Could Antimicrobial Resistance Prove to Be Both a Threat and an Opportunity for us?
Introduction: Diversification of viral and bacterial strains, climate and environmental changes, population mobility, uneven population density, different sanitary conditions and medical care, changes in animal habitats, and increasing antibiotic resistance contribute to increased pressure on the host-pathogen-environment system. Materials: Resistance exists in nature even to drugs that have not yet been developed. Will using the effects of probiotic strains, which simultaneously serve as 'deliverers' of proteinaceous antibiotic-like substances, allow us to remodel the interactions between the components of the holobiont?” Results: Probiotic strains can be used to restore the composition of the microbiota, bacteriocin-induced inhibition of other organisms, limit the spread of non-communicable diseases (eczema, allergy, or asthma), suppress their manifestations, in strategies for cancer protection and prevention, and treatment of infectious diseases. Conclusion: The use of probiotic strains increases competition for adhesion and nutrients with those with pathogenic potential. In sufficient quantities, they create a maximally hostile environment for colonization of pathogens and limit invasion, overgrowth of the latter, and even a return to the orbit of commensals. Unlike broad-spectrum antibiotics, which do not distinguish between commensals and pathogens and can reduce the microbiota up to 30%, probiotic strains inhibit or inactivate other organisms without adversely affecting the beneficial flora. To achieve our goals, probiotics must stop being food supplements and meet the safety and efficacy characteristics set in their preparation.
Introduction
Dynamics in habits, lifestyles, migration, and the cultivation of domestic animals for food over the centuries have allowed the jump and stabilization of many disease- causing agents in the human population. Diversification of viral and bacterial strains, climate and environmental changes, population mobility, uneven population density, different sanitary conditions and medical care, changes in animal habitats, and increasing antibiotic resistance contribute to increased pressure on the host-pathogen- environment system.
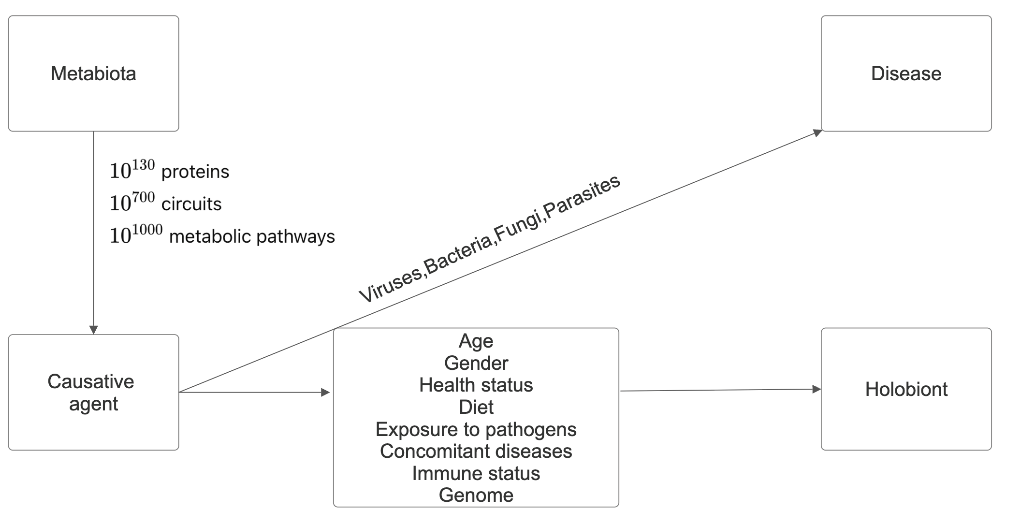
The genetic material of all organisms in a given environment - water, soil, the surrounding animal and plant world, and the microbiota of the individuals inhabiting the environment form the metagenome. Logically, the carriers of this genetic material are called metabiota. Microorganisms represent the majority of the planet’s biodiversity. The changes that all organisms have undergone during the approximately four billion years since the emergence of life on Earth have led to the evolutionary formation of 10130 protein molecules, 10700 signaling cascades, and 101000 metabolic pathways in nature.
Organisms live in a variety of conditions, and many of them cannot be cultivated using standard techniques. Metagenomics gives us information about their diversity and functional potential. The number of genes carried by the metagenome is probably comparable to the number 10130. Between organisms in the metabiota are observed positive - a) one-way (commensalism) and b) two-way - cooperation, mutualism and syntrophy, and negative interactions - competition, deception and parasitism, which participate in their biology, ecology and evolution [1]. The basis of the interactions is the sharing of a large set of molecules - enzymes, QS molecules, siderophores, and others [2]. Regardless of the direction of interactions, a specific aspect of the metabiota is its tendency towards equilibrium or eubiosis. Studying them will help us understand the course of infectious, inflammatory, and non-communicable diseases. The evolved defense mechanisms - virulence factors, signaling molecules redirecting metabolic pathways, and interactions ensuring the survival of microorganisms will allow us to shed light on how they can remain unnoticed by our immune system and the reasons for the different course of the same disease in patients.
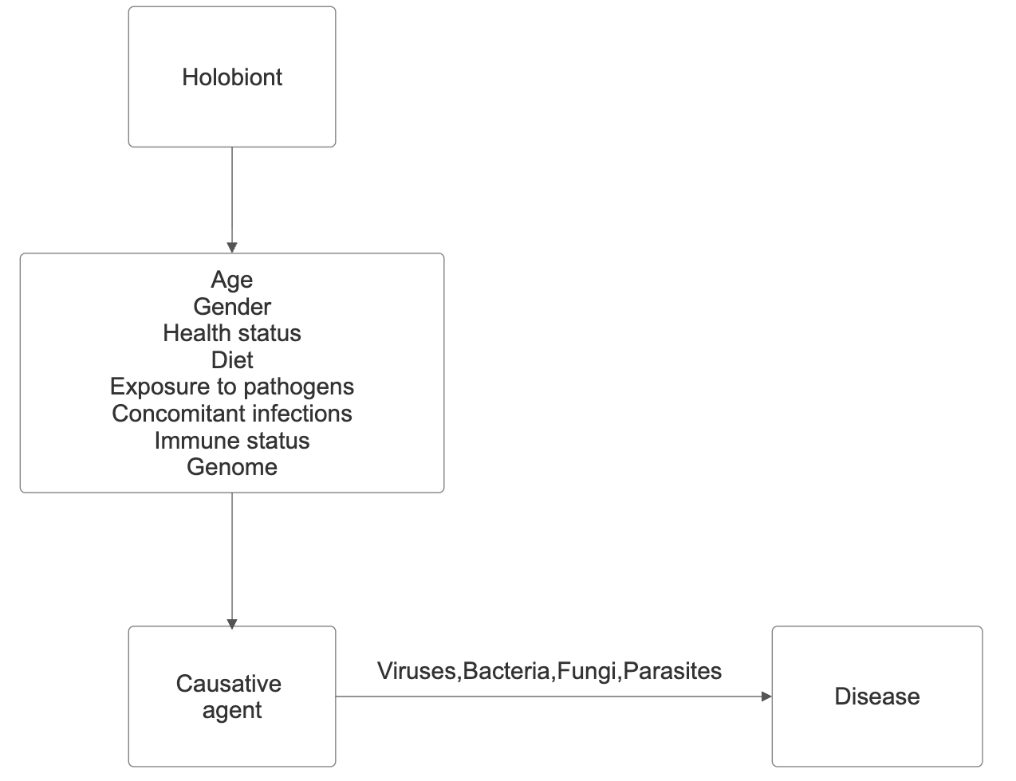
The relationships between the host and its associated microbial genomes shape the various human phenotypes, and the symbiosis between species underlies the concept of the holobiont. It was formulated by Margulis L [3] in 1991, and currently, the holobiont is considered a single entity between the host and the taxonomically and ecologically associated microbial communities or microbiota. It is characterized by synergistic to antagonistic interactions. In 2007, Zilber- Rosenberg I, et al. [4] introduced the term hologenome, representing the sum of the host genome and the genomes of its associated communities, or the collective genome of the holobiont. In 2007, Zilber-Rosenberg I, et al. [4] introduced the term hologenome, which refers to the sum of the host genome and the genomes of its associated communities or the collective genome of the holobiont. The human genome contains about 20000 genes, and our hologenome contains >33 million genes [5, 6, 7, 8, 9, 10]. The vitality of communities in it, according to the “Black Queen” hypothesis, is because in natural communities, evolutionarily positive interactions in complex habitats are strengthened through gene loss, leading to dependencies between microorganisms [11, 12]. Over time, interactions become so complex that individual strains cannot prosper on their own. The theory is that dependencies between microorganisms, especially those of a nutritional nature, enhance their vitality and strengthen their bonds against competitors, cheaters, and stress.
Metabolic dependencies are fundamental drivers of species survival in nature [13]. The sharing of metabolites in cross-feeding is crucial for positive interactions, according to D’Sousa. Microorganisms use two methods. The first is by sharing hydrolytic extracellular enzymes such as invertases, lipases, and proteases, which break down the necessary substrates from the common pool of molecules. In the second, we observe cross-feeding, in which one microorganism/ donor converts a primary substrate into a product that is subsequently used by another/acceptor [14].
The human microbiota, or host-associated microbial communities, is involved in the pathogenesis of disease. It is a natural community in which natural selection manages its dynamics. Its evolutionary selection over the centuries has contributed to human physiology, metabolism, and immunity. It consists of four kingdoms: archaea, viruses, bacteria, fungi, and, in some populations, parasites. Humans are host to over 100 trillion microbes, most of which are in the intestines and actively participate in metabolic, protective, and trophic functions. The metabolic activity of the intestinal microbiota (predominant) is expressed in the regulation of positive or negative homeostasis, the formation of an anaerobic environment, the suppression of the virulence of pathogens, and the induction of immune tolerance. At its base are the products of the metabolic pathways of tryptophan (Trp), histidine (His), and phenylalanine (Phe). The protective one is expressed in the production of bacteriocins inhibiting the growth or survival of the pathogen, and the trophic one in the mobilization of bacteriophages that attack specific bacterial strains, minimizing their impact on the commensal component. Changes in the microbiota can trigger the onset of many non-communicable (chronic inflammatory and metabolic) diseases, with the intestinal component serving as a biomarker of disease risk.
The increasing rate of antimicrobial resistance (AMR) represents one of the ten greatest threats to public health, according to the WHO [15]. It occurs when viruses, bacteria, fungi, and parasites become resistant to antimicrobial treatments to which they were previously sensitive. AMR can be seen as an evolutionary response to interspecific interactions for survival in adverse conditions, and human activity (pollution with xenobiotics) only facilitates its spread. Therefore, antimicrobial compounds and AMR predate the emergence of humans [16, 17, 18, 19]. The recent pandemic, together with AMR, is forcing us to take the next step. The human gut is a reservoir of antibiotic-resistant microorganisms, but in fact, organisms in water, soil, plants, and animal life in the metabiota are the main reservoir. Collectively, the genes for AMR are called the resistome. A major cause of the spread of AMR is our agricultural and industrial activities [20, 21, 22, 23].
Properly selected and administered antibiotic therapy is an important component of our pharmacological interventions. There has been an increase in antibiotic use worldwide, with a 65% increase between 2000 and 2015. In treatment, we treat not only the host (our body), but also the microbiota. Broad-spectrum antibiotics do not differentiate between commensal and pathogenic bacteria and can lead to a 30% reduction in the microbiota and disruption of bacterial and phage communities. Phages interact with symbiotic bacteria, leading to the expression of genes responsible for the induction and transcription of lytic genes in prophages, lysis of infected bacterial cells, generation of new phage particles, and integration of the phage genome into the bacterial one [24, 25, 26]. This alters the phenotype and bacterial viability, conveys genes for antibiotic resistance and virulence, alters the ability of bacteria to produce toxins, increases their tolerance to oxidative and acid stress, and improves energy utilization. Temperate phages help the bacterial host adapt better due to the expression of newly acquired phenotypes [27, 28, 29]. DNA damage that destabilizes repressors causes prophage induction. The extent of the damage depends on the type of antibiotic. When used, they can damage bacteria important for our health: Clostridia, Bacteroides, Lactobacilli, and Bifidobacteria, and replace them with histamine-secreting strains of the same genera [30, 31, 32]. Changes in taxonomic composition have been linked to diseases such as inflammatory bowel disease (IBD) and asthma. Patients with asthma have been shown to have increased numbers of histamine-secreting bacteria in the gut [33, 34, 35]. Their use may have long-lasting effects on the gut microbiota during the first years of life. Microbial diversity is particularly strongly affected in anaerobic gut bacterial species.
The increased presence of species of the genus Proteobacteria containing PLP-dependent histidine decarboxylase is considered an indicator of intestinal dysbiosis and a cause of the manifestation of inflammatory diseases [36]. The change in the diversity of the healthy phageome in response to antibiotic therapy provokes the excitation of the DNA repair system regulated by the repressor LexA and the inducer RecA or the SOS response [37, 38]. The reduced amount of commensal bacteria during antibiotic treatment is compensated for by pathogens, leading to similar or more severe disease, which further worsens the clinical picture of patients.
Materials
Could AMR be both a threat and an opportunity for us? Extracting or synthesizing antibiotic molecules, stabilizing and testing them for side effects, and determining their ability to retain their properties under adverse conditions in the body requires a lot of time, effort, and money. Separately, resistance exists in nature even to drugs that have not yet been developed. This requires us to use our knowledge of the stages of colonization and interactions in the metabiota more actively. Using the proven benefits of probiotic strains and, at the same time, as “suppliers” of proteinaceous antibiotic-like substances of bacterial origin with a low probability of developing resistance will allow us to remodel the interactions between the components of the holobiont, pathogens, and microbial communities in our environment. They have been tested for effectiveness over billions of years of life on earth and will improve control of host-microbe-environment signaling, disease progression, and maintenance of homeostasis in this complex system. If we get this step right, we could climb to the next level of therapeutic approaches.
Probiotics have been part of our diet for centuries. Examples include Lactobacillus plantarum RL34 in cottage cheese, L. delbrückii subsp. Vulgaris 1043 and Streptococcus thermophilis L81 in Bulgarian yogurt (kiselo mljako), Lactococcus lactis B14 in boza (fermented food), Lactobacillus plantarum JLA-9 isolated from Suan-Tsai, a traditional Chinese fermented cabbage, etc. Fuller defines them as living organisms contained in food, which, after ingestion, can alter the intestinal microbiota and stimulate the immune system, and according to the International Scientific Association for Probiotics and Prebiotics, probiotics are defined as “live microorganisms which, when administered in adequate amounts, confer a health benefit.” Prebiotics are usually fibers or complex carbohydrates that serve as food for beneficial microorganisms. Probiotics properties include acid stability, specificity, lack of side effects, reduction of pathogenic microbial numbers and viability during storage. The benefits of probiotic strains for human and animal health are immunological - increased secretion of IgA locally and systemically, modeling of cytokine profiles and suppression of immune responses to food antigens and non- immunological - production of bacteriocins, stimulation of mucin secretion and strengthening of the intestinal barrier, lowering of intestinal pH, competition with pathogens for nutrients and adhesion sites and neutralization of superoxide radicals and toxins released by pathogenic bacteria. The bacteriocins secreted by them are ribosomally synthesized protein antibiotic-like substances that inhibit or inactivate other organisms, most often homologous or similar bacteria, with which they compete for nutrients and space. They are a heterogeneous group of protein substances differing in size, mechanism of action, microbial target, release, and amino acid sequence. Their inhibitory activity depends on specific receptors on target cells. To date, over 200 bacteriocins from the following groups have been studied: lantibiotic or nisin, mersacidin, cytolysin, and lactocin S groups. They are divided into those with narrow-spectrum antimicrobial bioactivity to strains closely related to the synthesizing one and those with broad-spectrum bioactivity to strains from different genera. The FDA has approved the use of bacteriocins as preservatives in the food industry. Their advantages over conventional antibiotics include tolerance to higher thermal stress, activity over a wider pH range, and non-toxic inhibition of target bacteria. Their protein nature minimizes the likelihood of resistance development among target bacteria due to their rapid-acting antimicrobial mechanisms that are effective at low concentrations - micromolar and nanomolar. Klaenhammer suggests that probably 99% of bacteria produce at least one bacteriocin. He created their classification in 1993, which was updated by Cotter, et al. in 2005 and last supplemented by Tagg H in 2006. Some of the better-known bacteriocins produced by pathogens are: E. coli – colicin, P. aeruginosa – pyocin, K. pneumonia – klebcin, Yersinia enterocolitica – corotovoricin, and St. aureus – lysostaphin.
For their proper use, knowledge of the stages of colonization of the host during ontogenesis, the mechanisms for entering into a permanent relationship, and the changes that occur with age is particularly important. In practice, food supplements containing probiotics are mono- and multi- strain assembled joint communities of 2 or more strains. Proper selection and cultivation achieve higher fitness and avoid competition. In addition to the “classic probiotics” (Lactobacillus and Bifidobacteria), new bacterial and fungal strains with potential for improving health are now being used. These are: Clostridium butyricum CBM588 or Miyairi strain, Saccharomyces boulardii, Saccharomyces cerevisiae, and Bacillus subtilis, Clausii, coagulans, and mesentericus.
Early colonization is crucial for shaping the immune system, neuroendocrine, and metabolic maturation, while colonization at later stages of life has a more limited effect. A question requiring clarification is whether there is a prenatal microbiome and whether it is possible for microbial fragments and metabolites to pass transplacentally and participate in shaping the fetal immune system. The intestines of infants after birth are free of microorganisms, but after only a few hours, we observe an increasing microbial diversity. The rapid colonization of the human body after birth is due to vertical and horizontal types of transmission.
In the first type of baby, through contact with the skin, milk, vaginal mucosa, and the maternal intestinal microbiota, the latter is dominant, and this transfer takes place. In the first 4 days, we observe a poor bacteriome and a rich phageome, while at 2 years, the bacteriome is already rich and diverse, and the phageome is poorer [39]. During the first month, the predominant aerobes (Enterobacteriaceae) begin to be replaced by strict anaerobes (Bifidobacterium, Bacteroides, and Clostridiales). Over time, the early colonizers, Bacteroides and Clostridiales, increase in number, while Bifidobacterium decreases, directly related to the cessation of breastfeeding and the transition to solid foods [40, 41, 42, 43, 44]. Early strains of Bifidobacterium are adapted to human milk oligosaccharides (HMOs) and some of them are stabilized and remain ready for vertical transmission to the next generation. HMOs oligosaccharides in infants have a bifidogenic prebiotic effect. They act as receptors-bait for Bifidobacterium sp. in the mucosal niche and also as antimicrobial adhesive substances preventing the adhesion of pathogens to the epithelium.
Heterogeneity is their characteristic quality. Each woman secretes structurally specific saccharides, which are reflected in vertical transfer. The strains that predominate after the transition to solid food use carbohydrates that are not part of the HMO. While the primary colonization can last for months, subsequent waves are shortened and in adults reach 2-4 weeks.
The first bacterial colonizers of the infant’s gut introduce colonizing phages or induced phages. For example, bifidobacteria transmitted through breastfeeding contain prophages known as bifidophages [45].
Early in life, the abundance of temperate phages is high because the biomass of bacterial species is low. As growth progresses, along with the expansion of bacterial species in the different niches of the gut, colonization by virulent phages from the crAss-like and Microviridae families accelerates, becoming established in early infancy. These families are considered to be specific to human hosts, and only a fraction of them can be shared between individuals-50% of the phageome is unique to each individual [46, 47, 48, 49]. In adults, the virome is abundant, persistent, and dynamically changing throughout our lives. The synchronous colonization of the human body by viruses and bacteria is an expression of their interactions.
The mode of delivery determines the colonizing strains. In vaginal delivery, Bifidobacterium, Escherichia, and Bacteroidetes predominate, which are more stable in the first year, while in cesarean section, the intestinal microbiota contains mainly hospital strains and is more unstable. Additional sources of microbes in horizontal transmission are considered to be hospital staff, father, siblings, relatives, contact with objects, and pets.
The composition of the human microbiota determines our predisposition to the development of noncommunicable, infectious, etc., diseases. Our task is to define how probiotic strains can help us in restoring the composition of the microbiota, inhibiting or inactivating other organisms through bacteriocins, limiting the spread of noncommunicable diseases (eczema, allergy, or asthma), alleviating their manifestations, strategies for protection against cancer and prevention, and treatment of infectious diseases. Each strain can play a role in solving one or more of these tasks. The genus Bifidobacterium, containing about 30 species, is one of the first colonizers and is involved in the early microbial assembly in infants and the associated with immune maturation, vaccine response, and metabolic programming. When present, four strains are mainly identified: Bifidobacterium breve, Bifidobacterium bifidum, Bifidobacterium longum subsp. longum and Bifidobacterium longum subsp. infantis. They are the most informative markers of GIT health. A two-year representative study tracked the impact of the absence of representatives of the genus in the United States. The main reasons for the decrease in the representation of the genus are considered to be delivery by cesarean section, antibiotic therapy, and feeding with adapted milk. Malesu MD investigated the reasons that led to the absence of the genus in a part of normally born and breastfed babies. Based on gene sequencing, he divided the babies into three groups. In the first group, including vaginally born and breastfed, an abundance of B. breve genes and those of HMO oligosaccharides was found.
In the second group, a higher presence of B. longum and Bacteroides with intermediate HMO potential was observed, and in the third (mainly after cesarean section), a lack of the genus Bifidobacterium and the presence of microbes such as Clostridium perfringens and other pathobionts, as well as increased levels of AMR genes and virulence factors (VF). The feces of the latter group contained lower amounts of thiamine and indole-3-lactate (ILA), which are responsible for immune maturation. The second and third groups were associated with a 3.2 and 3.0 times higher tendency to develop eczema, allergy, or asthma compared to children from the first group. The abundance of Bifidobacterium reduced the relative risk by 3.1 times and B. breve alone by 4.8 times. Gene sequencing looked for genes for the synthesis of phage repressors (Proteobacteria) and lipopolysaccharides (LPS) of Firmicutes - factors predicting adverse outcomes. The study is indicative of the importance of microbial representation in immune maturation [50].
Results
Possible use of Probiotic Strains in Clinical Practice
Restoring the Composition of the Microbiota:
- Probiotic strains, mainly strains Bifidobacterium breve, bifidum, and longum subsp. longum and longum subsp. Infants of the genus Bifidobacterium, as mono- and multistrains in sufficient quantities, can help us restore the composition of the microbiota and early microbial assembly in infants born by cesarean section, who have undergone previous antibiotic therapy and who are fed with adapted milk. By limiting changes in immune maturation and metabolic programming, we will be able to limit the spread of non-communicable diseases (eczema, allergy, or asthma).
- Lactobacillus reuteri is another strain that shows a similar trend. It is naturally found in breast milk. According to data from the 1960s, L. reuteri was part of the microbiome of 30-40% of the world’s population. Today, its presence is estimated to be only 10-20% the population. In sufficient quantities, it can help us restore the composition of the microbiota and microbial assembly in those who have undergone antibiotic therapy.
Inhibition or Inactivation of other Organisms through the Release of Bacteriocins: Bacteriocins produced by lactic acid bacteria (LAB) have received considerable attention due to their potential to be harmless to the human body and their ability to inhibit the growth of foodborne pathogens, such as Listeria monocytogenes, Escherichia coli, Staphylococcus aureus, and Bacillus spp. as well as in natural preservation and food safety. They are divided into four classes; class I, thermostable and lanthionine-containing; class II, thermostable and non-lanthionine-containing; class III, thermolabile, protein-like; and class IV, complex bacteriocins containing lipid or carbohydrate moieties. Their producers exist in fermented foods, and LAB are considered the dominant microorganism in fermented fish products.
Two have been identified in L. reuteri. Reutericyclin is produced in yeast and exhibits bacteriostatic and bactericidal effects against a broad spectrum of gram (+)
organisms and pathogens. Gram (-) bacteria are resistant to reutericyclin. Reutericyclin-producing strains persist for 10 years in industrial yeast fermentation, and it is secreted in concentrations active against competitors during the growth of L. reuteri. They are used for food preservation. The second compound, β-hydroxypropionaldehyde (3-HPA), or reuterin, is a broad-spectrum antimicrobial agent active against gram (+) and gram (-) bacteria, as well as against yeasts, molds, and protozoa. Reuterin demonstrates potent antimicrobial activity against a broad spectrum of Campylobacter spp. Isolates from human and poultry meat. It does not adversely affect the beneficial intestinal flora. The compound is resistant to proteolytic and lipolytic enzymes and remains active at different pH levels.
- Staphylococcus lugdunensis is a gram (+) bacterium considered a CoNS (coagulase-negative staphylococci) and was first described by Freney, et al. in 1988. It colonizes 30% to 50% of patients and, like other CoNS isolates, is a component of the skin microbiota of healthy individuals. S. lugdunensis produces lugdunin, an antibiotic with bactericidal activity against S. aureus and S. pneumoniae [51].
- Streptococcus salivarius K12 was first isolated from the throat of a New Zealand child and produces two lantibiotics, salivaricin A2 and B. They are ribosomally synthesized antimicrobial peptides. Salivaricin A (SalA), the first characterized lantibiotic of S. salivarius, is inhibitory to most strains of Streptococcus pyogenes. Salivaricin B is a 25-amino acid polycyclic peptide belonging to the type AII lantibiotic. Salivaricin B requires micromolar concentrations for its action. It interferes with cell wall biosynthesis. Transmission electron microscopy of salivaricin B-treated cells shows a reduction in cell wall thickness, in the absence of any apparent changes in cytoplasmic membrane integrity. Lantibiotics aid the survival of host bacteria in their preferred ecosystem by suppressing the growth of competing bacteria in the ecological niche [52, 53].
- Streptococcus lactis produces the polypeptide lantibiotic nisin. The first descriptions of the inhibitory properties of nisin appeared in 1944. Since the mid-20th century, it has been used as a food preservative with the number E234. Nisin is found in raw milk, is considered harmless to healthy people, and is degraded by digestive enzymes in a short time. Nisin is a rare exception to the “broad spectrum” and is effective against many gram (+) organisms, including Lactobacilli, Listeria monocytogenes, Staphylococcus aureus, Bacillus cereus, Clostridium botulinum, etc [54, 55].
- Lactobacillus plantarum JLA-9, isolated from Suan- Tsai, a traditional Chinese fermented cabbage, releases plantaricin JLA-9 and Lactobacillus plantarum LPL-1 from fermented fish products, releasing plantaricin LPL-1. It exhibits broad-spectrum antibacterial activity against Gram (+) and Gram (-) bacteria, especially Bacillus spp., high thermal stability (20 min, 121 °C), and narrow pH stability (pH 2.0-7.0). Its mode of action is interesting in inhibiting the growth of Bacillus cereus spores, responsible for both the self-limiting emetic and diarrheal forms as well as for rare and sometimes fatal cases of liver failure, pulmonary hemorrhage, and cerebral edema in children and young people. The onset of germination is considered a prerequisite for the action of plantaricin JLA-9. It inhibits growth by preventing the establishment of oxidative metabolism and disrupting the integrity of the membrane in germinating spores within 2 hours [56].
Limiting the Spread of Noncommunicable Diseases (Eczema, Allergies, or Asthma): Induction of the DNA repair system or SOS response because of antibiotic therapy is associated with the regulatory response of bacterial cells against the loss of diversity of the gut microbiota and phageome, pathogenic bacteria in the gut, and the induction of prophages [57, 58]. Intestinal autobionts produce indole-3-propionic acid (IPA) involved in the regulation of positive and negative metabolic homeostasis. IPA is produced mainly by Clostridium sporogenes, is a product of tryptophan, and is responsible for the maturation of lung cells, the prevention of allergic airway inflammation, and the development of asthma. In addition to limiting the spread of noncommunicable diseases (eczema, allergy, or asthma), in patients with low levels of IPA in the blood, we observe insulin resistance, overweight, a tendency to low-grade inflammation, and symptoms of metabolic syndrome, in contrast to those with high IPA. There is also a known negative relationship with polymorbidity. Patients with operational taxonomic units (OTUs) including Ruminococcus, Alistipes, Blautia, Butyrivibrio, and Akkermansia are in the high-IPA group, while in the low-IPA group, we observe an abundance of Escherichia-Shigella, Megasphera, and the genus Desulfovibrio [59]. Fungal dysbiosis may further exacerbate the manifestation of allergic diseases. Lactobacillus reuteri is thought to induce the SOS response by activating specific metabolic pathways in the GIT [60]. Our ability to use it as a probiotic monoproduct during the first 3 years of life after antibiotic therapy will likely allow us to reduce the potential risk of developing health problems in at least some patients.
Using the metabolic pathways of Lactobacillus microbes to break down dietary purines to produce energy and building blocks helps break down urates, reducing their reabsorption and hyperuricemia. As a component of the renal microbiota, the uroprotective bacterium L. crispatus, through the inclusion of CaOx inhibitors and promoters together with a strain of E. coli, drives CaOx crystallization and kidney stone formation. The use of antibiotics alters the renal microbiota and shifts the balance from beneficial Lactobacillus to E. coli, leading to stone formation. It is suggested that Lactobacillus is a protective factor and Enterobacteriaceae is a pathogenic factor contributing to the formation of urate kidney stones.
Attenuation of the Manifestations of Noncommunicable Diseases: Allergic diseases are caused by an inappropriate activation of the Th2 immune response. Th2 cytokines (IL-4 and IL-5) induce B cells to produce antibodies - immunoglobulin E (IgE) and activate eosinophils (Eo), basophils (Ba), and MCs. The latter infiltrate the airways and, upon degranulation, release inflammatory molecules and proteases, leading to tissue edema in asthmatic lungs. The Th2 response is considered anti-inflammatory, but in allergic disease, overproduction of Th2 cells leads to harmful immune responses. The use of probiotic monoproducts in allergies has certain benefits:
- L. acidophilus significantly improves nasal symptoms in some patients with perennial allergic rhinitis.
- L. paracasei affects the quality of life in allergic rhinoconjunctivitis in children.
- L. rhamnosus GG improves the immune response in immunotherapy [61, 62, 63, 64].
- Clostridium butyricum may alleviate allergic reactions, possibly by modulating Tregs. The strain CGMCC0313.1 in mouse models of asthma reduces airway hyperreactivity and lung inflammation by suppressing the Th2 response and increasing IL-10 production. In this model, the number of infiltrating MCs, levels of Th2 cytokines (IL-4 and IL-5), markers of MC degranulation, and Ig E levels are reduced [65, 66, 67].
We have yet to specify at what point and in which patients it is appropriate to apply them alone or in combination with other therapeutic strategies.
V. Strategies for Protection Against Cancer:
- Metabolites and cell wall components of Bifidobacterium spp. - Bifidobacterium bifidum, B. longum subsp. longum and B. pseudolongum stimulate the adaptive immune response against tumor cells and can be used as a microbial defense strategy and in cancer immunotherapy [68, 69].
- In mouse models, the use of L. reuteri strains with high levels of cell wall peptidoglycan in the treatment of melanoma leads to an increase in antitumor CD8+ T cell responses by metabolizing the tryptophan indole-3- aldehyde ligand of the aryl carbon receptor (ANR) and facilitates therapy with immune checkpoint inhibitors [70, 71].
- Bacteriocins produced by lactic acid bacteria have anticancer properties, inhibiting the growth of various cancer cells in vitro and in vivo. The mechanisms of their antitumor action include induction of apoptosis, disruption of the cell cycle, suppression of cell migration, and angiogenesis [72].
- Immune checkpoints (ICIs) are proteins that prevent excessive immune responses. If checkpoint proteins on cancer cells bind to T-cell checkpoint receptors, the T cell is turned off, and cancer rejection is inhibited. Bacteria often “seed” cancers. The microbiota mediates the immune system’s response to cancer. Metabolites and cell wall components are involved in priming the adaptive immune response by secreting cytokines and activating immune effector cells (tumor-infiltrating lymphocytes-TILs) [68, 69, 73]. Taking antibiotics shortly before or after starting immune checkpoint inhibitor (ICI) therapy negatively affects overall survival (OS) and progression-free survival (PFS).
Our eating habits can have a similar effect. Overacre- Delgoffe A found in mouse models that Sucralose consumption can alter the gut microbiota and interfere with cancer immunotherapy, weakening the response of cancer patients to immunotherapy. Sucralose consumption negatively affected the response to immune checkpoint inhibitors and their progression-free survival. Mice consuming Sucralose showed resistance to immune checkpoint blockade, increased tumor growth, and decreased overall survival. It is believed that the intake of prebiotics is more appropriate because during digestion, prebiotics are broken down to short-chain fatty acids by bacterial enzymes, possibly serving as receptor bait for certain strains and as antimicrobial adhesive substances preventing the adhesion of pathogens to the epithelium and stimulating the growth of certain microbiota. Prevention and Treatment of Infectious Diseases: They require a flexible use of pharmacological and non- pharmacological interventions in conjunction with probiotics, dietary components, and vaccine strains. This requires:
- Sequencing patients early in the disease to identify elevated viral load.
- Bacterial carriage by whole genome sequencing (WGS) is more sensitive in detecting pathogens after antimicrobial treatment, and fluorescence in situ hybridization (FISH) helps detect bacterial pathogens in biofilms [74, 75, 76, 77].
- Amplicon-based sequencing techniques targeting the 16S ribosomal RNA gene (16S rRNA) identify bacterial pathogens in critical infections when standard test samples are negative due to previous antibiotic treatment and shorten the time to 2 days. These techniques will guide us in using the right combination.
- Monocolonization with Saccharomyces cerevisiae maintains intestinal homeostasis and protects against virus-induced lung inflammation and intestinal barrier disruption [78, 79].
- Oral administration of Bifidobacterium longum in mice induces reactive oxygen species in alveolar macrophages and enhances protection against pulmonary infection caused by Klebsiella pneumoniae [80].
- Lactobacillus strains attenuate viral diarrhea caused by rotavirus infection and, together with commensals from the genera Lactobacillus and Bacteroides, inhibit rotavirus attachment by modulating glycan receptors on intestinal epithelial cells [81].
- Bacillus clausii protects the intestine from rotavirus infection by inducing the synthesis of β-defensin 2 and cathelicidin, reducing the proportion of necrotic enterocytes, and increasing the synthesis of mucin and TJ proteins [82].
- Streptococcus lactis inhibits Clostridium botulinum via the lantibiotic nisin [54, 55].
- Saccharomyces boulardii CNCM 1-745 secretes a protease that degrades the toxin A169 of C. difficile.
- Saccharomyces boulardii alters gut microbiota, reduces hepatic fat accumulation, inflammation, and total fat mass in mouse models of obesity and T2D. In healthy humans, it does not affect the microbiota, but it helps restore eubiosis of the gut microbiota after diarrhea [83].
- Clostridium butyricum CBM588 of the genus Furmicutes processes undigested fiber and secretes short-chain fatty acids (SCFA), acetate (C2), and butyrate (C4). When secreted into the colon, these are involved in modulating immune homeostasis, improving intestinal barrier function, and influencing inflammation [84]. Genomic analyses of C. butyricum have shown that some strains lack genes responsible for pathogenicity and coding for toxin production [85]. The observed effects are:
- Butyrate (C4) stimulates mucin production by goblet cells in-vitro and increases mucus layer thickness [86].
- C. butyricum modulates immune homeostasis by stimulating IL-17 production by γδ T cells, a subset of intraepithelial T cells that act as part of the first line of defense in the lamina propria of the colon [87, 88].
- In mouse models, C. butyricum promotes IL-10 secretion by Treg cells through the production of anti- inflammatory lipid metabolites - palmitoleic acid, prostaglandin metabolites, etc [89]. It also suppresses DC maturation and the appearance of an inflammatory phenotype as well as the levels of pro-inflammatory cytokines of Th1 and Th17 cells - IFNγ and IL-17 [90].
- In antibiotic-associated diarrhea (AAD), Seki reported a reduction in the incidence of diarrhea from 59% in the group receiving antibiotics alone to 5% in that receiving C. butyricum.
- It is suggested to participate in modeling the composition of the gut microbiome, increasing bacterial taxa from the genera Lactobacillus and Bifidobacteria.
- Induced phages prevent infection of commensal bacteria by other lysogenic or lytic phages through a phenomenon of superimmunity.
- C. butyricum and commensal flora inhibit the growth of C. difficile by consuming nitrogen-containing amino acids, sialic and succinic acids, and glycans, which are food sources for C. difficile [91, 92].
- Bacillus subtilis helps increase bacterial taxa from the genera Lactobacillus and Bifidobacteria in the ileum and cecum, and reduces coliforms and Clostridium perfringens in the cecum.
- Bacillus coagulans and B. mesentericus stimulate intestinal peristalsis, metabolic environment, limit toxin accumulation, and compete for epithelial cell surface binding sites with vancomycin-resistant enterococci and E. coli [93].
- L. rhamnosus GG, Bifidobacterium spp., L. acidophilus, and S. thermophilus reduce nasal colonization with potential pathogens such as S. aureus, S. pneumoniae, and β-hemolytic streptococci and reduce episodes of respiratory infections [94, 95, 96].
- Lower incidence of respiratory tract infections in neonates receiving prebiotics and probiotics [97, 98, 99].
- The use of S. salivarius and S. oralis limits the abundance of S. aureus and increases the total number of beneficial microbial taxa [100].
- The use of topical vaccines or oral supplements containing α-hemolytic streptococci (AHS) - S sanguis, S. mitis, S. salivarius, and S. oralis - helps to increase microbial diversity and prevent diseases caused by respiratory pathogens by inhibiting them with bacteriocins.
- Streptococcus mitis influences the adaptive immune response to induce cross-reactive immunity (antibodies and IL-17) to S. pneumoniae in mice. A similar phenomenon is observed with Neisseria lactamica and N. meningitidis.
- L. casei induces the production of antibodies against S. pneumoniae through the expression of the surface protein antigen Pspa [101].
- Bifidobacterium infantis modifies the IL-10/IL-12 ratio and has anti-inflammatory effects. It also exhibits immunomodulatory effects by increasing the number of mucosal dendritic cells and decreasing Th1 and Th17 helper T cells.
- Streptococcus salivarius inhibits mucosal binding of S. pneumoniae, Haemophilus haemolyticus inhibits nontypeable Haemophilus influenzae, and the genus Corynebacterium competes for mucosal binding with pathogens of the GI tract [102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112].
- The surface protein layer of the genus Lactobacillus provides antimicrobial inhibitory effects by competing for binding sites on the surface of epithelial cells and antagonizing viral entry and replication, but not attachment. Limits colonization of S. aureus [113, 114]. The ability to attach to epithelial surfaces is important for maintaining persistent colonization in the intestine and other mammalian tissues, but even an optimal microbiota dominated by the genus Lactobacillus does not guarantee a reduction in the risk of some infections.
Diet is determined by our habits, customs, and individual preferences and has a significant impact on the composition of the microbiota. Similar to how our body follows its internal clock (circadian rhythm), intestinal microorganisms also have their own rhythms corresponding to time and food intake. These rhythms cause daily fluctuations in the composition and function of intestinal microbes. Tracking them shows the significant impact of food intake during daily activity and sleep time on our metabolic health.
Cooking is a form of food processing that increases digestibility and reduces the amounts of substances reaching the colonic microbiota. It most severely affects individuals when they switch from a diet rich in fruits and vegetables to one high in meat and energy-dense foods. A study shows that only 5% of the population in our industrialized society consumes an adequate amount of fiber. The type of diet plays a role in the health-disease balance. Microbes provide metabolic capacity for energy extraction and are associated with obesity. Dietary fiber is a part of food that can only be broken down by microbes into simpler products - short- chain fatty acids (SCFA: C2-acetate, C3-propionate, and C4- butyrate). Fiber content determines the composition of the intestinal microbiota, and the Prevotella/Bacteroides ratio is used as an indicator [115, 116, 117, 118, 119, 120, 121]. They affect intestinal and extraintestinal physiology, including neuroendocrine, brain, immune, and metabolic functions. Our diet determines which microorganisms will survive and which will disappear, and loss of dietary diversity leads to loss of microbial diversity. Loss of microbiota diversity limits self-regulation (loss of autobionts and abundance of pathobionts), reduces defensive competition against pathogens, and limits signaling to the host that maintains homeostasis in this complex system. This also leads to the risk of developing many chronic inflammatory and metabolic diseases. Long- term dietary patterns determine the enterotype of the gut microbiota [122]. The Bacteroides enterotype is associated with reduced microbial activity and genetic diversity, insulin resistance, risk of obesity and nonalcoholic steatosis, and the Prevotella enterotype with a diet dominated by plant carbohydrates [123]. Diet during infectious diseases can have unintended effects and worsen the course of the disease. It is important to limit the consumption of histamine-rich foods during illness - nuts, tomato paste, mature cheese, cocoa, chocolate, sauerkraut, smoked fish, salami, ham, dried fruits, some fruits and vegetables, yeast, and others, as they can increase neurological, gastrointestinal, and respiratory complaints. Almost all organs and systems have four types of histamine receptors - H1, H2, H3, and H4. Calcium, magnesium, and casein in dairy products, as well as iron in dietary supplements, can affect the action of quinolones and tetracyclines.
Discussion
The recent pandemic and the increasing AMR force us to take the next step due to the pressure on the pathogen-host- environment system to climb the next tread in therapeutic approaches. This should not be done blindly, but after weighing both the benefits and negatives associated with the application of these microorganisms and their products. The Food and Agriculture Organization (FAO) and the World Health Organization have clearly defined guidelines for the safety and efficacy of probiotic strains, and these characteristics should be used when preparing preparations. They are:
- Genus, species, strain.
- Minimum amount of viable microorganisms at the expiration date of the preparation.
- Effective dose.
- Safety, synergistic interaction with the microbiota, stability in gastrointestinal conditions, protective effect, validation, and quality of the strain.
They will guarantee the effectiveness of the strains used and the exclusion of potential contaminants in their composition through the use of standard molecular techniques - PCR and direct sequencing to identify mutations conferring specific individual traits of resistance and virulence. This will prevent the marketing of preparations such as those containing four bacterial strains of B. clausii - O/C, SIN, N/R, and T, characterized by an expanded resistance pattern to many antibiotics. Phenotypically, they have high levels of resistance to chloramphenicol, streptomycin, rifampicin, and tetracycline. These probiotic preparations have been used in clinical practice during antibiotic therapy to prevent intestinal microbial imbalance. The supposed benefit in this case conflicts with the safety criteria that require reducing the transmission of antibiotic resistance from probiotics to pathogens in the gastrointestinal tract. Monitoring the pattern and stability of resistance spectra of probiotics is important in assessing the risk-benefit ratio. The degree of danger is different for bacteria with natural phenotypic traits and for those with acquired resistance (via mobile genetic elements - plasmids and transposons). Resistance through mutation has a lower risk of spreading. AMR should be considered as an evolutionary response to interspecies interactions for survival under adverse conditions, and therefore, resistance exists even to drugs that have not yet been developed. When evaluating the benefits of probiotic preparations for clinical practice, the prevention of microbial imbalance should probably be given priority [124, 125, 126].
To achieve the desired results and avoid future problems, it is good to find the answers to some questions in advance. We need to establish the recommended doses for the different niches. As sufficient for colon colonization, FAO recommends 108-1010 CFU to achieve clinical benefit. Probiotics used for the prevention and treatment of infectious diseases and those for restoring the composition of the microbiota should meet different criteria. In the former, the appropriate probiotic is a transient strain or strains, whose task is to “load” the resources of the ecological niche and create the most unfavorable conditions for the pathogen.
In the latter, it is more appropriate to be monoproducts, to avoid competition and create the opportunity to become permanent members of the microbiota. The composition alone is not enough. It is also necessary to use dietary solutions to stabilize them in the niche. Another question that needs to be answered is whether their use is appropriate in immunocompromised and immunosuppressed patients. It is always better to anticipate possible risks than to seek a solution to problems we have caused.
This year’s congress of the French Society of Pediatrics discussed guidelines for the use of probiotics. Mosca A, MD, also presented the upcoming recommendations of the European Society for Pediatric Gastroenterology, Hepatology and Nutrition (ESPGHAN) on probiotics in adapted milk for infants.
According to ESPGHAN, a limited number of strains with proven benefits have been identified. They are Lactobacillus rhamnosus GG (ATCC 53103), Saccharomyces boulardii (CNCM I-745), Lactobacillus reuteri (DSM 17938), and the combination of Lactobacillus rhamnosus (19070- 2) and Lactobacillus reuteri (DSM 12246). In France, the term “probiotic” has been authorized by the Directorate General for Competition Policy, Consumer Affairs, and Fraud Control since December 2022. According to ESPGHAN experts, many products do not provide information on the exact composition, dosage, and potential contaminants. It is recommended to use strains supported by at least two randomized controlled trials per condition. In human milk, oligosaccharides such as 2´-fucosyllactose, 3-fucosyllactose, lacto-N-tetraose, 3´-sialyllactose, and 6´-sialyllactose, in addition to the effect of decoy receptors for bacteria and adhesive substances preventing the adhesion of pathogens to the epithelium, at high doses, soften the stool.
Microbiota science, metabolomics, precision medicine, and the One Path, One Health concept should not be developed as separate directions, but rather as components of a puzzle. It is time to move from a one-dimensional to a three- dimensional model in understanding the depth, diversity, and complexity of the ongoing interactions and relationships in nature in which we also participate. The therapeutic use of probiotics can be seen as a beginning in mastering the two- dimensional model of these complex processes.
In poultry farming, they have already proven themselves as an alternative to antibiotics [127]. Simplifying complex phenomena and relationships to obtain understandable concepts sometimes leads to the opposite effect - loss of the overall picture.
Metabolic dependencies are the main drivers of the existence of species in nature. Over the years, various life forms have evolved and continue to evolve. They are characterized by rationality and profitability when using the window of opportunity. It is time for us to do the same. It is very likely that the 101000 metabolic pathways identified by Wagner A in nature represent different scenarios played out for coping with adverse conditions and survival of organisms. Each microorganism, to survive in a given ecosystem, constantly passes through the commensal- pathogen continuum. During the first three years of our lives, the formation of the phenotype is in its infancy, as permanent relationships in the holobiont have not yet been established. During this period, our interventions must be extremely balanced so as not to push the formation of the phenotype in one direction or another, the consequences of which are currently unpredictable. Through probiotics and changing our diet, we may have the opportunity to restore homeostasis in the holobiont.
Conclusion
Pathobionts or exogenous pathogens require increased energy resources during their expansion, unlike commensals. The use of transient probiotic strains increases competition for adhesion and nutrients with those with pathogenic potential. Applied in sufficient quantities, they can create a maximally hostile environment for colonization of pathogens, limiting invasion, overgrowth of the latter, and even returning to the orbit of commensalism (Corynebacterium spр.→Staphylococcus aureus). Unlike broad-spectrum antibiotics, which do not distinguish between commensals and pathogenic bacteria and can reduce the microbiota by up to 30%, probiotic strains inhibit or inactivate other organisms without adversely affecting the beneficial flora.
To achieve our goals, however, probiotics must stop being food supplements and begin to strictly meet safety and efficacy criteria.
References
-
Giri S, Waschina S, Kaleta C, Kost C (2019) Defining division of labor in microbial communities. J Mol Biol 431: 4712-4731.
-
West SA, Diggle SP, Buckling A, Gardner A, Griffin AS (2007) The social lives of microbes. Annu Rev Ecol Evol Syst 38: 53-77.
-
Margulis L (1991) Symbiosis as a source of evolutionary innovation: speciation and morphogenesis. In: Cambridge MA MLFR (Edn.), Symbiogenesis and Symbionticism, MIT Press, pp: 1-14.
-
Rosenberg E, Koren O, Reshef L, Efrony R, Rosenberg IZ (2007) The role of microorganisms in coral health, disease and evolution. Nature Reviews Microbiology 5: 355.
-
Simon JC, Marchesi JR, Mougel C, Selosse MA (2019) Host-microbiota interactions: from holobiont theory to analysis. Microbiome 7: 5.
-
Bordenstein SR, Theis KR (2015) Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. Plos Biology 13(8): e1002226.
-
Faure D, Simon JC, Heulin T (2018) Holobiont: a conceptual framework to explore the eco-evolutionary and functional implications of host-microbiota interactions in all ecosystems. New Phytologist 2018: 1321-1324.
-
Zilber-Rosenberg I, Rosenberg E (2008) Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiol Rev 32(5): 723-735.
-
Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH, et al. (2012) Structure, function and diversity of the healthy human microbiome. Nature 486(7402): 207- 214.
-
Lloyd-Price J, Abu-Ali G, Huttenhower C (2016) The healthy human microbiome. Genome Med 8(1): 51.
-
Morris JJ, Lenski RE, Zinser ER (2012) The Black Queen Hypothesis: evolution of dependencies through adaptive gene loss. mBio 3(2): e00036.
-
Sachs JL, Hollowell AC (2012) The origins of cooperative bacterial communities. mBio 3: e00099-12.
-
Zelezniak A, Andrejev S, Ponomarova O, Mende DR, Bork P, et al. (2015) Metabolic dependencies drive species co- occurrence in diverse microbial communities. Proc Natl Acad Sci, USA, 112: 6449-6454.
-
Canon F, Nidelet T, Guédon E, Thierry A, Gagnaire V (2020) Understanding the Mechanisms of Positive Microbial Interactions That Benefit Lactic Acid Bacteria Co-cultures. Front Microbiol 4(11): 2088.
-
Logan LK, Weinstein RA (2017) The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of global menace. J Infect Dis 215(suppl_1): S28-S36.
-
Walker AW, Hoyles L (2003) Human microbiome myths and misconceptions. Nat Microbiol 8: 1392-1396.
-
Shanahan F, Hill C (2019) Language, numeracy and logic in microbiome science. NatRev Gastroenterol Hepatol 16: 387-388.
-
Miller BM, Liou MJ, Lee JY, Bumler AJ (2021) The longitudinal and cross-sectional heterogeneity of the intestinal microbiota. Curr Opin Microbiol 63: 221-230.
-
Lee JY, Tsolis RM, Bumler AJ (2002) The microbiome and gut homeostasis. Science 377(6601): eabp9960.
-
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez BMG, et al. (2012) Human gut microbiome viewed across age and geography. Nature 486: 222-227.
-
Ansaldo E, Farley TK, Belkaid Y (2021) Control of immunity by the microbiota. Annu Rev Immunol 39: 449-479.
-
Carmody RN, Sarkar A, Reese AT (2021) Gut microbiota through an evolutionary lens. Science 372: 462-463.
-
Suzuki TA, Fitzstevens JL, Schmidt VT, Enav H, Huus KE, et al. (2002) Co-diversification of gut microbiota with humans. Science 377: 1328-1332.
-
Ungaro F, Massimino L, D’Alessio S, Danese S (2019) The gut virome in inflammatory bowel disease pathogenesis: from metagenomics to novel therapeutic approaches. United European Gastroenterology Journal 7(8): 999- 1007.
-
Wang X, Kim Y, Ma Q, Hong SH, Pokusaeva K, et al. (2010) Cryptic prophages help bacteria cope with adverse environments. Nature Communications 1(1): 1-9.
-
Modi SR, Lee HH, Spina CS, Collins JJ (2013) Antibiotic treatment expands the resistance reservoir and ecological network of the phage metagenome. Nature 499(7457): 219-222.
-
Simmons SS, Isokpehi RD, Brown SD, McAllister DL, Hall CC, et al. (2011) Functional annotation analytics of Rhodopseudomonas palustris genomes. Bioinform Biol Insights 5: 115-129.
-
Oda Y, Larimer FW, Chain PSG, Malfatti S, Shin MV, et al. (2008) Multiple genome sequences reveal adaptations of a phototrophic bacterium to sediment microenvironments. Proc Natl Acad Sci USA 105(47): 18543-18548.
-
Ceapa C, Davids M, Ritari J, Lambert J, Wels M, et al. (2016) The variable regions of Lactobacillus rhamnosus genomes reveal the dynamic evolution of metabolic and host-adaptation repertoires. Genome Biol Evol 8(6): 1889-1905.
-
Kohanski MA, Dwyer DJ, Collins JJ, White MV, O’Mahony L, et al. (2010) How antibiotics kill bacteria: from targets to networks. Nat Rev Microbiol 8: 423-435.
-
Falony G, Joossens M, Silva SV, Wang J, Darzi Y, et al. (2003) Population-level analysis of gut microbiome variation. Science 352(6285): 560–564.
-
Guarner F, Malagelada JR (2003) Gut flora in health and disease. Lancet 361(9356): 512-519.
-
Kostic AD, Xavier RJ, Gevers D (2014) The microbiome in inflammatory bowel disease: current status and the future ahead. Gastroenterology 146(6):1489–1499.
-
Huang YJ, Boushey HA (2015) The microbiome in asthma. J Allergy Clin Immunol 135(1): 25-30.
-
Barcik W, Pugin B, Westermann P, Perez NR, Ferstl R, et al. (2016) Histamine-secreting microbes are increased in the gut of adult asthma patients. J Allergy Clin Immunol 138(5): 1491-1494.
-
Mukhopadhya I, Hansen R, El-Omar EM, Hold GL (2012) IBD-what role do Proteobacteria play? Nat Rev Gastroenterol Hepatol 9(4): 219-230.
-
Gutiérrez B., Domingo-Calap P (2020) Phage therapy in gastrointestinal diseases. Microorganisms 8: 1420.
-
McFarland L V (2008) Antibiotic-associated diarrhea: epidemiology, trends, and treatment. Future Microbiol 3: 563-578.
-
Cao Z, Sugimura N, Burgermeister E, Ebert MP, Zuo T, et al. (2022) The gut virome: a new microbiome component in health and disease. Ebio Medicine 81: 104113.
-
Kennedy KM, de Goffau MC, Perez Muñoz ME, Marie- Claire A, Fredrik B, et al. (2023) Questioning the fetal microbiome illustrates pitfalls of low-biomass microbial studies. Nature 613: 639-649.
-
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Bello MGD, et al. (2012) Human gut microbiome viewed across age and geography. Nature 486: 222-227.
-
Roswall J., Olsson LM, Datchary KP, Kristiansen K, Dahlgren J, et al. (2021) Developmental trajectory of the healthy human gut microbiota during the first 5 years of life. Cell Host Microbe 29(5): 765-776.e3.
-
Enav H, Bäckhed F, Ley RE (2022) The developing infant gut microbiome: A strain-level view. Cell Host Microbe 30(5): 627-638.
-
Masi AC, Stewart CJ (2021) Untangling human milk oligosaccharides and infant gut microbiome. iScience 25(1): 103542.
-
Tiamani K, Luo S, Schulz S, Xue J, Costa R, et al. (2022) The role of virome in the gastrointestinal tract and beyond. FEMS Microbiol 46: 1-12.
-
Dahlman S, Avellaneda-Franco L, Barr J (2021) Phages to shape the gut microbiota? Curr Opin Biotechnol 68: 89-95.
-
Shkoporov AN, Clooney AG, Sutton TDS, Ryan FJ, Daly KM, et al. (2019) The human gut virome is highly diverse, stable, and individual-specific. Cell Host Microbe 26: 527.
-
Liang G, Bushman FD (2021) The human virome: assembly, composition and host interactions. Nat Rev Microbiol 19: 514–527.
-
Cao Z, Sugimura N, Burgermeister E, Ebert MP, Zuo T, et al. (2022) The gut virome: a new microbiome component in health and disease. E Bio Medicine 81: 104113.
-
Jarman JB, Torres PT, Stromberg S, Sato H, Stack C, et al. (2025) Bifidobacterium deficit in United States infants drives prevalent gut dysbiosis. Commun Biol 8(1): 867.
-
Heilbronner S., Foster T (2020) Staphylococcus lugdunensis: a Skin Commensal with Invasive Pathogenic Potential, Clin Microb 34: 2.
-
Dufour А, Hindré Т, Haras D, Pennec JP (2007) The biology of lantibiotics from the lacticin 481 group is coming of age. FEMS Microbiol Rev 31(2): 134-67.
-
Barbour А, Tagg J, Abou-Zied ОК, Philip K (2016) New insights into the mode of action of the lantibiotic salivaricin B. Scientific Reports 6: 31749.
-
Kirtonia K, Salauddin M, Bharadwaj KK, Pati S, Dey A, et al. (2021) Bacteriocin: A new strategic antibiofilm agent in food industries, Biocatalysis and Agricultural Biotechnology 102141: 1878-8181.
-
Chen H, Hoover DG (2006) Bacteriocins and their Food Applications.
-
Wang Y, Qin Y, Xie Q, Zhang Y, Hu J, et al. (2018) Purification and Characterization of Plantaricin LPL-1, a Novel Class IIa Bacteriocin Produced by Lactobacillus plantarum LPL-1 Isolated From Fermented Fish. Front Microbiol 9: 2276.
-
Early antibiotic exposure can trigger long-term susceptibility to asthma. Monash University.
-
Sanders DJ, Inniss S, Sebepos-Rogers G, Rahman FZ, Smith AM (2021) The role of the microbiome in gastrointestinal inflammation. Biosci 41: 3850.
-
Ballanti M, Antonetti BL, Mavilio BM, Casagrande BV, Moscatelli A, et al. (2024) Decreased circulating IPA levels identify subjects with metabolic comorbidities: A multi-omics study, Pharmacol Res 204: 107207.
-
Kirsch J M, Brzozowski RS, Faith D, Round JL, Secor PR, et al. (2021) Bacteriophage-Bacteria interactions in the gut: from invertebrates to mammals. Ann. Rev 8: 95-113.
-
Ishida Y, Nakamura F, Kanzato H, Sawada D, Hirata H, et al. (2005) Clinical effects of Lactobacillus acidophilus strain L-92 on perennial allergic rhinitis: a double-blind, placebo-controlled study. J Dairy Sci 88: 527-533.
-
Lin WY, Fu LS, Lin HK, Shen CY, Chen YJ (2014) Evaluation of the effect of Lactobacillus paracasei (HF.A00232) in children (6-13 years old) with perennial allergic rhinitis: a 12-week, double-blind, randomized, placebo- controlled study. Pediatr Neonatol 55: 181-188.
-
Jerzynska J, Stelmach W, Balcerak J, Woicka-Kolejwa K, Rychlik B, et al. (2016) Effect of Lactobacillus rhamnosus GG and vitamin D supplementation on the immunologic effectiveness of grass-specific sublingual immunotherapy in children with allergy. Allergy Asthma Proc 37: 324-334.
-
Kang HM, Kang JH (2021) Effects of nasopharyngeal microbiota in respiratory infections and allergies. Clin Exp Pediatr 64(11): 543-551.
-
Berger A (2000) Th1 and Th2 responses: what are they? BMJ 321: 424.
-
Juan Z, Zhao-Ling S, Ming-Hua Z, Chun W, Hai-Xia W, et al. (2017) Oral administration of Clostridium butyricumCGMCC0313-1 reduces ovalbumen-induced allergic airway inflammation in mice. Respirology 22: 898-904.
-
Krystel-Whittemore M, Dileepan KN, Wood JG (2015) Mast Cell: a Multi-Functional Master Cell. Front Immunol 6: 620.
-
Lee KA, Thomas AM, Bolte LA, Bjork JR, de Ruijter LK, et al. (2022) Cross-cohort gut microbiome associations with immune checkpoint inhibitor response in advanced melanoma. Nat Med 28: 535-544.
-
Choi Y, Lichterman JN, Coughlin LA, Poulides N, Li W, et al. (2023) Immune checkpoint blockade induces gut microbiota translocation that augments extraintestinal antitumor immunity. Sci Immunol 8: 8.
-
Bender MJ, McPherson AC, Phelps CM, Davar D, Zarour HM, et al. (2023) Dietary tryptophan metabolite released by intratumoral Lactobacillus reuteri facilitates immune checkpoint inhibitor treatment. Cell 186: 1846-1862.
-
Simpson RC, Shanahan ER, Batten M, Reijers ILM, Read M, et al. (2022) Diet-driven microbial ecology underpins associations between cancer immunotherapy outcomes and the gut microbiome. Nat Med 28: 2344-2352.
-
Niamah AK, Al-Sahlany STG, Verma D, Shukla RK, Patel AR, et al. (2024) Emerging lactic acid bacteria bacteriocins as anti-cancer and anti-tumor agents for human health. Heliyon 10(7): e37054.
-
Tanoue T, Morita S, Plichta DR, Skelly AN, Suda W, et al. (2019) A defined commensal consortium elicits CD8 T cells and anticancer immunity. Nature 565: 600-605.
-
Stepinska M, Olszewska-Sosinska O, Lau-Dworak M, Zielnik-Jurkiewicz B, Trafny E (2013) Identification of intracellular bacteria in adenoid and tonsil tissue specimens: The efficiency of culture versus fluorescent in situ hybridization FISH. Curr Microbiol 68: 21-29.
-
Klein EY, Boeckel TPV, Martinez EM, Pant S, Gandra S, et al. (2018) Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Environmental Sciences 115(15): E3463-E3470.
-
Neuman H, Forsythe P, Uzan A, Avni O, Koren O (2018) Antibiotics in early life: dysbiosis and the damage done. FEMS Microbiol Rev 42: 489-499.
-
Francino MP (2015) Antibiotics and the human gut microbiome: dysbiosis and accumulation of resistance. Front Microbiol 6: 1543.
-
Mason KL, Downward JRE, Mason KD Falkowski NR, Eaton KA, et al. (2012) Candida albicans and bacterial microbiota interactions in the cecum during recolonization following broad-spectrum antibiotic therapy. Infection and Immunity 80(10): 3371-3380.
-
Hogan DA, Vik A, Kolter RA (2004) Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol Microbiol 54: 1212- 1223.
-
Vieira AT, Rocha VM, Tavares L, Garcia CC, Teixeira MM, et al. (2016) Control of Klebsiella pneumoniae pulmonary infection and immunomodulation by oral treatment with commensal probiotic Bifidobacterium longum 51A. Microbes Infect 18: 180-189.
-
Varyukhina S, Freitas M, Bardin S, Robillard E, Tavan E, et al. (2012) Glycan-modifying bacteria-derived soluble factors from Bacteroides thetaiotaomicron and Lactobacillus casei inhibit rotavirus infection in human intestinal cells. Microbes and Infection 14(3): 273-278.
-
Paparo L, Tripodi L, Bruno C, Pisapia L, Damiano C, et al. (2020) Protective action of Bacillus clausii probiotic strains in an in vitro model of Rotavirus infection. Sci Rep 10(1): 12636.
-
More MI, Swidsinski A (2015) Saccharomyces boulardii CNCM I-745 supports regeneration of the intestinal microbiota after diarrheic dysbiosis - a review. Clin Exp Gastroenterol 8: 237-255.
-
Koh A, De Vadder F, Kovatcheva-Datchary P, Backhed F (2016) From Dietary Fiber to Host Physiology: short- Chain Fatty Acids as Key Bacterial Metabolites. Cell 165(6): 1332-1345.
-
Cassir N, Benamar S, La Scola B (2016) Clostridium butyricum: from beneficial to a new emerging pathogen. Clin Microbiol Infect 22(1): 37-45.
-
Gaudier E, Jarry A, Blottiere HM, De Coppet P, Buisine MP, et al. (2004) Butyrate specifically modulates MUC gene expression in intestinal epithelial goblet cells deprived of glucose. Am J Physiol Gastrointest Liver Physiol 287(6): G1168-G1174.
-
Hagihara M, Kuroki Y, Ariyoshi T, Higashi S, Fukuda K, et al. (2020) Clostridium butyricum Modulates the Microbiome to Protect Intestinal Barrier Function in Mice with Antibiotic-Induced Dysbiosis. iScience 23(1): 100772.
-
Lee JS, Tato CM, Joyce-Shaikh B, Gulen MF, Cayatte C, et al. (2015) Interleukin-23-Independent IL-17 Production Regulates Intestinal Epithelial Permeability. Immunity 43(4): 727-738.
-
Majewska J, Kaźmierczak Z, Lahutta, K, Lecion D, Szymczak A, et al. (2019) Induction of phage- specific antibodies by two therapeutic staphylococcal bacteriophages administered per os. Front Immunol.
-
Zhang HQ, Ding TT, Zhao JS, Yang X, Zhang HX, et al. (2009) Therapeutic effects of Clostridium butyricum on experimental colitis induced by oxazolone in rats. World J Gastroenterol 15(15): 1821-1828.
-
Leigh BA (2019) Cooperation among conflict: prophages protect bacteria from phagocytosis. Cell Host & Microbe 26(4): 450-452.
-
Keen EC, Dantas G (2018) Close encounters of three kinds: bacteriophages, commensal bacteria, and host immunity. Trends in Microbiology 26(11): 943-954.
-
Chen CC, Kong MS, Lai MW, Chao HC, Chang KW, et al. (2010) Probiotics have clinical, microbiologic, and immunologic efficacy in acute infectious diarrhea. Pediatr Infect Dis J 29(2):135-138.
-
Konstantinov SR, Smidt H, de Vos WM, et al. (2008) S-layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc Natl Acad Sci USA 105(49): 19474-19479.
-
Gluck U, Gebbers JO (2003) Ingested probiotics reduce nasal colonisation with pathogenic bacteria (Staphylococcus aureus, Streptococcus pneumoniae, and β-hemolytic streptococci). Am J Clin Nutr 77(2): 517-520.
-
Bernstein JM, Haase E, et al. (2006) Bacterial interference of penicillin-sensitive and resistant Streptococcus pneumoniae by Streptococcus oralis in adenoid organ culture: Implications for the treatment of recurrent upper respiratory tract infections in children and adults. Ann Otol Belg Laryngol 115(5): 350-356.
-
Di Pierro F, Donato G, Formia F, Adami T, Careddu D, et al. (2012) Preliminary pediatric clinical evaluation of the oral probiotic Streptococcus salivarius K12 in preventing recurrent pharyngitis and/or tonsillitis caused by Streptococcus pyogenes and recurrent acute otitis media. Int J Gen Med 5: 991-997.
-
Luoto R, Ruuskanen O, Waris M, Kalliomäki M, Salminen S, et al. (2014) Prebiotic and probiotic supplementation prevents rhinovirus infections in preterm infants: a randomized, placebo-controlled trial. J Allergy Clin Immunol 133(2): 405-413.
-
Panigrahi P, Parida S, Nanda NC, Satpathy R, Pradhan L, et al. (2017) A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 548: 407-412.
-
De Grandi R, Drago L, Bidossi A, Bottagisio M, Gelardi M, et al. (2019) Putative microbial population shifts attributable to nasal administration of Streptococcus salivarius 24SMBc and Streptococcus oralis 89a. Probiotics Antimicrob Proteins 11: 1219-1226.
-
Campos IB, Darrieux M, Ferreira DM, et al. (2008) Nasal immunization of mice with Lactobacillus casei expressing the pneumococcal surface protein A: induction of antibodies, complement deposition, and partial protection against Streptococcus pneumoniae challenge. Microbes Infect 10(5): 481-488.
-
Kim HJ, Jo A, Jeon YJ, An S, Lee K-M, et al. (2019) Nasal commensal Staphylococcus epidermidis enhances interferon-λ-dependent immunity against influenza virus. Microbiome 7: 80.
-
Bradley KC, Finsterbusch K, Schnepf D, Crotta S, Llorian M, et al. (2019) Microbiota-driven tonic interferon signals in lung stromal cells protect from influenza virus infection. Cell Rep 28: 245-256.e4.
-
Liu Q, Liu Q, Meng H, Lv H, Liu Y, et al. (2019) Staphylococcus epidermidis contributes to healthy maturation of the nasal microbiome by stimulating antimicrobial peptide production. Cell Host Microbe 27: 68–78. e5.
-
Janek D, Zipperer A, Kulik A, Krismer B, Peschel A (2016) High frequency and diversity of antimicrobial activities produced by nasal Staphylococcus strains against bacterial competitors. PLoS Pathog 12: e1005812–e1005820.
-
Latham RD, Gell DA, Fairbairn RL, Lyons AB, Shukla SD, et al. (2017) An isolate of Haemophilus haemolyticus produces a bacteriocin-like substance that inhibits the growth of nontypeable Haemophilus influenzae. Int J Antimicrob Agents 49(4): 503-506.
-
Engen SA, Valen Rukke H, Becattini S, Jarrossay D, Blix IJ, et al. (2014) The oral commensal Streptococcus mitis shows a mixed memory Th cell signature that is similar to and cross-reactive with Streptococcus pneumoniae. PLoS One 9: e104306-e104309.
-
Zipperer A, Konnerth MC, Laux C, Berscheid A, Janek D, et al. (2016) Human commensals producing a novel antibiotic impair pathogen colonization. Nature 535: 511-516.
-
Manning J, Dunne EM, Wescombe PA, Hale JDF, Mulholland EK, et al. (2016) Investigation of Streptococcus salivarius-mediated inhibition of pneumococcal adherence to pharyngeal epithelial cells. BMC Microbiol 16: 225.
-
Bomar L, Brugger SD, Yost BH, Davies SS, Lemon KP (2016) Corynebacterium accolens releases antipneumococcal free fatty acids from human nostril and skin surface triacylglycerols. mBio 7(1): 144-157.
-
Ramsey MM, Freire MO, Gabrilska RA, Rumbaugh KP, Lemon KP (2016) Staphylococcus aureus shifts toward commensalism in response to Corynebacterium species. Front Microbiol 7: 279-294.
-
Santagati M, Scillato M, Patanè F, Aiello C, Stefani S (2012) Bacteriocin-producing oral streptococci and inhibition of respiratory pathogens. FEMS Immunol Med Microbiol 65(1): 23-31.
-
Heng NC, Hammes WP, Loach DM, Tannock GW, Hertel C (2003) Identification of Lactobacillus reuteri genes specifically induced in the mouse gastrointestinal tract. Appl Environ Microbiol 69(4): 2044-2051.
-
Shukla SK, Ye Z, Sandberg S, Reyes I, Fritsche TR, et al. (2017) The nasal microbiota of dairy farmers is more complex than oral microbiota, reflects occupational exposure, and provides competition for staphylococci. PLoS One 12: e0183898.
-
Su Q, Liu Q (2021) Factors affecting gut microbiome in daily diet. Front Nutr 8: 644138.
-
Shanahan F, van Sinderen D, O’Toole PW, Stanton C (2017) Feeding the microbiota: transducer of nutrient signals for the host. Gut 66(9): 1709-1717.
-
Armet AM, Deehan EC, O’Sullivan AF, et al. (2022) Rethinking healthy eating in light of the gut microbiome. Cell Host Microbe 30(6): 764-785.
-
Jardon KM, Canfora EE, Goossens GH, Blaak EE (2022) Dietary macronutrients and the gut microbiome: a precision nutrition approach to improve cardiometabolic health. Gut 71(6): 1214-1226.
-
Gill SK, Rossi M, Bajka B, Whelan K (2021) Dietary fibre in gastrointestinal health and disease. Nature Rev Gastroenterol Hepatol 18: 101-116.
-
Deehan EC, Yang C, Perez-Muсoz ME, Zhang Z, Bakal JA, et al. (2020) Precisionmicrobiome modulation with discrete dietary fiber structures directs short-chain fatty acid production. Cell Host Microbe 27: 389-404.
-
Hughes RL, Kable ME, Marco M, Keim NL (2019) The role of the gut microbiome in predicting response to diet and the development of precision nutrition models. Part II: results. Adv Nutrition 10: 979-998.
-
Costea PI, Hildebrand F, Arumugam M, Backhed F, Blaser MJ, et al. (2018) Enterotypes in the landscape of gut microbial community composition. Nat Microbiol 3: 8-16.
-
Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, et al. (2011) Linking long-term dietary patterns with gut microbial enterotypes. Science 334: 105-108.
-
Abbrescia A, Palese LL, Papa S, Gaballo A, Alifano P, et al. (2014) Antibiotic Sensitivity of Bacillus clausii Strains in Commercial Preparation. Curr Med Chem 1(2): 102-110.
-
Ciffo F (1984) Determination of the spectrum of antibiotic resistance of the “Bacillus subtilis” strains of Enterogermina. Chemioterapia 3: 45-52.
-
Courvalin P (2006) Antibiotic resistance: the pros and cons of probiotics. Dig Liver Dis 38: 261-265.
-
Soren S, Mandal GP, Roy B, Samanta I, Hansda RN (2023) Assessment of Bacillus subtilis based probiotics on health and productive performance of poultry: A review. Indian J Anim Health 62: 2.
- 4th Branchial Cleft Sinus Anomaly Presenting as Recurrent Thyroid Abscess in A Child: A Case Report
- Parotid Duct Injury Repaired Using an Angiocatheter Stent: A Case Report
- Organization and Functionality of the Referral and Counter-Referral System for ENT Disorders in District Hospitals of N’Djamena, Chad: A Cross-Sectional Analytical Study
- Facial Metastases from a Gastrointestinal Stromal Tumor: A Case Report
- Panorama of Ent Cancers and Literature Review: Epidemiological Profile and Therapeutic Management
- Allergic Rhinitis in Senegal: Epidemiological, Clinical and Therapeutic Aspects