptors on spermatozoa suggests that Melatonin supplementation may directly influence sperm function, exerting some regulation of the antioxidant defense mechanism [26]. Studies have already demonstrated the effect of Melatonin in the spermatozoa of humans and animals, but the regulatory mechanism that it exerts is still unknown [22, 25].
Due to its antioxidant’s properties, Melatonin could be added to the equine semen extender minimizing the sperm losses in cryopreserved cells by capturing the ROS produced during the process reducing the membrane damage. Studies have been conducted in other animal species, demonstrating it beneficial effect of the Melatonine use in sperm cells [27, 28]. However not much attention was giving in the use of Melatonin in the equine semen. In addition to that the existance of breed variation in the efficiency of cryopreservation protocols in horses [29]. Therefore the optimization of protocols for the cryopreservation of sperm cells in the Crioulo horses might be beneficial to the breed conservation and dissemination. In this context, the objective of this study was to evaluate different concentrations of Melatonin on the sperm viability characteristics in cryopreserved semen of Crioulo stallions.
Materials and Methods
Animals, Collection and Semen Processing
The experiment was conducted in accordance with the use of animals for research. A total of five Crioulo breed stallions with different genetic backgrounds, the average age of 14.2+2.9 years, and the bodyweight of 350kg to 450kg were used in this study. The stallions were from different owners; however, they were housed in the same breeding center for the whole year. Ejaculates were collected and examined regularly for breeding purposes during the reproductive season (two to three times per week), and animals had proven fertility (˃50% pregnancy rate using fresh semen in the breeding season).
Experimental Design
For the experiment, three ejaculates were used per stallion. Semen was collected using an artificial vagina (Botupharma, Botucatu, SP, Brazil) warmed up to 50ºC, equipped with a nylon filter to allow the collection of gel-free semen. Ejaculates were evaluated for total volume, sperm motility, and concentration. All analyses were performed according to the Brazilian School of Animal Reproduction [30].
The filtered ejaculates were then diluted in a skim milk- based extender (FG Mix Plus, FG Veterinária, Sorocaba, SP, Brazil) to obtain a final sperm concentration of approximately 50 x 106 spermatozoa ml-1, and centrifuged at 600g for 12 min. The supernatant was discarded, and the sperm pellet was re-suspended in an egg yolk-based commercial extender (Botucrio, Botupharma, Botucatu, SP, Brazil) to a final concentration of 200 x 106 spermatozoa ml-1 preheated at 37°C to avoid cold shock.
The semen was divided into four aliquots with different concentrations of Melatonin (Melatonin powder ≥98% TLC, Sigma (Merck KGaA, Darmstadt, Germany) catalog number M5250): 0, 1.25,2.5, and 5mM pre-diluted in isopropanol. The samples were packaged in 0.5 ml straws, sealed and cooled to 5°C for 20 min (≤3°C / min). After cooling, the semen was frozen at a rate of 20 to 50°C per minute. For this, the straws were placed on a floating shelf, 6 cm above the liquid nitrogen, for 20 min. Then, they were immersed in liquid nitrogen and stored for at least seven days.
For the evaluation of the sperm samples, the straws were thawed at 37˚C for 30 seconds. After thawing, the contents of each straw were diluted in 1 ml the Tyrode´s albumin lactate and pyruvate (TALP) at 37ºC. Motility was evaluated in a phase-contrast microscope, on an objective lens of 200x at
37˚C for three times by the same observer. Flow cytometry was used to evaluate four sperm variables (integrities of the plasma membrane and acrosome, mitochondrial membrane potential, and intracellular concentrations of reactive oxygen 198 species). In each assay, sperm concentration was adjusted to a final concentration of 60 × 106 spermatozoa/ mL with Tyrode´s albumin lactate and pyruvate (TALP). Three replicates per concentration were examined. The samples were analyzed using the flow cytometer BD AccuriC6 (Becton & Dickson, Santiago, Chile) equipped with standard optics and an air-cooled argon laser operated at 488 nm excitation and 20 mW, and three-light filters (FL-1: 533±30 nm; FL-2: 585±40 nm; FL-3: 675±25 nm). Forward scatter (FSC), and side scatter (SSC) were used to gate the sperm population and to exclude debris. A total of 10,000 sperm events at 600cells/sec were acquired from each measurement. The acquisition was performed with a sample aspiration speed of 14μL/min. The data were acquired on a logarithmic scale. When required, the spectral overlap of each particular staining was compensated.
Integrities of plasma and acrosome membranes were assessed with Propidium Iodide (PI; Sigma-Aldrich Co., Saint Louis Missouri, USA; 10μg/mL; BP: 610/20 nm) and Pisum sativum agglutinin conjugated to fluorescein isothiocyanate (FITC-PSA; Sigma-Aldrich Co., Saint Louis Missouri, USA; 1.25 μg/ mL; BP: 530/30 nm), respectively. After stained, sperm samples were and incubated at 37 °C for 10 min in a darkened area of the laboratory and then analyzed by flow cytometry. Mitochondrial membrane potential was evaluated by 5,5′,6,6′-tetra-chloro-1,1′,3,3′ tetraethylbenzimidazolyl carbocyanine iodide (JC1: 10 μg/mL; BP: 585/40 nm). The samples were incubated in a darkened area at 37 °C for 10 min before the time of flow cytometric analysis. Intracellular concentrations of hydrogen peroxide were assessed with 2′,7′-dichlorodihydrofluorescein diacetate (DCFDA; 6.67 μg/ mL; BP: 530/30 nm), and PI (PI; Sigma-Aldrich Co., Saint Louis Missouri, USA; 10 μg/mL; BP: 610/20 nm). Sperm samples were simultaneously stained with DCFDA and PI, and incubated at 37 °C for 10 min in the darkened area of the laboratory, and following analyzed by flow cytometry. The analysis was performed only on PI negative sperm, that is, with an intact plasma membrane. The result of the analysis of ROS generation was expressed as a percentage of sperm with low oxygen reactive species.
Statistical Analysis
The data was evaluated using SAS Institute Inc., Cary, NC, EUA. The ANOVA test was performed. All the parameters evaluated presented a normal distribution; therefore, the Tukey statistical test was used to compare the mean; the significance level was considered if P<0.05. The data presented as a mean percentage.
Results
When evaluating the characteristics of the newly collected ejaculates, an average of the volume is 62.9 + 18.10 ml per collection (min = 30ml, max = 90ml Males S3 and S4 presented higher volume of ejaculate than male S3 (P <0.04) (Table 1). The average sperm motility of the stallions at the time of collection was 75.3%, with a concentration of 131.5x106+47x106 spermatozoa per ml, with total sperm concentration ranging from 3.7x109 to 13.6x109 (Table 1). Three different concentrations of Melatonin (1.25 mM, 2.5 mM e 5mM) were used in the freezing semen extender, and parameters of sperm viability were measured. The greatest reductions in sperm motility were observed in groups 2.5 and 5 mM (42.6±8.83% and 33.0±11.61%) when compared to the control group (52.0±9.41%) and 1.25 mM (50.0±8.45%) (P<0.0007).
| S1 | S2 | S3 | S4 | S5 | |
|---|---|---|---|---|---|
| Volume (ml) | 63 ± 15.27AB | 50 ± 10AB | 81 ± 7.63A | 76 ± 11.54A | 43 ± 11.54B |
| [ ] x 106/ml | 191 ± 30 | 108±10 | 169 ± 21 | 101 ± 33 | 88 ± 26 |
| [ ] x 109 total | 11.8 | 5.3 | 13.6 | 7.6 | 3.7 |
| Fresh motility (%) | 73 ± 5.77 | 80 ± 11.54 | 83 ± 5.77 | 70 ± 0 | 70 ± 0 |
Table 1: Average of the collections of the five 12 individual stallions, volume and sperm concentration per ml. The total con- ce
Table 1: Average of the collections of the five 12 individual stallions, volume and sperm concentration per ml. The total con- centration of the ejaculate and the motility of the spermatozoa at the time of collection.[ ] (concentration); S1: (stallion 1); S2: (stallion 2); S3: (stallion 3); S4: (stallion 4); S5: (stallion 5). The presentation data are the means ± SD. Letters superscript in the same line represent significant difference (P <0.05) About the plasma membrane integrity evaluation, the group with 1.25mM of Melatonin presented a higher percentage of sperm cells with intact membranes when compared with the Control group and the 5.0mM of Melatonin (P<0.05) (Figure 1A). A significant difference was observed in the acrosomal membrane integrity between the Control group and the treated groups, and no difference was observed for this parameter among the different Melatonin doses (Figure 1B). There was no effect of Melatonin on mitochondrial activity (Figure 1C). For the ROS production, it was observed a protective effect among the treated group with 1.25mM of Melatonin when compared with 5mM (P<0.05) (Figure 1D).
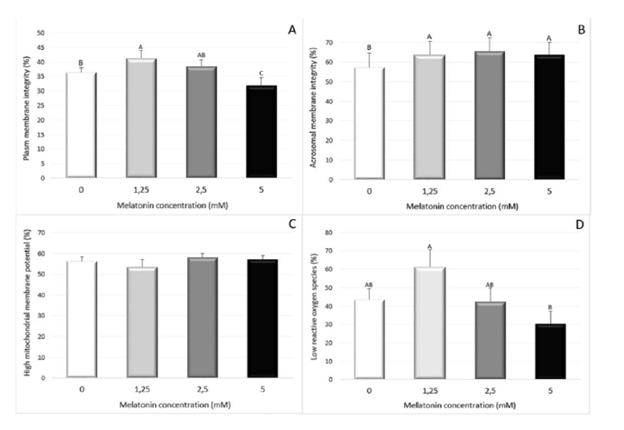
Figure 1: The effect of different melatonin doses (0, 1.25, 2.25, 5 mM) in the sperm cell parameters. (A) in the Plasma Membrane Integrity, (B) Acrossomal Membrane Integrity, (C) Mitocondrial Membrane Potential and (D) Reactive Oxigen Species production. Means denoted by a different letter indicate significant differences between treatments (P<32 0.05).
Discussion
The process of freezing/thawing semen generates reactive oxygen species (ROS) that can lead to a reduction in spermatic quality [31]. Exogenous ROS can induce DNA fragmentation, impairing sperm viability; consequently, the addition of antioxidants to the spermatozoa could reduce the loss of integrity, minimizing the deleterious effects of the oxidative stress [15]. In the present study, we investigated the use of different concentrations of Melatonin in the freezing extender in the cryopreserved spermatozoa of Crioulo stallions concerning parameters of sperm viability.
The collected semen, gel-free, must be diluted in skim milk or casein extender in a dilution of at least 1:1 (vol/ vol) [11]. In our study, as a force of 600g per 12 minutes was used for centrifugation, the sperm concentration before centrifugation was maintained between 30-50 x 106 sptz /ml in order to reduce sperm loss during the removal of seminal plasma and avoiding the formation of highly compressed pellets and challenging to re-suspend [32].
The freezing protocol used was described [9]. For the assessment of melatonin efficiency, different concentrations were chosen according to previous studies that evaluated the relationship between Melatonin and the viability of sperm of different species [22, 23, 25, 33].
The dilution of Melatonin with dimethylsulfoxide (DMSO) carried out by Izadnapah and collaborators were found not to affect the parameters of sperm viability. However, in our study, the cryopreservation process associated with the use of ethanol to dilute Melatonin had no beneficial effects on the sperm viability of the stallion. Martín-Hidalgo, et al. [34] has described that the Melatonin dose was chosen after performing an ethanol dose-response curve to select a concentration that did not compromise sperm characteristics using different concentrations of ethanol (0.001%, 0.003%, 0.01%, 0.03%, 0.3%, and 1%), identifying a significant decrease in the total percentage of mobile spermatozoa with the increase in ethanol concentration, where the dose of 0.003% did not affect this parameter. In our study, the highest ethanol concentration was 1% in the highest melatonin concentration treatment (5mM). Some parameters evaluated had lower responses in the 5mM group, which may have been due to an association between the highest concentrations of Melatonin and also of ethanol. Possibly the vehicle used (ethanol) in the melatonin dilution may have altered the cryopreservation extender, impairing sperm quality concomitantly with an increase of melatonin concentrations.
However, in a pre-experiment (unpublished data), we did not observe a difference in the motility of fresh semen samples diluted in a commercial diluter with and without the addition of 1% ethanol and evaluated for 20min. Melatonin’s antioxidant activity helps prevent the testicular damage caused by toxic environments and testicular inflammation; however, it seems to promote death by apoptosis in some cancerous cells. That is, Melatonin contributes both to protection and apoptosis in different cell types [35]. The anti-apoptosis and antioxidative melatonin effects can act by reducing oxidative stress in the sperm and consequently increase the quality of a bovine embryo resulting from its fertilization [26]. In their study, the melatonin dose of 1mM was the dose that contributed most to obtain better results. Just like other hormones involved in reproductive regulation, abnormal levels of Melatonin may also cause infertility [35].
The possible role of Melatonin in protecting the spermatozoa against free radical damage has not yet been fully elucidated [23]. Bornman, et al. [36] demonstrated that there is no difference in melatonin concentration between the seminal plasma of patients with good and impaired semen motility, concluding that Melatonin in seminal plasma has no important role in sperm motility.
Pang, et al. [26] observed that the melatonin supplementation did not affect the kinetics of bull spermatozoa after the freezing/thawing process, just as they did not observe a significant statistical difference in the proportion of total mobile sperm, as well as any other kinetic parameter was observed between the treated groups and the control.
Our studies show that there was no effect of adding 1.25mM of Melatonin on sperm motility after thawing; however, there was a decrease in motility in the highest concentrations (2.5 and 5mM), indicating that for horses, doses greater than 2.5mM may have undesirable effects.
The melatonin supplementation in the freezing extender was capable of neutralizing the adverse effects of the freezing/thawing process on the motility, viability, normal morphology, and plasma membrane integrity in bull spermatozoa [31]. For them, the inclusion of 2 or 3 mM of Melatonin in the semen extender before the freezing/thawing process can improve the efficiency of artificial insemination in bovine animals. In the present study, concerning the plasma membrane integrity parameter, it was identified that the 1.25mM presented a higher percentage of non-damaged sperm cells when compared with the control group and the 5mM of Melatonin (P< 0.001). These results agree with the findings of Pang, et al. [26], who described that 1mM of Melatonin improved both plasma membrane integrity and acrosomal integrity when compared to the control group.
When evaluating the acrosomal integrity, it was observed
that all the Melatonin treated groups protected the acrosomal sperm cells. In sheep, the melaton in addition to the extender media, before cryopreservation, at the concentration of 100 pM, helped to protect the acrosomal membrane integrity, when compared with the other treatments used. In the concentrations used (1 mM; 100 μM; 100 nM, and 100 pM), the authors observed that regardless of the concentration of Melatonin added to the extender media, the percentage of cells with intact acrosomes after thawing was superior to the control treatment, demonstrating the melatonin antioxidant potential allowing more excellent protection of the acrosomal membrane against oxidative and mechanical damage caused by cryopreservation [33].
A study has revealed that MT1 and MT2 melatonin receptors are also present in human sperm, and the supplementation of Melatonin (0.1mM) as effectively diminished the presence of free radicals, enhancing the reduction of damage caused by ROS, and has not yet presented cytotoxicity to the frozen/thawed human sperm [37]. In corroboration, a further study has demonstrated that in humans, the addition of 1.5mM and 3mM of Melatonin to the freezing extender obtained positive results when compared the motility parameters and the selected dose as optimal was 3mM, it obtained the highest results on the sperm quality including a decrease of the intracellular levels of H2O2 and O2 in the sperm after the cryopreservation [25]. The results obtained with the addition of Melatonin in fresh equine semen under different concentrations (1mM, 1.5mM, and 2mM) demonstrated that in all treatments, Melatonin had positive effects on sperm viability during the liquid preservation of semen for 48 hours [22]. They also concluded that the 1.5mM melatonin treatment could be used in the semen extender to improve the in vitro quality of the semen of Caspian stallions in cooling conditions. In our study, the cryopreserved sperm cells with 1.25mM of Melatonin showed to have an efficient reduction in the production of ROS.
Thus, our study demonstrated that as for the cryopreserved semen in other species and the equine semen cooled, the addition of Melatonin is beneficial for sperm during the cryopreservation process, and that, besides, the concentration of 1.25 mM the best results[38, 39, 40].
Conclusion
In conclusion, in our experimental conditions, the addition of 1.25mM of the Melatonin to stallions freezing extender showed to have a beneficial effect on sperm viability parameters. Besides, it can also be seen that the concentration of 5mM did not show beneficial effects.
Conflict of Interest: None of the authors have any conflict of interest to declare.
Acknowledgment
We would like to thank the breeding center Viva Equi for semen collection and freezing, to Embrapa Sunos eAves/ Concordia-SC for making the laboratory available for the sample evaluation and to Zerlotti Genetics for translating the text into English.
References
-
Brazilian Association of Crioulo Horse Breeders (2014) Breeders manual. Pelotas, pp: 7-9.
-
Mariante A da S, Albuquerque M do SM, Egito AA, McManus C, Lopes MA, et al. (2009) Present status of the conservation of livestock genetic resources in Brazil. Livest Sci 120: 204-212.
-
Shouksmith G, Hesketh B (1986) Changing horses in mid-stream: Job and life satisfactions for veterinarians. N Z Vet J 34(9): 141-144.
-
Centre TH (2004) A report of research on the horse industry in Great Britain. British Horse Industry Confederation, Derfra.
-
Soares JCM, Zanella R, Bondan C, Alves LP, de Lima MR, et al. (2011) Biochemical and Antioxidant Changes in Plasma, Serum, and Erythrocytes of Horses before and after a Jumping Competition. J Equine Vet Sci 31(7): 357- 360.
-
Lima R, Cintra A (2016) Study of the complex of the gronegócio horse cepea / ESALQ / USP.
-
Aurich C (2008) Recent advances in cooled-semen technology. Anim Reprod Sci 107(4): 268-275.
-
Galli C, Colleoni S, Duchi R, Lagutina I, Lazzari G (2013) Equine assisted reproduction and embryo technologies. Anim Reprod 10(3): 334-343.
-
McCue PM (2014) Breeding Soundness Evaluation of the Stallion, in: Equine Reproductive Procedures. John Wiley & Sons, Inc, Hoboken, NJ, USA, pp: 319-324.
-
Ramires Neto C, Monteiro GA, Soares RF, Pedrazzi C, Dell’aqua JA, et al. (2013) New seminal plasma removal method for freezing stallion semen. Theriogenology 79(7): 1120-1123.e1.
-
Alvarenga MA, Papa FO, Ramires Neto C (2016) Advances in Stallion Semen Cryopreservation. Vet. Clin. North Am Equine Pract 32(3): 521-530.
-
Hartwig FP, Lisboa FP, Hartwig FP, Monteiro GA, Maziero RRD, et al. (2014) Use of cholesterol-loaded cyclodextrin: an alternative for bad cooler stallions. Theriogenology 81(2): 340-346.
-
Carvalho CPT, Lançoni R, Santos GC, Berber RCA (2015) Main Advances in Biotechnologies Used in Artificial Insemination of Horses: A Major Advances in Biotechnology Review Used on Artifial Insemination of Horses : A Review. Sci Electron Arch 8(3): 72-78.
-
Stock KF, Jönsson L, Ricard A, Mark T (2016) Genomic applications in horse breeding. Anim Front 6(1): 45-52.
-
Baumber J, Ball BA, Linfor JJ, Meyers SA (2003) Reactive Oxygen Species and Cryopreservation Promote DNA Fragmentation in Equine Spermatozoa. J Androl 24: 621- 628.
-
Aitken J, Fisher H (1994) Reactive oxygen species generation and human spermatozoa: The balance of benefit and risk. BioEssays 16(4): 259-267.
-
Cocchia N, Pasolini MP, Mancini R, Petrazzuolo O, Cristofaro I, et al. (2011) Effect of sod (superoxide dismutase) protein supplementation in semen extenders on motility, viability, acrosome status and ERK (extracellular signal-regulated kinase) protein phosphorylation of chilled stallion spermatozoa. Theriogenology 75(7): 1201-1210.
-
Dutta S, Majzoub A, Agarwal A (2019) Oxidative stress and sperm function: A systematic review on evaluation and management. Arab J Urol 17(2): 87-97.
-
Deichsel K, Palm F, Koblischke P, Budik S, Aurich C (2008) Effect of a dietary antioxidant supplementation on semen quality in pony stallions. Theriogenology 69(8): 940-945.
-
Contri A, De Amicis I, Molinari A, Faustini M, Gramenzi A, et al. (2011) Effect of dietary antioxidant supplementation on fresh semen quality in stallion. Theriogenology 75(7): 1319-1326.
-
Freitas ML, Bouéres CS, Pignataro TA, Gonçalves de Oliveira FJ, de Oliveira Viu MA, et al. (2016) Quality of Fresh, Cooled, and Frozen Semen From Stallions Supplemented with Antioxidants and Fatty Acids. J Equine Vet Sci 46: 1-6.
-
Izadpanah G, Zare-Shahneh A, Zhandi M, Yousefian I, Emamverdi M (2015) Melatonin Has a Beneficial Effect on Stallion Sperm Quality in Cool Condition. J Equine Vet Sci 35(7): 555-559.
-
Du Plessis SS, Hagenaar K, Lampiao F (2010) The in vitro effects of Melatonin on human sperm function and its scavenging activities on NO and ROS. Andrologia 42(2): 112-116.
-
González-Arto M, Vicente-Carrillo A, Martínez-Pastor F, Fernández-Alegre E, Roca J, et al. (2016) Melatonin receptors MT1 and MT2 are expressed in spermatozoa from several seasonal and nonseasonal breeder species. Theriogenology 86(8): 1958-1968.
-
Najafi A, Adutwum E, Yari A, Salehi E, Mikaeili S, et al. (2018) Melatonin affects membrane integrity, intracellular reactive oxygen species, caspase3 activity and AKT phosphorylation in frozen thawed human sperm. Cell Tissue Res. 372(1): 149-159.
-
Pang YW, Sun YQ, Jiang XL, Huang ZQ, Zhao SJ, et al. (2016) Protective effects of Melatonin on bovine sperm characteristics and subsequent in vitro embryo development. Mol Reprod Dev 83(11): 993-1002.
-
Monllor F, Espino J, Marchena AM, Ortiz Á, Lozano G, et al. (2017) Melatonin diminishes oxidative damage in sperm cells, improving assisted reproductive techniques. Turkish J Biol 41(6): 881-889.
-
Sharbatoghli M, Valojerdi MR, Bahadori MH, Yazdi RS, Ghaleno LR (2015) The relationship between seminal melatonin with sperm parameters, DNA fragmentation and nuclear maturity in intra-cytoplasmic sperm injection candidates. Cell J 17(3): 547-553.
-
Kuhl J, Tichy A, Aurich C, Aurich J (2020) Efficiency of Semen Cryopreservation in Stallions. Animals 10(6): 1033.
-
CBRA (1998) Manual for andrological examination and evaluation of animal semen. Brazilian College of Animal Reproduction, Belo Horizonte.
-
Ashrafi I, Kohram H, Ardabili FF (2013) Antioxidative effects of Melatonin on kinetics, microscopic and oxidative parameters of cryopreserved bull spermatozoa. Anim Reprod Sci 139(4): 25-30.
-
Larzul C, Delaunay I, Schwob S, Mercat MJ (2008) Genetic parameters of the main porcine congenital anomalies. 40th Rech Days. Pork. Paris Fr 40: 141-142.
-
Souza WL, Moraes EA, Costa JMS, Sousa PHF, Lopes Junior ES, et al. (2016) Effect of different concentrations of melatonin to ram spermatozoa on oxidative stress after cryopreservation. Pesq Vet Bras 36(7): 657-664.
-
Martín-Hidalgo D, Barón FJ, Bragado MJ, Carmona P, Robina A, et al. (2011) The effect of melatonin on the quality of extended boar semen after long-term storage at 17°C. Theriogenology 75(8): 1550-1560.
-
Li C, Zhou X (2015) Melatonin and male reproduction. Clin Chim Acta 446: 175-180.
-
Bornman MS, Oosthuizen JM, Barnard HC, Schulenburg, GW, Boomker D, et al. (1989) Melatonin and sperm motility. Andrologia 21(5): 483-485.
-
Deng SL, Sun TC, Yu K, Wang ZP, Zhang BL, et al. (2017) Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic Biol Med 113: 347-354.
-
Da Silva Z, De Souza AP, Pandolfi JRC, Da Fonseca FN, Da Veiga Lima-Rosa, et al. (2018) Comparison between electroporation and polyfection in pig sperm: Efficiency and cell viability implications. Zygote 26(4): 286-293.
-
Karimfar M, Niazvand F, Haghani K, Ghafourian S, Shirazi R, et al. (2015) The protective effects of Melatonin against cryopreservation-induced oxidative stress in human sperm. Int J Immunopathol Pharmacol 28(1): 69- 76.
-
Lee JA, Spidlen J, Boyce K, Cai J, Crosbie N, et al. (2008) MIFlowCyt: the minimum information about a Flow Cytometry Experiment. Cytometry. Part A: the journal of the International Society for Analytical Cytology 73(10): 926-930.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review