Glycolysis Levels During Postprandial and During the Physiological and Prolonged Fasting In Different Tissues in the Human Organism
The aim in this reviews is to evaluate the glycolysis levels during as the postprandial, as well as fasting period physiologic and prolonged, considering the metabolic specificity of the following cell or tissues: red blood cell, brain, liver, muscle and fat, focusing the deep metabolic integration that occurs in living organisms. This mini-review is part of chapter publisher by the Rocha et al. 2017 focusing only of part of carbohydrates metabolism.
Introduction
Most carbohydrates from the diet are directly released to the liver by the hepatic portal vein [1, 2, 3, 4]. In general, the concentration of nutrients in the blood is extremely controlled by the liver, which captures and distributes them. The liver will be the organ responsible to maintenance of homeostasis of carbohydrates and degradation by glycolysis pathway up to glycolysis [1, 2, 4, 5, 6]. In the fasting period, glycogen degradation and muscle proteolysis and lipolysis are responsible for maintaining the energy supply in the body .This review analyzes the metabolism of carbohydrates in different cells (red blood cells) and tissues (brain, muscles, liver and adipose tissue) during in the postprandial period and fasting.
Glycolysis Levels During Postprandial and During the Physiological and Prolonged Fasting In Different Tissues in the Human Organism
Carbohydrate Metabolism in the Postprandial and Fasting Periods
The metabolism of glycolytic pathway in the red blood cells is predominantly anaerobic. The red blood cells have not mitochondrial apparatus for the oxidation of nutrients, thus the red blood cells become dependent on the anaerobic. The consumption of glucose in these cells occurs in a constant way and independent of the nutritional profile. The uptake of glucose by the GLUT1 is independent of the presence of insulin (Figure 1). [1, 3, 4, 7]. In these cells, the glycolytic pathway culminates in the constant production of lactate, which will be captured by the liver. The lactate produced by the red blood cells is converted to glucose by hepatic gluconeogenesis and is one of the sources of maintenance Adv Pharmacol Clin Trials
of fasting glycemic. This cycle is denominated Cycle of Cori [1, 3, 4, 6, 8].
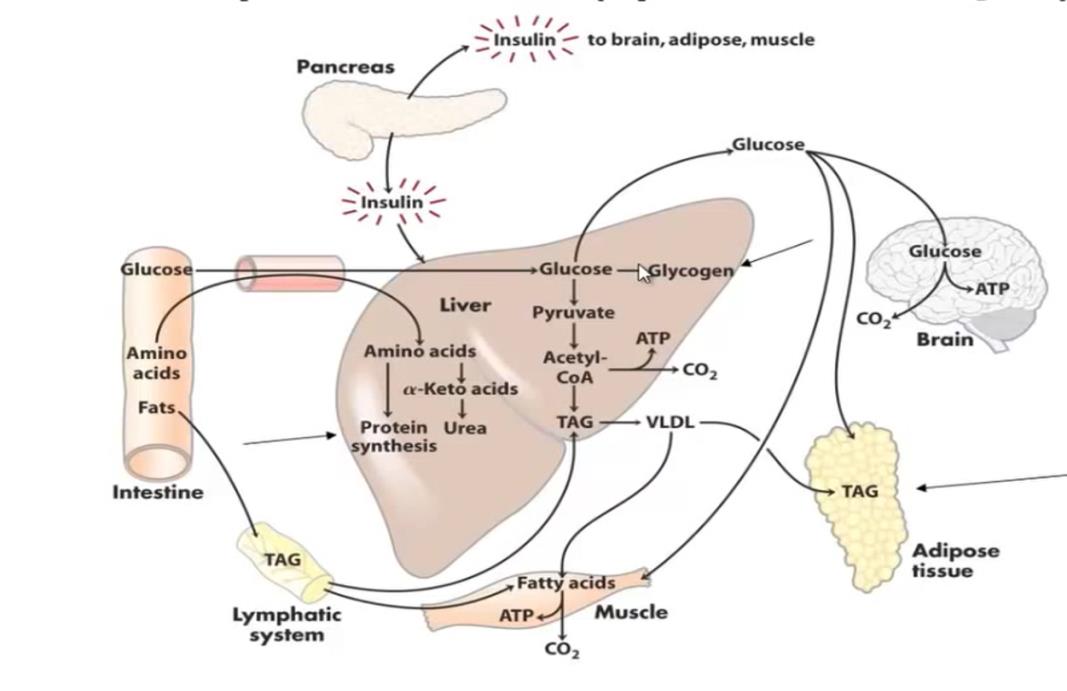
In the physiological situation the consumption of glucose by the CNS reaches 120 g/day, only being lower than the consumption of glucose by the skeletal muscle in physical activity [3]. Most of the energy used by the brain is used in the Na+ and K+ ATPase pump [3], responsible for the repolarization of membrane potential in nerve cells. The CNS, even during sleep, maintains a constant glucose 60% of the total glucose consumed by the rest of the body [9], and is the greatest consumer of this nutrient.
However, the transport of glucose in the liver occurs by GLUT2 transporters, which maintain the concentration of glucose in the hepatocyte in the same proportion that this nutrient exists in the blood circulation by insulin independent manner [1, 3, 4, 7]. However, the glucose can only be used by the hepatic tissue after being phosphorylated. The enzyme responsible for this reaction, a glucokinase, has low affinity for glucose [1, 3, 7], thus, the liver will only phosphorylate and guarantee the permanence of glucose within of liver cells, since there is a sufficiently high concentration of glucose In circulation. This is because the liver can use other energy substrates such as fatty acids or amino acids as an energy source. The high glucose supply, together with the presence of insulin will also stimulate glycogen synthesis, and at this time, the liver becomes a store of glucose [1, 3, 4, 8].
In the other hand, the time of fasting occur glycogenolysis partway process is activated and the liver will export the glucose that had stored in the form of glycogen. Since glycogen is a limited reserve and can only supply the glucose demand in the body for a few hours.
For to supply more source of glycolysis the liver perform the gluconeogenesis [1, 3, 6, 8].
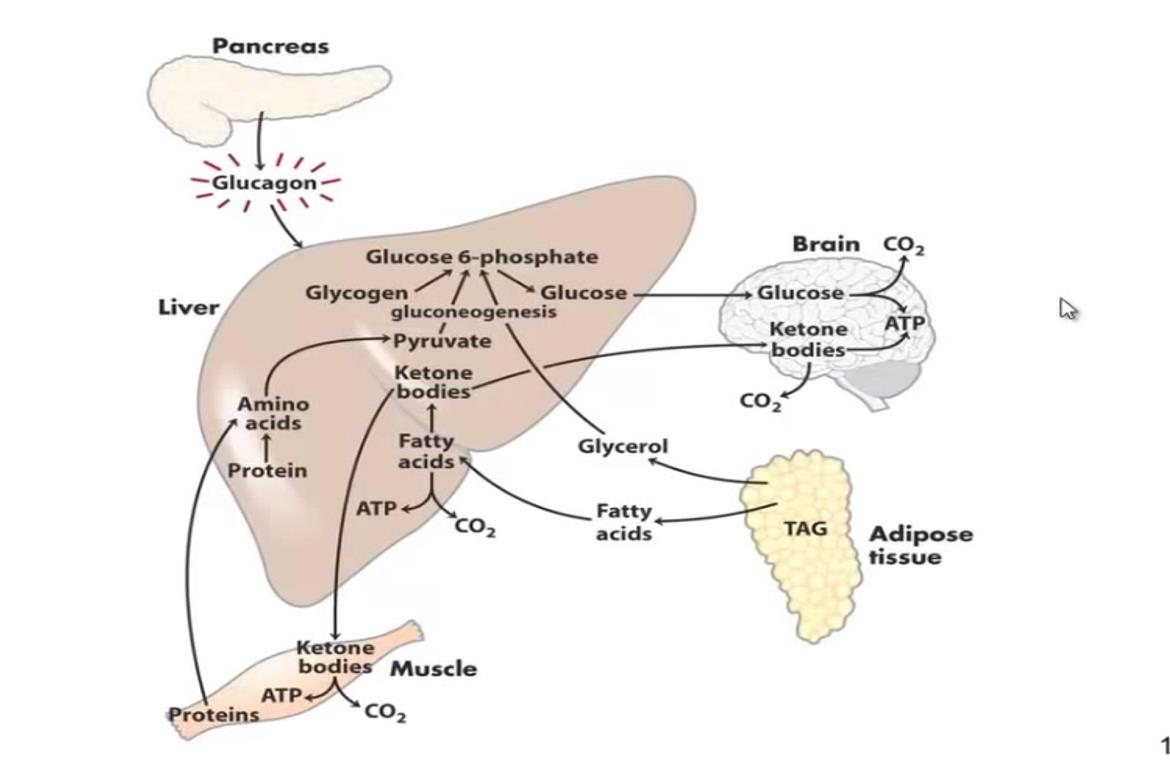
Gluconeogenesis occurs predominantly in the hepatic tissue by the stimulus of glucagon and is simultaneous to hepatic glycogenolysis in depletion mechanism store by the increasement glucagon levels in the blood [1, 3, 4, 8]. While there is glycogen, the rate of gluconeogenesis is small, however, this pathway will occur in the maximum speed after hepatic glycogen depletion [1, 3, 4, 8]. Therefore, in the prolonged fasting, glycemia is maintained only by gluconeogenesis, which means an important metabolic cost, since this pathway is related to the significant loss of muscle mass and adipose tissue that accompany fasting [1, 3, 4, 6, 8]. The synthesis of glucose occurs in the liver during periods of fasting the main precursors are amino acids, skeletal muscle, glycerol, resulting from the mobilization of adipose tissue triglycerides and lactate, coming from the red blood cells, and having as energy source the intense beta-oxidation of the fatty acids released by the mobilization of triglycerides (Figure 2) [1, 4, 8]. Gluconeogenesis may continue to occur in the liver for some time even with the arrival of foods the production of glycogen from amino acids from the diet [10, 11].
As the fasting period increases, hormones like adrenaline and cortisol are secreted, developing the chronically maintenance of glucose homeostasis. These hormones increase the lipolysis, providing fatty acids and glycerol necessary to gluconeogenesis and ketone bodies synthesis [4, 6, 8, 12]. In addition, the body decreases its metabolism is promote the lower release of thyroid hormones (T3 and T4) [13, 14]. With this, the body consumes less energetic substrates but the consumption of glucose by some tissues is greatly diminished as for example the skeletal muscle. This type of adaptation favors the availability of glucose for tissues that use it exclusively as for example erythrocytes and brain [15].
Conclude Remarks
The liver is a center of metabolism and the organ is responsible to maintenance of homeostasis of carbohydrates and degradation by glycolysis pathway up to glycolysis. In the fasting period, glycogen degradation and muscle proteolysis and lipolysis are responsible for maintaining the energy supply in the body. In the postprandial periods occur increase of the glycolysis in the blood is uptakes in the liver. The liver can use other energy substrates such as fatty acids or amino acids as an energy source. The high glucose supply, together with the presence of insulin will also stimulate glycogen synthesis, and at this time, the liver becomes a store of glucose.
In the other hand, the time of fasting occur glycogenolysis partway process that is activated and the liver will export the glucose that had stored in the form of glycogen. Since glycogen is a limited reserve and can only supply the glucose demand in the body for a few hours. For to supply more source of glycolysis the liver perform the gluconeogenesis, a partway almost inverse of glycolysis pathway, except for 3 reaction irreversible in which are of point of regulation for production of glycolysis in the organism for maintenance of glycemic rate in the blood.
Conflicts of Interest
The author declared not possess none conflicts of interesting
References
-
Nelson DL, Cox MM (2014) Principles of Biochemistry of Lehninger. 6th (Edn.), Freeman and Company.
-
McPherson RA, Pincus MR (2011) Henry's Clinical Diagnosis and Management by Laboratory Methods. 22nd (Edn.), Elsevier Saunders.
-
Voet D, Voet JG, Pratt CW (2015) Fundamentals of Biochemistry. 5th (Edn.), Wiley.
-
Malheiros SVP (2006) Integração metabólica nos períodos pós-prandial e de jejum - um resumo J Biochem Educ 1: C1-C7.
-
Brosnan JT (2000) Glutamate, at the interface between amino acid and Carbohydrate metabolism. J Nutr 130(4S Suppl): 988S-990S.
-
Schutz Y (2015) Protein turnover, ureagenesis and gluconeogenesis. Inter J Vit Nutr Res 81: 101-107.
-
Adeva-Andany MM, González-Lucán M, Donapetry- García C, Fernández-Fernández CF, Ameneiros- Rodriguez E (2016) Glycogen metabolism in humans. BBA Clin 5: 85-100.
-
Rui L (2014) Energy metabolism in the liver. Compr Physiol 4(1): 177-197.
-
Berg JM, Tymoczko JL, Stryer L (2011) Biochemistry 6th (Edn.), Freeeman.
-
Shulman GI, Landau BR (1992) Pathways of glycogen repletion. Physiol Rev 72(4): 1019-1035.
-
Guyton AC, Hall JE (2002) Tratado de Fisiologia Médica. Guanabara Koogan.
-
Finn PF, Dice JF (2016) Proteolytic and lipolytic responses to starvation. Nutrition 22(7-8): 830-844.
-
De Groot LJ (1999) Dangerous dogmas in medicine: the nonthyroidal illness syndrome. J Clin Endocr Metab 84(1): 151-164.
-
Chan JL, Heist K, DePaoli AM, Veldhuis JD, Mantzoros CS (2003) The role of falling leptin levels in the neuroendocrine and metabolic adaptation to short- term starvation in healthy men. J Clin Invest 111(9): 1409-1423.
-
Coelho RG (2016) Integrações do metabolismo em exercício, jejum e no estado alimentado. Revista Ciência Atual 8: 2-9.
- Gallic and Citric Acid Present in the Peels of Tropical Fruits as an Alternative in the Fight against Cancer
- Treating the Forehead Lines with Combination of Forehead and Glabellar Botulinum Toxin Among Japanese Patients
- Clinical Evaluation of Patients Suffering from Breast Cancer & Determination of Treatment Therapies and Better Strategies Related to Breast Cancer
- Medieval Recipes by Al-Zahrāwī for Heart Palpitations Treatment
- Etiology and Prescription Errors of Myocardial Infarction in Different Health Care Systems of Azad Kashmir
- Early Diagnosis and Multidisciplinary Management of Turner Syndrome: A Paediatric Case Study