Laboratory Resistance Induction of Anopheles Minimus and Anopheles Dirus by the Exposure of Adult Females to Permethrin Treated Mosquito Nettings
A laboratory resistance induction study was conducted to determine the effects of permethrin impregnated nettings on the two major vectors of malaria in Thailand, Anopheles minimums and Anopheles dirus. To simulate the field use of bed nets, only females of the two species were exposed. An. minimus and An. dirus were selected using WHO bioassay tubes and cones, respectively, both using permethrin treated netting materials for three minutes. Changes in the phenotypic expression of resistance were monitored through the baseline and subsequent assessments of susceptibilities from the reference strains and within the generations of selected populations. The effects of permethrin selection on adult size were observed between the reference and last selected generation of both vectors. The activity of detoxifying enzymes (mixed function oxidases and non-specific esterases) were also compared between the reference and selected groups of both species for possible resistance mechanisms. Results showed a decrease in susceptibility level of 1.56 folds from the ninth selected generation (S9 ) of An. minimus. An. dirus developed low level resistance of 4.88 folds at S12. No significant effect was observed on adult sizes of the progenies of selected An. minimus and its reference strain. Varying results however were observed from An. dirus, with body weight significantly increased at S12 while no significant difference was observed between the wing lengths of the selected (S12) and reference strains. Significant increase in the levels of mixed function oxidases were detected from both selected generations of the two species. Further studies are needed to directly determine the presence of the resistance genes and the magnitude by which they are expressed in successive generations of vectors.
Introduction
During those days when malaria was one of the leading causes of mortality and morbidity worldwide, the novel idea of using insecticide treated bed nets to reduce transmission came up. This strategy was envisioned to be the answer to an urgent need for an inexpensive self-help control strategy that would reduce risk of malaria transmission. The World Health Organization (WHO) promoted the idea by including the use of insecticide impregnated bed nets as a part of the personal protection measure in primary health care programs. For several decades now, pyrethroid impregnated bed nets (PIBNs) have been the vector control method of choice due to their long and lasting effects and low mammalian toxicity [1, 2, 3]. Early documented reports attested to PIBNs effectiveness in preventing malaria. Preventive effects decreased the malaria incidence ratio [4] and the number of deaths among children <10 years old [5]. The large-scale trials conducted in China showed PIBNs to be as equally effective as DDT spraying [6]. PIBNs are still considered the most effective method of controlling malaria until now [7, 8]. The wide scale use of factory insecticide pre-treated bed nets now known as long lasting insecticidal nets (LLINs) has resulted in major reduction in malaria transmission. As a result, most countries in Southeast Asia have shifted from control to elimination.
In 2018, the WHO South-East Asian Region estimated 8 million malaria cases [9]. Global cases estimate reached 241million in 2020 with Thailand reporting 2,836 indigenous malaria cases [10]. This record propelled Thailand towards malaria elimination by 2025 [11, 12]. In 1998, Thailand reported 120,000 malaria cases giving an annual parasite index (API) of 2.2 per 1000 population. Anopheles dirus and Anopheles minimus are the two major malaria vectors not only in Thailand but in the greater Mekong region. An. dirus ranked first in terms of vectorial capacity followed by An. minimus [13]. These two species complemented one another to maintain the transmission from forest reservoirs to communities living in forest fringes. An. dirus is sylvatic, mainly exophilic and exophagic but it enters the house to feed on man and leaves soon after [14]. On the other hand, An. minimus is anthropophilic, endophilic and endophagic forest fringe species. Previous vector control measures relied mainly on insecticide residual spraying using DDT [13]. In early 2000, DDT was employed in remote areas at the dosage of 2 g/m2 twice a year while deltamethrin 5% WP was used for the house residual sprays, twice a year in perennial transmission areas. PIBNs were then introduced as a supplementary measure in areas where residual house spraying acceptance was low. In high malaria transmission areas, free nets were then and now being provided. Early on, bed nets used were impregnated with 0.3 g/ m2 of permethrin, twice a year. The use of PIBNs spread all over malarious areas of Thailand as the major vector control measure. With observed continuous reduction of cases, use of the PIBNs was scaled up. Currently, LLINs are commercially available and have replaced conventional treated bed nets. These LLINs exhibit safety and long residual efficacy of 2-3 years or for about 20 washes [15].
It has been theoretically believed that a steep increase in the number of people protected with any insecticide- based vector control measure will likely result in increased selection pressure due to insecticides. Pyrethroid resistance genes (in Anopheles vectors) of various protective capacities can be expected to ascend, thus the main concern then is to detect them at an early stage [16]. Minimal selection pressure is expected with the use of PIBNs. Only the female mosquitoes with endophagic and endophilic behavior are expected to be in contact with the insecticide treated mosquito nets, halving the degree of the selection pressure compared when the selection is directed towards the larval or adult stages that may kill or select for both sexes. This study therefore was designed to evaluate in the laboratory probable development of resistance and its expression (susceptibility/resistance levels) after the intergenerational selection exposures to permethrin treated nettings. Back then, this study was expected to provide insights in the scaled-up and prolonged use of PIBNs on the resistance development in the two malaria vectors. Ideally, the study should reflect the intergenerational period for the resistance to develop while possible resistance mechanism(s) could be detected. Identification of the biochemical mechanisms will allow early detection of resistance using advanced techniques (biochemical and molecular), thereby providing information on appropriate choice of alternative vector control compounds for future use. The identification of resistance mechanisms indicator will help determine the cross-resistance spectrum and will allow mapping of areas with resistant populations.
The objectives of the study were:
- To determine whether the selection with permethrin impregnated bed net will result in tolerance or resistance in the Anopheles mosquito populations,
- To determine the possible after exposure effects on the adult size and
- To screen for the mechanisms of pyrethroid resistance in the two mosquito species; Anopheles minimus and Anopheles dirus.
Materials and Methods
Collection and Rearing of Anopheles minimus and Anopheles dirus
Using cattle-baited traps [17], fully engorged Anopheles females were collected, segregated and reared at the Department of Medical Entomology, Faculty of Tropical Medicine, Mahidol University. Pure and established strains of Anopheles minimus and Anopheles dirus collected from Kanchanaburi, Ratchaburi, and Phrae provinces were separated for resistance induction purposes of this study. The populations were mass-produced in separate insectary compartments to select for resistance. Mosquito colonies were maintained in the insectary under 80-90% relative humidity at 27oC. Adults were kept in cages (30 x 30 x 30 cm) with circular openings securely tied with sleeves to serve as the door in collecting adults and placing pupae for emergence. Cotton pads soaked in 10% sugar solution and 10% multivitamin solution were provided for the newly emerged mosquitoes.
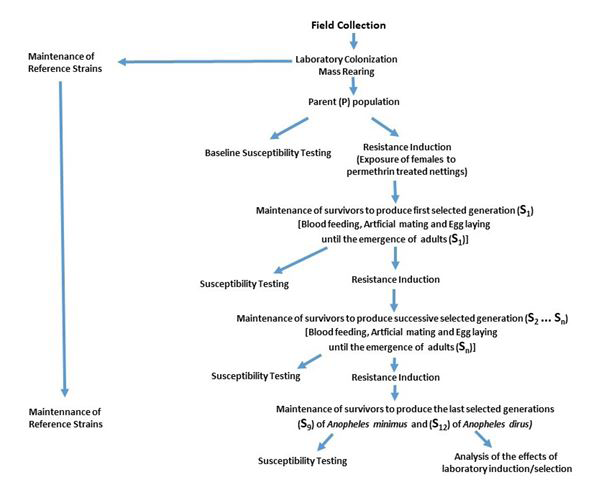
Matured females (3-6 days old) were collected and transferred to separate cups for blood feeding. Fully engorged females were mated following the artificial mating technique described by Ow-Yang CK, et al. [18]. The mass reared populations of An. dirus and An. minimus were divided into halves, one part to be used for susceptibility testing and the other half for resistance induction. The activities and general procedures were presented in the flow charts (Figures 1 & 2). The study was conducted from June 1998 to June 2000 at the Department of Medical Entomology, Faculty of Tropical Medicine, Mahidol University, and Bangkok, Thailand.
Test Insecticide
The insecticide used in the bed net impregnation programs in Thailand at early use of PIBNs was permethrin [13]. Permethrin belongs to the group of pyrethroids which is classified under moderately hazardous category. Permethrin has no unpleasant side effects [19, 20] and has been approved and recommended by the WHO Expert Committee on bed net use [21]. Permethrin used in this study has the following specification: [(3- phenoxy phenyl) methyl 3- (2,2 dichloroethenyl)-2 dimethyl cyclopropane carboxylate] cis: trans isomeric ratio 25:75 w/v emulsifiable concentrate. Welcome Singapore Pte. Ltd.
Net Impregnation
Synthetic nylon netting material (Thai Bednets Manufacture Co. Ltd., 162 Moo 5, Tumbon Pantainorasingha, Amphur Muang, Samutsakorn, Thailand) was used based on the earlier reports that nylon nets produced high mortalities compared to its cotton alternatives and retained insecticides on the surface for a longer duration [22, 23, 24]. To determine the absorption rate of the nylon nets, a measured piece (12 x 15 cm) was dipped into a cylinder filled with a specific volume of water. After complete saturation, the net was removed and the new volume of water was recorded. The difference between the two readings was the amount of water used to dissolve the insecticide and impregnate individual pieces.
The amount/volume of insecticide formulation was calculated using the formula:
( ) ( ) ( )
2 2 Target dose mg of active ingredient ai / m x area m Amount of Insecticide = Insecticide concentration mg ai / ml Insecticide solutions were prepared by adding the calculated amount of the formulation with the required volume of water diluent. Pieces of net were soaked individually in a prescribed volume of insecticide solution in a non-absorbent plastic bag. The individual pieces were rubbed and squeezed to obtain uniform distribution of the insecticide in the nylon net. Impregnated pieces were spread over a plastic mantle indoors to dry overnight. After drying, the impregnated nets were grouped according to insecticide concentration, and were placed individually in plastic bags. The impregnated nets were labeled and were stored in opaque envelopes at room temperature to prevent insecticide degradation.
Susceptibility Testing
Baseline and regular assessments of susceptibilities were done to monitor development of resistance from the reference strain and within the generations of selected populations (Figure 1). World Health Organization (WHO) Standard Susceptibility Procedure [25] was followed to conduct susceptibility tests. Three to four replicates of 15- 25 sugar fed females of each test mosquito species (An. minimus and An. dirus) were collected in individual holding tubes and were kept there for one hour prior to exposure. Afterwards mosquitoes were transferred to exposure tubes containing papers impregnated with a predetermined range of concentrations; 5 concentrations each for An. dirus and An. minimus. The mosquitoes were kept for an hour in the exposure tubes covered with black cloth. Afterwards, the mosquitoes were blown back to the holding tubes and were given sugar solution. Mortalities from each concentration were then recorded after 24 hours. There was no susceptibility test made at S4 and S5 of An. minimus. No susceptibility test was conducted at S3, S5, S7, S9 S10 and S11 due to low population turnout on An. dirus. Table 1 summarizes the susceptibility tests conducted throughout the laboratory inductions done for both of the species.
Selection for Resistance to PIBNs
After the determination of the baseline susceptibility levels, pieces of nylon nets were impregnated with the LC10 to LC50 of the test insecticides in preparation for resistance induction. Selection of An. minimus was done using WHO tube [25] method while An. dirus was done using WHO cone [26]. The initial plan was to determine the possible effects of the two methods in the selection. Due to difficulty in reaching the numbers enough to replicate for both methods, only the use of the above methods were performed. Insufficient numbers did not allow comparison between the two selection methods (tube vs cone). Both species were mass reared to reach population numbers sufficient to start laboratory induction (selection) and conduct baseline and successive susceptibility tests (Figure 1 & Table 1). It took several generations (3-4) to have the induction process started. The generations (An. minimus and An. dirus) first exposed to permethrin treated nettings were considered “parent (P)” while the progenies were considered the first selected generations (S1). Reference strains were grown parallel to the selected generations of An. minimus and An. dirus.
| Laboratory Grown Generations | Anopheles minimus Phrae strain | Anopheles dirus Ratchburi strain | ||
|---|---|---|---|---|
| Selection Conducted | Susceptibility Tests Conducted | Selection Conducted | Susceptibility Tests Conducted | |
| (Yes - √) | (Yes - √) | (Yes - √) | (Yes - √) | |
| (No - X) | (No - X) | (No - X) | (No - X) | |
| Parent (P) | √ | √ | √ | √ |
| First Selected (S ) 1 | √ | √ | √ | √ |
| S 2 | √ | √ | √ | √ |
| S 3 | √ | √ | X | X |
| S 4 | X | X | √ | √ |
| S 5 | X | X | √ | X |
| S 6 | √ | √ | √ | √ |
| S 7 | √ | √ | √ | X |
| S 8 | √ | √ | √ | √ |
| S 9 | X | √ | √ | X |
| S 10 | -- | -- | √ | X |
| S 11 | -- | -- | √ | X |
| S 12 | -- | -- | X | √ |
Table 1: Summary of laboratory inductions (selections) and susceptibility tests conducted for Anopheles minimus and Anopheles dir
Table 1: Summary of laboratory inductions (selections) and susceptibility tests conducted for Anopheles minimus and Anopheles dirus For An. minimus, LC10 (20 mg-ai/m2) was used to induce resistance for the first four generations (P, S1 to S3). Replicates of 25 females were exposed for 3 minutes inside WHO susceptibility tubes with treated net pieces on top of clean filter papers [25]. Survivors were grown continuously. At the start of selection, female mosquitoes were exposed to treated bed nets before giving blood meal and followed by artificial mating. However, direct exposure (selection before blood feeding) reduced the population drastically specially An. minimus Kanchanaburi strain. The colony was wiped out eventually after a few generations probably due to selection pressure that caused feeding inhibition (summarized in the results) and laboratory induction procedure proceeded with only An. minimus Phrae strain. No selection was made at the fourth (S4) and fifth (S5) generations due to low population of test mosquitoes. After the An. minimus population gained sufficient numbers at S6, selection continued with Phrae strain. Modification to the methodology was made by feeding females first with blood and only those that were fully engorged were exposed or selected. Blood meals were given earlier to increase the strength of the population and recover more survivors after exposure, using LC50 (29 mg- ai/m2) this time to increase pressure to see if it would likely result in the development of resistance. Only An. minimus Phrae strain was successfully maintained and selected for seven times within nine generations that was described above (Table 1).
For An. dirus Ratchaburi strain, blood fed females were selected using WHO bioassay test cones [26]. The cones were fastened to a piece of nylon netting material impregnated with 6.25 mg-ai/m2 of permethrin. Replicates of 25 female three to seven days old mosquitoes were aspirated into the test cones. After three minutes, the mosquitoes were placed back on their respective cups used to initially collect each batch. After 24 hours, surviving females were given a blood meal and mated using the artificial mating technique described earlier. The same procedure was followed for each generation of selection. Continuous selection was done except at S3 due to insufficient number of mosquitoes produced from the previous selection (Table 1) thereby completing 11 selections throughout the 12 continuous generations of An. dirus. Susceptibility tests were carried out starting with the parent, and succeeding selected generations: S1, S2, S4, S6, S8 and S12 (Table 1).
Effects of Selection on Adult Size
A minimum of 10 females were randomly selected to compare adult size. Female mosquitoes were fasted for 12 hours and body weights were measured individually using analytical balance. After the body weight was measured, mosquitoes were anaesthetized. Wings were removed and temporarily mounted on a slide using distilled water. After the water had dried and the wing had adhered to the slide, wing lengths were measured from the axillary incision to the apical margin excluding the fringe [27].
Effects of Selection on the Activity of Detoxifying Enzymes
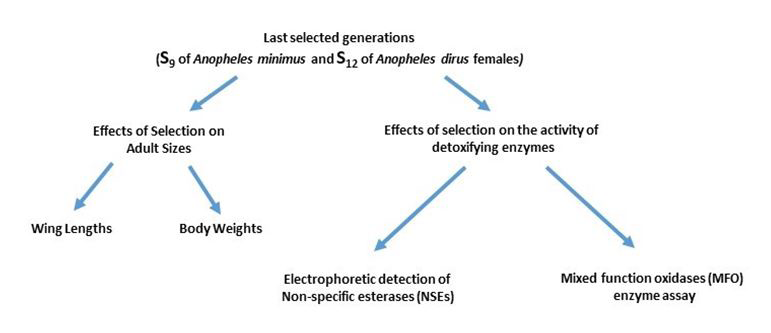
Progenies from the last selected generations (S12 for An. dirus and S9 for An. minimus) and reference (unselected) strains were used in all the biochemical assays (mixed function oxidase and esterase assays) (Figure 2).
Mixed Function Oxidases (MFOs) Enzyme Assay
To measure the activity of monooxygenases from individual insect samples, the procedure described by Vulule J, et al. [28] was adopted. Individual mosquitoes were homogenized in 100ul of potassium phosphate buffer (90 mg Na2HPO4 and 34 mg KH2PO4 in 10 ml of distilled water) adjusted to pH 7.0. The homogenates were diluted with an additional 400 ul of this buffer. Activity of the homogenates from the selected strains and unexposed strain were compared.
A 0.16-mol/liter solution of 3, 3’, 5, 5’- tetramethyl dihydrochloride (TMBZ) was prepared by dissolving 50 mg TMBZ in 25 ml of methanol. Later this solution was diluted with 75 ml of 0.25 mol/liter sodium acetate buffer with pH 5. 200 ul of TMBZ solution was added to the 100 ul of mosquito homogenate in each well followed by 25 ul of 3.0% hydrogen peroxide. The plates were read after 10 minutes using an Immunoassay Reader under 620 nm wavelengths. Optical density (OD) readings were compiled for analysis. Association of heme peroxidase with monooxygenase levels in mosquitoes was earlier demonstrated in Anopheles albimanus pyrethroid resistant strain [29]. A total of 87 and 117 whole mosquito samples for the reference and selected strains respectively were used in the MFO assay.
Electrophoretic Detection Non-Specific Esterases (NSEs)
Randomly selected adult mosquito samples from the reference and last selected generations were examined using vertical polyacrylamide gel slab electrophoresis. About 30 samples were analyzed for each of the normal and selected strains for An. dirus while 50 samples of each strain were analyzed from An. minimus. The methodology published by Apiwathnasorn, et al. and Sucharit, et al. was followed [30, 31]. The loci were designated based on their relative mobilities. The designation is done to allow differentiation of proteins or isoenzymes from different samples across certain localities. The process was made to facilitate characterization of resistance mechanisms. The isoenzyme with the least anodal migration was designated locus 1 and the rest followed. Comparison between the activities of the enzymes present was also done by noting the staining intensities and frequency of the occurrence of the bands. A total of 71 and 96 whole mosquito samples for the reference and selected strains respectively, were used in the NSE assay.
Data Analysis
Susceptibility Tests: The Probit Analysis program developed by Raymond M, et al. [32] was used to analyze mortality data. This computer program was based on Probit Analysis developed by Finney JD, et al. [33]. The resistance ratio (RR) was calculated by dividing the LC50 of the selected strain by the LC50 obtained from baseline susceptibility testing.
Adults Size: Body weight and wing length measurements from the reference and selected strains of both An. minimus and An. dirus were compared using Student T-test for Independent samples by SPSS 7.5. (1997 Copyright, SPSS Inc.).
MFO Assays: Values from optical density (OD) readings were compared between the reference strain and last selected generation using Student T-test for independent samples of both species of An. minimus and An. dirus.
Effects of Selection on Susceptibility
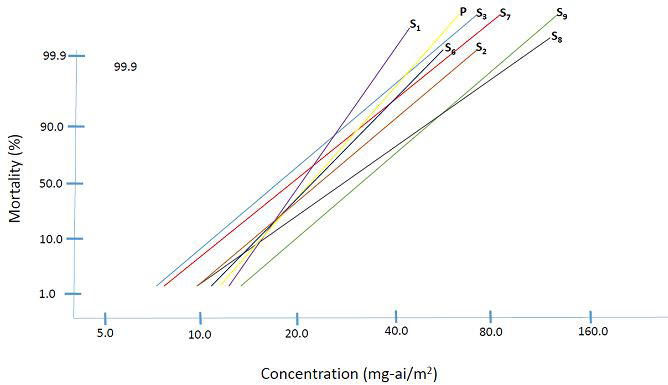
Anopheles minimus: Preliminary experiments showed variations in the effects of exposure on An. minimus Kanchanaburi strain to pyrethroid impregnated nets (LC10) starting from the parent to the third selected generation. Briefly, based on probit analysis of the baseline susceptibility tests, the expected mortality when exposed to LC10 was 8.11% to 15.10% after 24 hours. However, the three-minute exposure to PIBNs affected blood feeding of the females. Only 23.60 to 41.67% fed fully; 19.25 to 43.57 % fed partially while 8.85 to 33.54 % did not feed at all (data not shown). Such that an additional mortality of 2.07 to 13.66 % was recorded after 48 hours. The normal strain showed 81.44% of fully engorged females. This value is considerably high compared to all the values obtained from exposed females starting from the parent to the third selected generation. Mosquito exposure to PIBNs decreased drastically the selected population. Continuous selection was only done in An. minimus Phrae strain. Other collections from Ratchaburi and Kanchanaburi failed to thrive under laboratory selection conditions. Seven selections made within nine generations of continuous colonization of An. minimus Phrae strain showed only slight development of tolerance (1.56 folds) at the last selected generation (S9) (Tables 1& 2, Figure 3). Initial selections (S1 to S2) decreased susceptibility very slightly (RR50 = 1.18) but became even more susceptible at S3. (RR50 = 0.87). There were no selection and susceptibility test done at S4 and S5 due to insufficient numbers. Following the release of selection pressure (no exposure to PIBNs) and blood feeding, the population regained almost equal susceptibility as the parent based on the LC50 values (RR50 = 1.07) calculated from S6. The susceptibility even increased at S7 (RR50 = 0.96) but started to decrease again at S8 (RR50 = 1.41) and S9 giving a final tolerance level of less than two folds (RR50 = 1.56). X2 values showed that responses of the parent from the susceptibility test did not follow exactly a linear pattern which may indicate initial vigor at the start of the colonization (Figure 3). Succeeding generations of exposure however resulted in more linear responses.
| Strain | LC 50 | LC 90 | RR 50 | Slope +SD | X2* |
|---|---|---|---|---|---|
| (mg-ai/m2) | (mg-ai/m2) | ||||
| Parent | 28.85 | 45.96 | - | 6.34+2.18 | 23.41 |
| S 1 | 28.15 | 40.01 | 0.98 | 8.39+1.19 | 2.03 |
| S 2 | 33.99 | 59.21 | 1.18 | 5.32+1.09 | 1.59 |
| S 3 | 25.14 | 44.01 | 0.87 | 5.27+0.87 | 1.1 |
| S 6 | 30.81 | 49.42 | 1.07 | 6.25+0.97 | 5.06 |
| S 7 | 27.79 | 50.54 | 0.96 | 4.94+0.86 | 0.32 |
| S 8 | 40.8 | 78.86 | 1.41 | 4.48+1.21 | 10.26 |
| S 9 | 45.05 | 78.82 | 1.56 | 5.28+0.76 | 5.22 |
Table 2: Probit mortality data from parent and selected generations of Anopheles minimus. Data were analyzed by Probit Analysis p
* X2> 10 not well represented by a line Table 2: Probit mortality data from parent and selected generations of Anopheles minimus. Data were analyzed by Probit Analysis program Raymond M, et al. [32].
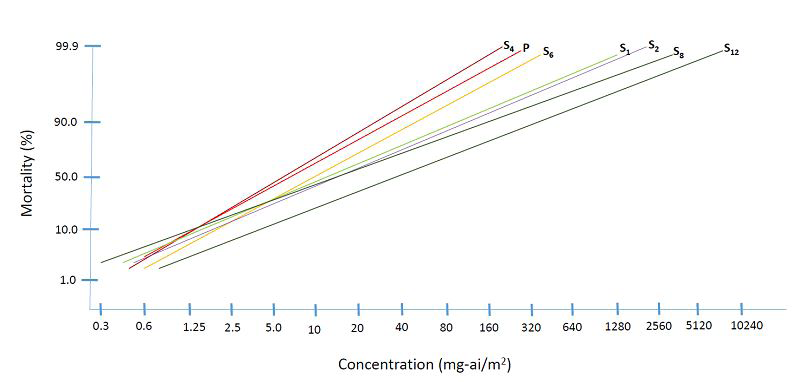
Anopheles dirus: Eleven generations of selection (except at S3) in 12 generations of colonization of An dirus Ratchaburi strain resulted in low level resistance of 4.88 folds at the last selected generation (S12). Parental strain gave an LC50 of 11.39 mg-ai/m2 and LC90 of 68.53 mg-ai/m2. Parental exposure until S2 showed decreased susceptibility based on the resistance ratio calculated from LC50. Absence of selection pressure at S3 due to insufficient number of mosquitoes reverted susceptibility to a level almost equal to the parent strain (RR50 = 1.51) (Table 3 & Figure 4).
| Strain | LC 50 | LC 90 | RR 50 | Slope +SD | X2* |
|---|---|---|---|---|---|
| (mg-ai/m2) | (mg-ai/m2) | ||||
| Parent | 11.39 | 68.53 | _ | 1.64+0.35 | 27.92 |
| S 1 | 17.22 | 191.36 | 1.51 | 1.23+0.45 | 16.58 |
| S 2 | 20.16 | 221.98 | 1.77 | 1.23+0.50 | 39.13 |
| S 4 | 17.22 | 60.94 | 1.51 | 2.34+0.55 | 12.63 |
| S 6 | 14.87 | 96.42 | 1.31 | 1.58+0.51 | 33.92 |
| S 8 | 24.63 | 397.57 | 2.16 | 1.06+0.59 | 47.71 |
| S 12 | 55.55 | 922.1 | 4.88 | 1.05+0.16 | 5.05 |
Table 3: Probit mortality data from parent and selected generations of Anopheles dirus. Data analyzed by Probit Analysis program
* X2> 10 not well represented by a line Table 3: Probit mortality data from parent and selected generations of Anopheles dirus. Data analyzed by Probit Analysis program Raymond M, et al. [32].
Continuous selection until S6 showed decreased susceptibility at nearly equal level as S2. However, at S8, tolerance started to appear as indicated by a two-fold resistance ratio and continued increasing until S12. There was no susceptibility test conducted at S3, S5, S7, S9 S10 and S11 of An. dirus due to low population turn out (Tables 1 & 3). Starting with the parent generation, the colony showed consistently variable response to permethrin as indicated by high X2 values obtained. X2 values higher than 10 indicated that mortality data do not exactly follow a linear pattern. Peak variation occurred at S6 (X2 = 33.92) and S8 (X2 = 47.71). Linear response was however obtained at S12 (X2 = 5.05) which may indicate that the population is starting to develop a true resistance trait. The probit lines showed flat slopes (Figure 4).
Effects of Selection on Adult Size
Anopheles minimus. Results showed that seven times of selection for resistance in nine generations of An. minimus do not show any significant effect on both body weight and wing length parameters (p>0.05). Student T-test for unequal variances showed that there is no significant difference between the mean body weights of the reference (0.95 ±0.03 mg) and S9 (1.04 ±0.05mg) females. Same statistical test showed no significant difference between wing lengths of reference (2.18± 0.01mm) and S9 (2.43± 0.02 mm) females.
Anopheles dirus. Varying results were obtained from the two parameters used to compare adult size of An dirus. Student T-test for unequal variances showed that body weight was significantly increased at S12 (1.40±0.30 mg) when compared to the reference (1.30 ±0.30 mg) (p<0.05). However, Student T-test for equal variances showed no significant difference between the wing lengths of S12 (3.55 ±0.01mm) and reference (3.52 ±0.03mm) strains.
Effects of Selection on the Activity of Detoxifying Enzymes
Mixed Function Oxidases (MFOs) Enzyme Assay
Anopheles minimus. Selection for permethrin resistance resulted in increased activity of mixed function oxidase enzymes at S9. Activity of MFOs after 10 minutes (T10) recorded an average 0.3384±0.02 and 0.4283±0.02 OD values for reference and selected strains; respectively (p<0.05).
Anopheles dirus. Selection for permethrin resistance resulted in significant increase in the activity of mixed function oxidase enzymes at S12. After 10 minutes, recorded OD values were 0.22±0.01 and 0.35±0.01 for reference and selected strains, respectively (p<0.05).
Electrophoretic Detection Non-Specific Esterases (NSEs)
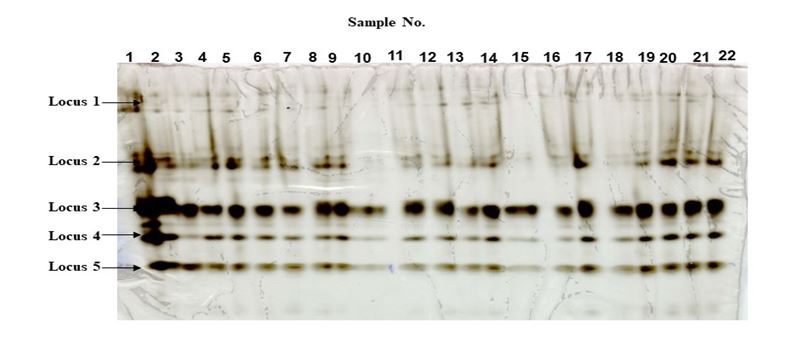
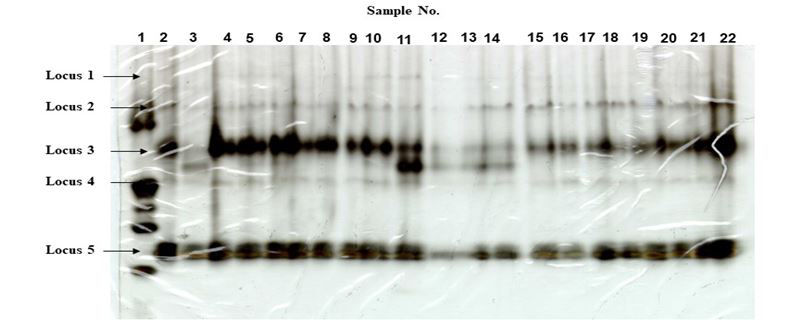
Anopheles minimus. Electrophoretic results showed similar banding patterns for both the reference and selected strains (Figure. 5). The electrophoretic pattern showed presence of five loci producing non-specific esterase enzymes. Loci 1, 2, and 5 were all monomorphic with two bands (alleles) while locus 4 was observed to have only one band. Locus no. 3 showed some specific variations of one or two bands. Locus no.3 also showed visibly higher staining intensity compared to other loci present. No visible difference, however, can be observed in the overall banding pattern from the five loci between reference and selected (S9) strains.
Anopheles dirus. Both the reference and selected strains showed five loci for NSEs (Figure 6). Loci 1, 2 and 4 were monomorphic with a single band. Locus no.5 showed the presence of two bands (alleles). There was no noticeable difference in staining intensities from loci 1, 2, 4 and 5 between the reference and selected strains indicating presence of enzymes almost at the same level. Slightly lower staining intensities of the bands at locus no. 3 of the selected strains may indicate lesser production of enzymes as a result of selection. This may also suggest that selection pressure is directed towards other mechanisms and may indicate non- involvement of non-specific esterases.
Discussion
Effects of Selection on Susceptibility
Preliminary selection exposures showed that three minutes of exposure to low concentration (20 mg-ai/m2) of permethrin resulted in partial female engorgement or total inability to feed the survivors of An. minimus (Kanchanaburi strain) causing failure of the colony. Feeding
inhibition, deterrence and irritation have been shown to be a major mechanism as to how the use of PIBNs contributes to disease reduction [13, 19, 23, 34]. Additionally, altered feeding may result in shorter life span, lower fecundity, irregular and lengthened oviposition cycle of mosquito vectors in communities using PIBNs [35, 36]. This was presented to provide support and further explanation to the published findings regarding the use of PIBNs. The results showed innate toxicity of permethrin towards An. minimus as compared to An. dirus. (Figures 3 & 4) An. minimus showed steeper slopes ranging from 4.48 to 8.39 indicating greater increase in mortality for every ten-fold increase in the logarithmic concentration of permethrin. Flatter slope on the other hand was observed from An. dirus ranging from 1 to 2.34. Theoretically, the population composed of almost entirely susceptible genotypes will produce lines at its steepest. Selection of populations with heterozygous genotypes however will show probit lines decreasing in slope as it moves rightward [37]. This could probably explain variable response and flatter slopes calculated from the probit regression lines of An. dirus.
Results showed that several generations of selection by exposing females alone are needed to induce low level resistance in An. dirus or reduce susceptibility in the case of An. minimus. The experiment showed unique responses of different species to the selection which means individual studies must be made and cannot be directly extrapolated between different species. These findings agree with most laboratory selection experiments undertaken. Laboratory induction experiments of exposing only females of An. maculatus and Aedes aegypti showed only slight development of tolerance after several generations of selection [38]. Pyrethroids however were simultaneously used for both crop protection and public health purposes. The wide scale and continued use of insecticides for both purposes has posed intensive selection pressure on Anopheles vectors. This has resulted in the high levels of deltamethrin resistance in An. sinensis populations from China and Korea [39, 40, 41, 42]. Resistance induction in the larval stage or adult stage exposing both sexes on the other hand showed considerable success. These successful resistance induction experiments done in the laboratory preceded discoveries of resistance in field populations of both anopheline and culicine mosquitoes [13, 43, 44]. In virtually all of the above-cited cases, dosage- mortality line moves to the right in successive generations and consequently LC50 levels increase, depicting similar trends as in An. minimus and An. dirus.
Effects of Selection on Adult Size
No significant effect was observed on adult sizes of the progenies of selected An. minimus (S9) and reference strains. Varying results however were observed from An. dirus with body weight significantly increased at S12 compared to the reference strain while no significant difference was observed between the wing length measurements of reference and selected (S12) strains. Theoretically, adult size is primarily determined by larval nutrition and environmental conditions. With a standardized procedure, changes may occur as a functional adaptation; probably as part of resistance development. Body size can also be associated with survival and dispersal capabilities of some mosquito species [45]. Further work must be done to determine how the change in size due to insecticide selection may affect fitness of the vectors.
Effects of Selection on the Activity of Detoxifying Enzymes
Mixed Function Oxidases: Results show that there is a significant increase in mixed function oxidase activity for both species of An. minimus and An. dirus. An. dirus showed low level resistance of 4.88 folds while An. minimus only showed reduced susceptibility (1.56 folds). Selection exposures to permethrin may have resulted in the increased production of the MFOs but further studies are needed to determine if this mechanism contributes to the development of resistance. MFOs have been reported as probable resistance mechanisms of An. darlingi for deltamethrin and DDT [46] as well as contributing resistance mechanisms for Anopheles as An. gambiae [47] and other Aedes vectors as well [48]. Nonspecific Esterases Non-specific esterases (NSEs) have been reported to have evolved as one mechanism of pyrethroid resistance in An. albimanus from Mexico [49]. This study revealed that there is no significant difference in the patterns and visible staining intensities of the bands obtained from polyacrylamide electrophoresis of non-specific esterases for An. minimus. However for An. dirus, slightly lower staining intensity was observed from locus no.3 of the selected strain. Lower staining intensity indicates reduced amount of enzyme which may or may not be linked to permethrin selection. Further studies are needed for a more conclusive outcome. Like MFOs, NSEs have been implicated as resistance mechanisms of Anopheles [46, 47, 49] and Aedes vectors [45, 48] to several classes of insecticides.
Conclusion
Seven selections made within nine generations of An. minimus Phrae strain resulted in reduced susceptibility (1.56 folds) at the last selected generation (S9). Eleven selections made within 12 generations of An. dirus resulted in the development of low-level resistance of 4.88 folds at the last selected generation (S12). Permethrin selection significantly increased An. dirus adult size based on the body weights but not in terms of wing lengths. Significant increase in the levels of the mixed function oxidases (MFOs) enzymes were both detected from selected strains of An. minimus and An. dirus. Increased MFOs may be responsible for the reduced susceptibility of An. minimus and low-level resistance of An. dirus. Further studies should be conducted to characterize resistance development using molecular biology tools to better predict the manifestation. Certainty on the presence of resistance genes will facilitate determination of the degree and expression of resistant phenotype in filial generations through laboratory induction approximating probable appearance in the field populations.
The limitation of the study includes use of laboratory grown population implying limited population gene pool when compared to the actual wild populations of the species. It is also recommended that WHO impregnated papers be used in the susceptibility tests to ensure adherence and comparability to the standards.
Conflict of interest: The authors declare that they have no conflict of interests.
References
-
Kadota T, Okumo Y, Kohala H, Miyamota J (1976) Mammalian toxicological study of permethrin. Insect Control Science Institute 41(4): 143-151.
-
Schreck CE, Haile DG, Kline DL (1984) The effectiveness of permethrin and deet, alone or in combination, for protection against Aedes tritaeniorhynchus. Am J Trop Med Hyg 33(4): 725-730.
-
Lim SS, Fullman N, Stokes A, Ravishankar N, Masiye F, Murray CJ, Gakidou E (2011) Net benefits: a multicountry analysis of observational data examining associations between insecticide-treated mosquito nets and health outcomes. PLoS Med 8(9): e1001091.
-
Choi HW, Breman JG, Teutsch SM, Liu S, Hightower AW, et al. (1995) The effectiveness of insecticide-impregnated bed nets in reducing cases of malaria infection: a meta- analysis of published results. Am J Trop Med Hyg 52(5): 377-382.
-
Aikins MK, Fox-Rushby J, DAlessandro U, Langerock P, Cham K, et al. (1998) The Gambian National Impregnated Bednet Programme: costs, consequences and net cost effectiveness. Soc Sci Med 46(2): 181-191.
-
Xu B, Xiao X, Weber RH, Lines JD (1998) Comparison of the effect of insecticide treated bednets and DDT residual spraying on the prevalence of malaria transmitted by Anopheles arthropophagus in China. Trans R Soc Trop Med Hyg 92(2): 135-136.
-
Wilson AL, Courtenay O, Kelly-Hope LA, Scott TW, Takken W, et al. (2020) The importance of vector control for the control and elimination of vector-borne diseases. PLoS Neglected Tropical Diseases 14(1): e0007831.
-
Killeen GF (2020) Control of malaria vectors and management of insecticide resistance through universal coverage with next-generation insecticide treated nets. Lancet 395(10233): 1394-1400.
-
(2019) Malaria on the decline in WHO South-East Asia Region; efforts must continue as risks persist: World Health Organization.
-
(2021) World malaria report 2021. World Health Organization.
-
Sudathip P, Kongkasuriyachai D, Stelmach R, Bisanzio D, Sine J, et al. (2019) The investment case for malaria elimination in Thailand: a cost beneft analysis. Am J Trop Med Hyg 100(6): 1445-1453.
-
Lertpiriyasuwat C, Sudathip P, Kitchakarn S, Areechokchai D, Naowarat S, et al. (2021) Implementation and success factors from Thailand’s 1-3-7 surveillance strategy for malaria elimination. Malaria Journal 20(1): 201.
-
Prasittisuk M, Prasittisuk C, Pothichiti V, Aum-aung B, Mongklangkul P (1996) The effect of pyrethroid impregnated mosquito nets on field malaria vector population in experimental huts and individual local houses. Southeast Asian J Trop Med Public Health 27(3): 610-616.
-
Ismail IAH, Phinichpongse S, Boonrasri P (1978) Responses of Anopheles minimus to DDT residual spraying in a cleared forested foothill area in central Thailand. Acta Tropica 35(1): 69-82.
-
Dev V, Raghavendra K, Singh SP, Phookan S, Khound K, et al. (2010) Wash resistance and residual efficacy of long- lasting polyester netting coated with α-cypermethrin (Interceptor) against malaria-transmitting mosquitoes in Assam, northeast India. Transactions of the Royal Society of Tropical Medicine and Hygiene 104(4): 273- 278.
-
Magesa SM, Aina O, Curtis CF (1994) Detection of pyrethroid resistance in Anopheles mosquitoes. Bulletin of the World Health Organization 72(5): 734-740.
-
Choochote W (2011) Evidence to support karyotypic variation of the mosquito, Anopheles peditaeniatus in Thailand. Journal of Insect Science 11(1): 1-12.
-
Ow-Yang CK, Sta Maria FL, Wharton RH (1963) Maintenance of laboratory colony of Anopheles maculatus by artificial mating. World Health Organization (23): 34- 35.
-
Snow RW, Rowan KM, Greenwood BM (1987) A trial of permethrin treated bed nets in the prevention of malaria in Gambian children. Trans R Soc Trop Med Hyg 81(4): 563-567.
-
Yap HH (1986) Effectiveness of soap formulations containing deet and permethrin as personal protection against outdoor mosquito in Malaysia. J Am Mosq Control Assoc 2(1): 63-67.
-
(1996) Report of the WHO Informal Consultation on the Evaluation and Testing of Insecticides. World Health Organization.
-
Lines JD, Myamba J, Curtis CF (1987) Experimental hut trials of permethrin-impregnated mosquito nets and eave curtains against malaria vectors in Tanzania. Med Vet Entomol 1(1): 37-51.
-
Rozendaal JA, Curtis CF (1989) recent researches on impregnated mosquito nets. J Am Mosq Control Assoc 5(4): 500-507.
-
Sexton JD, Ruebush TK, Brandling-Bennett AD, Breman JG, Roberts JM, et al. (1990) Permethrin-impregnated curtains and bed-nets prevent malaria in western Kenya. Am J Trop Med Hyg 43(1): 11-18.
-
(2013) Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. World Health Organization.
-
(2013) Guidelines for laboratory and field-testing of long-lasting insecticidal nets. World Health Organization.
-
Eritja R (1996) Wing biometry and statistical discriminant analysis as a technique to determine sex of a Culex pipiens (Dipters: Culicidae) gynandromorph. Journal of Economic Entomology 89(5): 1338-1341.
-
Vulule JM, Beach RF, Atieli FK, Roberts JM, McAllister JC, et al. (1999) Elevated oxidase and esterase levels associated with permethrin tolerance in Anopheles gambiae from Kenyan villages using permethrin impregnated nets. Med Vet Entomol 13(3): 239-244.
-
Brogdon WG, McAllister JC, Vulule J (1997) Heme peroxidase activity measured in single mosquitoes identifies individuals expressing an elevated oxidase for insecticide resistance. J Am Mosq Control Assoc 13(3): 233-237.
-
Apiwathnasorn C, Komalamisra N, Vutikes S, Deesin T (1987) Mosquito Survey. In: Sucharit S, et al. (Eds). Practical Entomology Malaria and Filariasis. Bangkok Museum and Reference Center. Faculty of Tropical Medicine, Bangkok, pp: 1-37.
-
Sucharit S, Komalamisra N, Leemingsawat S, Apiwathnasorn C, Thonrungkiat S (1988) Population genetic studies on Anopheles minimus complex in Thailand. Southeast Asian J Trop Med Public Health 19(4): 717-723.
-
Raymond M (1993) PROBIT CNRS-UMII. License L 93019. Avenix, 34680. St. George d’Orques, France.
-
Finney JD (1971) Probit Analysis. 3rd (Edn), Cambridge. The University Press.
-
Pleass RJ, Armstrong JRM, Curtis CF, Jawara M, Lindsay SW (1993) Comparison of permethrin treatment for bed nets in The Gambia. Bulletin of Entomological Research 83(1): 133-140.
-
Charlwood JD, Graves PM (1987) The effect of permethrin-impregnated bednets on a population of Anopheles farauti in coastal Papua New Guinea. Med Vet Entomol 1(3): 319-327.
-
Hii J, Alexander N, Chee KC, Rahman HA, Safri A, et al. (1995) Lambdacyhalothrin impregnated bednets control malaria in Sabah, Malaysia. Southeast Asian J Trop Med Public Health 26(2): 371-374.
-
Brown AWA, Pal R (1971) Insecticide resistance in arthropods. 2nd (Edn), World Health Organization.
-
Pant SK (1999) Pyrethroid Resistance in Association with the Use of Insecticides Impregnated Bednets. Unpublished MSc. Thesis. Faculty of Tropical Medicine, Mahidol University, Bangkok, pp: 78.
-
Zhong D, Chang X, Zhou G, He Z, Fu F, et al. (2013) Relationship between knockdown resistance, metabolic Detoxification and Organismal Resistance to Pyrethroids in Anopheles sinensis. PLoS One 8(2): e55475.
-
Kang S, Jung J, Lee S, Hwang H, Kim W (2012) The polymorphism and the geographical distribution of the knockdown resistance (kdr) of Anopheles sinensis in the Republic of Korea. Malaria Journal 11(1): 151.
-
Wang D, Xia Z, Zhou S, Zhou X, Wang R, Zhang Q (2013) A potential threat to malaria elimination: extensive deltamethrin and DDT resistance to Anopheles sinensis from the malaria-endemic areas in China. Malar J 12(1): 164.
-
Hii J, Rueda LM (2013) Malaria vectors in the Greater Mekong Subregion: overview of malaria vectors and remaining challenges. Southeast Asian J Trop Med Public Health 44(S1): 73-165.
-
Brealey CJ, Crampton PL, Chadwick PR, Rickett FE (1984) Resistance mechanisms to DDT and transpermethrin in Aedes aegypti. Pesticide Science (15): 121-132.
-
Chadwick PR, Invest JF, Bowron MJ (1977) An example of cross-resistance to pyrethroids in DDT-resistant Aedes aegypti. Pesticide Science 8(6): 618-624.
-
Maciel-de-Freitas R, Codeco CT, Lourenco-de-Oliveira R (2007) Body size-associated survival and dispersal rates of Aedes aegypti in Rio de Janeiro. Med Vet Entomol 21(3): 284-292.
-
Fonseca-Gonzalez I, Quinones ML, McAllister M, Brogdon WG (2009) Mixed-function oxidases and esterases associated with cross-resistance between DDT and lambda-cyhalothrin in Anopheles darlingi Root 1926 populations from Colombia. Mem Inst Oswaldo Cruz 104(1): 18-26.
-
Nwane P, Etang J, Chouaibou M, Toto JC, Koffi A, et al. (2013) Multiple insecticide resistance mechanisms in Anopheles gambiae s.l. populations from Cameroon, Central Africa. Parasites & Vectors 6: 41.
-
Flores AE, Grajales JS, Salaz IF, Gracia GP, Becerra MHK, et al. (2006) Mechanisms of insecticide resistance in field populations of Aedes aegypti (L) from Quintana Roo, Southern Mexico. J Am Mosq Control Assoc 22(4): 672-677.
-
Penilla RP, Rodriguez AD, Hemingway J, Torres J, Arredondo-Jimenez JI, et al. (1998) resistant management strategies in malaria vector mosquito control. Baseline data for a large scale field trial against Anopheles albimanus in Mexico. Med Vet Entomol 12(3): 217-233.
- Intersecting Epidemics and Climate Vulnerabilities in Conflict- Driven Displacement: Epidemiology, Systemic Challenges, and One Health Gaps in South Sudan
- Advancing Domestic Health Financing for Community Health System Sustainability in South Sudan: The Boma Health Initiative Model (2025–2035)
- Prevalence and Correlates of Post-Exposure Prophylaxis Uptake among Men Who Have Sex with Men in Kisumu County, Kenya
- Medical, Ethical, and Legal Conflicts Surrounding Euthanasia in Argentina. Its Global Implications
- Knowledge and Attitude on Menstrual Hygiene among Adolescent Girls Studying in Secondary Level in Public Schools of Chitwan District, Nepal
- Biological Efficacy of an Adulticide Mixture (clothianidin + deltamethrin) as an Indoor Residual Spray against Adult Anopheles flavirostris in Palawan, the Philippines