The Bioactive Flavonoid Taxifolin Inhibits Differentiation and the Production of the Inflammatory Cytokine Interleukin-6 in Cultured Human Adipocytes
Background: Taxifolin, a bioactive flavonoid that possesses potent antioxidant activity, has been reported to show multiple pharmacological properties, including protective effects against obesity-related diabetic nephropathy and diabetic cardiomyopathy. However, knowledge regarding the effects of taxifolin on adipocytes, which are closely associated with obesity and diabetes, is insufficient. Objective: This study aimed to explore the direct effects of taxifolin on differentiation and inflammation adipocytes byculturing human preadepocytes (HPAds). Methods: HPAds were cultured for 16 days in a differentiation medium with or without taxifolin to examine its effect on differentiation. On day 16, levels of lipid and differentiation-related gene expression (PPARγ, C/EBPα, adiponectin, CD36, and GLUT4 mRNAs) in adipocytes were measured using the Oil Red O assay and the real-time polymerase chain reaction assay, respectively. Adiponectin levels in the medium were measured using an enzyme-linked immunosorbent assay (ELISA). Thetaxifolin effect on inflammation was assessed using mature adipocytes differentiated for 15 days. After incubating mature adipocytes in a differentiation medium containing tumor necrosis factor-α (TNF-α) or TNF-α + taxifolin for three hours, the level of interleukin-6 (IL-6), an inflammatory cytokine in the medium, was measured using ELISA. Results: Exposure of taxifolin to adipocytes during differentiation decreased the levels of lipid in adipocytes and adiponectin in the medium. It also decreased the expression levels of C/EBPα, adiponectin, CD36, and GLUT4 mRNAs, but not PPARγ mRNA. Taxifolin inhibited the increase in IL-6 levels in the medium induced by TNF-α in mature adipocytes. Conclusion: These results suggest that taxifolin has anti-differentiation and anti-inflammatory effects on adipocytes. Additionally, taxifolin is expected to have the potential as a therapeutic drug for obesity and metabolic syndrome.
Introduction
Obesity, especially visceral fat obesity, is commonly associated with type 2 diabetes, coronary artery disease, and hypertension, and the coexistence of these diseases is called metabolic syndrome [1, 2]. Obesity is a pathologically excessive accumulation of body fat, and its prevalence has recently increased dramatically [3, 4, 5]. Particularly, the accumulation of visceral fat induces metabolic disorders, such as glycolipids, increasing the risk of developing insulin resistance and diabetes. Additionally, diabetes increases the risk of developing complications, such as diabetic retinopathy, nephropathy, and neuropathy. It also increases the risk of developing life-threatening arteriosclerosis, such as myocardial and cerebral infarctions [6, 7, 8, 9].
Adipose tissue is a storage organ that stores excess energy and an endocrine organ that produces and secretes various adipocytokines. Normal and healthy adipocytes promote glycolipid metabolism [10, 11]. However, obese adipose tissue is excessively enlarged by a combination of increased size (hypertrophy) and number (hyperplasia) of adipocytes [5]. In addition to excessive lipid accumulation, hypertrophic adipocytes produce and secrete proinflammatory cytokines and chemokines, such as interleukin-6 (IL-6) and monocyte chemoattractant protein-1 (MCP-1) [12]. During the development of obesity, macrophage infiltration into adipose tissue increases, and macrophage-derived tumor necrosis factor-α (TNF-α) enhances proinflammatory cytokines production in adipocytes [13, 14, 15]. That is, the macrophage- derived TNF-α activates the nuclear factor-κB (NFκB) in hypertrophic adipocytes and induces the synthesis and secretion of proinflammatory cytokines such as IL-6 and MCP-1 [13, 14, 16] However, the production and secretion of the anti-inflammatory adipocytokine adiponectin, which inhibits the NFκB pathway, is conversely reduced in obese hypertrophic adipocytes [15, 17]. Such regulatory disruption of adipocytokine production in hypertrophic adipocytes is thought to cause chronic inflammatory conditions, leading to insulin resistance and diabetes [6]. Therefore, reducing hypertrophic adipocytes and promoting the normal functions of adipocytes are crucial for treating and preventing various diseases caused by obesity and metabolic syndrome.
Many pharmacological approaches have been reported to treat obesity-related diabetes and insulin resistance. For example, peroxisome proliferator-activated receptor γ (PPARγ) agonists, such as thiazolidine derivatives promote adipocyte differentiation [18, 19, 20], and increase adiponectin production, exerting an anti-inflammatory effect by suppressing the TNF-α-activated NFκB pathway [13, 14, 16]. AdipoRon, an adiponectin receptor agonist, has been reported to bind to the adiponectin receptors AdipoR1 and AdipoR2, and exert antidiabetic effects through activation of AMP-activated protein kinase and PPAR-α pathways, respectively [21, 22]. Conversely, xanthine derivatives, such as theobromine and theophylline, suppress adipocyte differentiation and promote lipolysis by inhibiting adenosine receptor and phosphodiesterase (PDE), causing an anti- obesity effect [23, 24].
Taxifolin (dihydroquercetin), is a bioactive flavonoid found in grapes, citrus fruits, onions, green tea, olive oil, and several herbs, such as milk thistle, French maritime bark, Douglas fir bark, and Smilacis Glabrae Rhizoma [25]. Taxifolin has a strong antioxidant activity and has shown promising pharmacological properties in managing inflammation, tumors, microbial infections, oxidative stress, dementia, cardiovascular, and liver disorders [26]. Recently, in addition to the multiple pharmacological properties, taxifolin was also reported to prevent diabetic nephropathy in rats [27] and diabetic cardiomyopathy in mice [28]. These reports suggest that taxifolin may be effective in diabetes management. However, there are few studies on adipocytes that are closely related to diabetes, but the effects of taxifolin on inflammatory and lipid metabolism in cultured adipocytes and macrophages were reported more recently [29]. The results showed that 1) taxifolin inhibited lipopolysaccharide (LPS)-induced nitric oxide production and tert-butyl hydroperoxide-induced reactive oxygen species production as well as LPS-induced expression of IL-1β and TNF-α mRNAs, in a mouse macrophage-like cell line, RAW264.7 cells. 2) Taxifolin increased the liver X receptor β mRNA expression, involved in preventing hyperlipidemia in macrophage-differentiated THP-1 cells. Additionally, 3) taxifolin reduced lipid accumulation in adipocyte-differentiated 3T3-L1 cells. These results suggest that taxifolin may improve obesity-related oxidative stress, inflammation, and lipid metabolism by acting on macrophages in obese adipose tissue. However, knowledge regarding the direct effects of taxifolin on adipocytes is insufficient. That is, there is only evidence that taxifolin reduced lipid accumulation in 3T3-L1 cells.
Therefore, this study aimed to clarify the direct effect of taxifolin on adipocytes. To clarify the objectives, the effects of taxifolin on adipocyte differentiation and TNF-α-induced
inflammatory cytokine IL-6 using cultured human adipocytes were examined.
Materials and Methods
Test Substances and Agents
Taxifolin (Da Hurian Larch Extract Dihydroquercetin, 98.7% purity, Ametis JSC, Amur Region, Russia) was supplied from DHQ Co., Ltd. (Saitama, Japan). TNF-α and dimethyl sulfoxide (DMSO) were purchased from Fujifilm Wako Pure Chemical Corporation (Osaka, Japan) and Sigma-Aldrich (St. Louis, MO, USA), respectively. Oil Red O and 2-propanol were purchased from Wako (Osaka, Japan) and Sigma-Aldrich (St. Louis, MO, USA). Other chemicals were purchased from commercial sources.
Cell Culture for Preadipocyte Proliferation
Primary human preadipocytes (HPAds) derived from human pericardiac adipose tissue (Cell Applications, Inc., San Diego, CA, USA) were seeded into a 75 cm2 culture flask (Corning, Corning, NY, USA) with the preadipocyte growth medium, including 10% FBS, 10 μg/mL insulin, 100 U/ mL penicillin, 100 μg/ml streptomycin, and 250-ng/ml amphotericin B (Cat No. 811K-500, Cell Applications, Inc), and cultured at 37°C in an atmosphere with 5% CO2 until it attained confluence. During growth culture, the growth medium was changed every two days.
Experimental Designs
Cell Viability: The confluent HPAds(16,500 cells/well) were seeded into a collagen-coated 96-well microplate (AGC TECHNO GLASS Co., Ltd., Shizuoka, Japan) containing preadipocyte growth medium (150 μL/well), and cultured at 37°C and 5% CO2 until 100% confluence. Confluent cells were used to determine adipocytes cell viability exposed to taxifolin for 16 d and three h.
Cytotoxicity for 16 days during differentiation: Differentiation of confluent cells cultured in 96-well microplates were initiated (Day 0) by replacing the growth medium with the adipocyte differentiation medium, including 10% FBS, 5 μg/mL insulin, 100 U/mL penicillin, 100 μg/ml streptomycin, and 250 ng/ml amphotericin B, 0.5 μM dexamethasone, 0.25 mM IBMX (Cat No. 811D-250, Cell Applications, Inc.) or the medium containing various taxifolin concentrations (final concentrations: 0.2, 2, and 10 µg/mL taxifolin dissolved 0.5% DMSO). The cells were cultured at 37°C with 5% CO2 during differentiation. The differentiation medium was changed every three days. On day 16 of differentiation, cell viability was measured using a cell counting kit-8 (CCK-8) assay.
Three hours cytotoxicity against differentiated mature adipocytes on day 15: Confluent HPAds prepared following the procedure described in the previous section were cultured for 15 d in the differentiation medium. On day 15 of differentiation, the culture medium was replaced with a fresh differentiation medium including 0.5% DMSO (control) or the medium containing various concentrations of taxifolin (final concentrations: 0.2, 2, and 20 µg/mL) and TNF-α (5 ng/mL) dissolved in 0.5% DMSO, and then the cells were incubated at 37°C with 5% CO2. After 3h-incubation, cell viability was measured using a CCK-8 assay.
CCK-8 assay: For cell viability determination, 15 μL of the CCK-8 solution (Dojindo Molecular Technologies, Gaithersburg, MD, USA) was added to wells containing cells exposed to taxifolin for 16 days and three hours. The cells were incubated for another 1–2 h at 37°C in a CO2 incubator and measured at 450 nm using a microplate reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA). Cell viability was evaluated as a percentage of the absorbance of drug-treated cells relative to that of the control (100% viability).
Changes in Adipocyte Lipid and Medium Adiponectin Levels during Differentiation
The confluent HPAds (42,000 cells/well) were seeded into a collagen-coated 48-well microplate (AGC TECHNO GLASS Co., Ltd.) containing the growth medium (150 μL/well), and cultured at 37°C with 5% CO2 until 100% confluence.
Differentiation was initiated (Day 0) by replacing the medium of the confluent preadipocytes with the differentiation medium (Cell Applications, Inc.). The cells were cultured at 37°C with 5% CO2 during differentiation, and the differentiation medium was changed every three days. The state of adipocyte differentiation was confirmed by staining adipocytes on days zero, two, seven, 16, 31, and 37 with Oil Red O after removing the medium, and measuring the lipid levels of adipocytes in the wells after microscopic examination. The media obtained on days zero, two, seven, 16, 31, and 37 of differentiation were used for the adiponectin assay.
Effects of Taxifolin on Adipocyte Differentiation
Confluent HPAds prepared according to the procedure described in the previous section were cultured for 16 days in the differentiation medium (control) or the medium containing various taxifolin concentrations (final concentrations: 0.2, 2, and 10 µg/mL) dissolved in 0.5% DMSO. The lipid accumulation in adipocytes and the adiponectin concentration in the medium were measured on day 16 of differentiation using Oil Red O staining and ELISA assays, respectively, to evaluate the effect of taxifolin on adipocyte differentiation. To further investigate the taxifolin effect on adipocyte differentiation, expression of differentiation-related genes [PPARγ, CCAAT/enhancer-binding protein (C/EBPα), adiponectin, cluster of differentiation 36 (CD36), and glucose transporter type 4 (GLUT4) mRNAs] was measured on day 16 in an experiment set separately by the same procedure using a collagen-coated 6-well microplate (AGC TECHNO GLASS Co., Ltd.) seeded with HPAds (440,000 cells/well). Each gene expression was measured using the real-time polymerase chain reaction (PCR) analysis.
Effects of taxifolin on TNF-α-induced proinflammatory cytokine IL-6
The taxifolin effect on TNF-α-induced increase in IL-6 was examined in mature adipocytes on day 15 of differentiation. Briefly, on day 15 of differentiation, the differentiation medium was replaced with a fresh differentiation medium containing TNF-α (final concentration: 5 ng/mL) or TNF-α plus various taxifolin concentrations (final concentrations: 0.2, 2, and 20 µg/mL) dissolved in 0.5% DMSO, and then the cells were incubated for three hours at 37°C with 5% CO2. Controls were similarly cultured in the normal differentiation medium. The aliquot of the culture medium was carefully collected after incubation. The medium IL-6 concentration was measured using ELISA kit.
Measurement of Adipocyte Lipid Stained with Oil Red O
The adipocytes cultured in a well were rinsed twice with 500 μL phosphate-buffered saline after the medium was removed, then 200 μL 0.36% Oil Red O prepared with a 60% 2-propanol was added to stain intracellular lipid for 15 min at room temperature. Then, the Oil Red O solution was removed and the cells were washed thrice with distilled water, after which 250 μL 2-propanol was added to extract the dye by shaking for 30 min. The extracted dye was measured using a microplate reader (Thermo Scientific Multiskan FC, Thermo Fisher Scientific, Inc.) at a 540 nm wavelength.
ELISA Assays for Adiponectin and IL-6
Concentrations of adiponectin and IL-6 in the medium were measured using the AssayMaxTM human adiponectin ELISA kit (Assaypro, LLC, St. Louis, MO, USA) and interleukin-6 ELISA kit (Chondrex, Inc, WA, USA), following the manufacturer’s instructions. The ELISA sensitivity was 0.391 ng/mL for adiponectin and 9 pg/mL for IL-6.
Statistical Analysis
All values were represented as the mean ± S.E.M. Dunnett’s test evaluated the statistical significance after a one-way analysis of variance (ANOVA). The accepted significance level was P < 0.05.
Results
Cell Viability
The non-cytotoxic taxifolin concentration was determined by exposing taxifolin (0.2, 2, 10, and 20 µg/ mL) to cells for 16 d from the onset of differentiation in a differentiation experiment. In anti-inflammatory experiment, taxifolin was exposed to the 15th day-mature cells for 3 h. Cell viability is calculated as the percentage of viable cells in the taxifolin-treated group relative to the control group, and the range of non-toxic concentrations in each experiment is shown in Table 1. The viability of cells exposed to 0.2–10 µg/ mL taxifolin for 16 d was 83.7 ± 3.5%, The viability of mature cells exposed to taxifolin at 0.2–20 µg/mL concentrations for three hours was 107.9 ± 2.2%, respectively. Each experiment was performed using the non-toxic concentration range of taxifolin shown in Table 1.
| Test substance | Exposure time | Concentration (µg/mL) | Cell viability (%) |
|---|---|---|---|
| Taxifolin | Differentiation experiment | ||
| 16 d | 0.2 – 10 | 83.7 ± 3.5 | |
| Anti-inflammatory experiment | |||
| 3 h | 0.2 – 20 | 107.9 ± 2.2 |
The viability of adipocytes treated with various concentrations of taxifolin were measured using CCK-8 assay. Cell viability was calculated as the percentage of viable cells in the taxifolin-treated group relative to the control group. Data represents the mean ± SEM (n = 18).
Changes in Lipid and Adiponectin During Differentiation
Figure 1 shows changes in lipid concentrations in adipocytes and adiponectin in the medium during differentiation. Photographs show that small mature adipocytes gradually increased until day 16 of differentiation, but many hypertrophic adipocytes were observed on day 37. The adipocyte lipid concentration increased linearly with increasing days of differentiation. Contrastly, adiponectin concentration in the medium increased rapidly until day 15 of differentiation, but the increased level almost reached a plateau on day 31 and decreased rather rapidly on day 37.
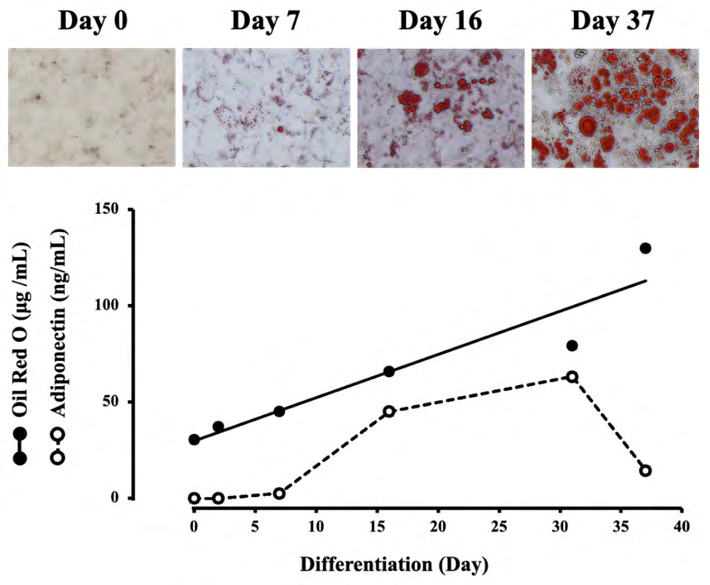
Figure 1: Changes in lipid concentrations in adipocytes and adiponectin concentration in the medium during differentiation. The upper photographs show typical adipocytes stained with Oil Red O on days 0, 7, 16, and 37 of differentiation. The concentrations of lipid and adiponectine were measured on days 0, 2, 7, 16, 31, and 37. Each value represents the mean ± SEM (n = 6).
Effect of Taxifolin on Adipocyte Differentiation
Lipid and Adiponectin
The effect of taxifolin on adipocyte differentiation was assessed by culturing confluent preadipocytes for 16 days in the differentiation medium (control) or medium containing taxifolin. Typical photographs of control and taxifolin-treated adipocytes on day 16 of differentiation are shown in Figure 2. Lipids’ concentration in adipocytes and adiponectin in the medium measured on day 16 as differentiation markers are also shown in Figures 2A and 2B, respectively. In the 0.2 and 2 µg/mL taxifolin-treated groups, mature adipocytes were predominantly observed over immature cells, as in the control group. Conversely, in the 10 µg/mL taxifolin- treated group, immature cells were predominantly observed over mature adipocytes. Concentrations of both Oil Red O-stained adipocyte lipids and the medium adiponectin were significantly decreased by treating with taxifolin (10 µg/mL) for 16 d compared to controls.
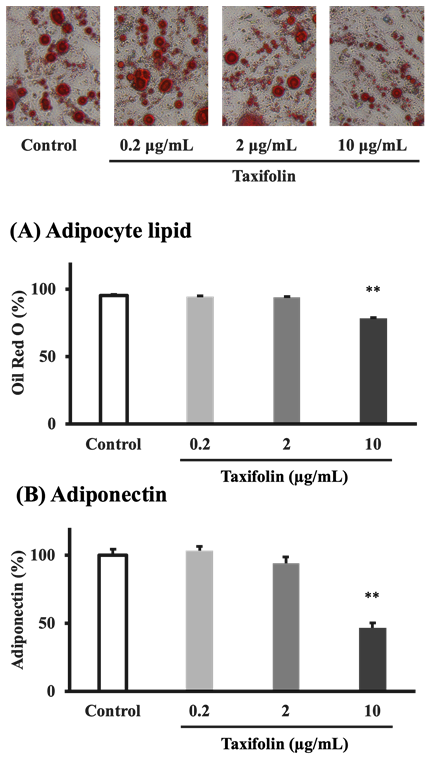
Figure 2: Taxifolin effects on adipocyte differentiation. Confluent preadipocytes were cultured for 16 d in the differentiation medium (control) or medium containing various taxifolin concentrations (final concentrations: 0.2, 2, and 10 µg/mL). The upper photographs show typical control and taxifolin-treated adipocytes on day 16 of differentiation (Oil Red O staining). The concentrations of adipocyte lipid (A) and the medium adiponectin (B) were measured on day16 of differentiation. Each data represents the mean ± SEM (n = 6). **P < 0.01 vs control: statistical significance was assessed using Dunnett’s test following one-way ANOVA.
Differentiation-related genes
To further investigate the taxifolin effect on adipocyte differentiation, expression of differentiation-related genes (PPARγ, C/EBPα, adiponectin, CD36, and GLUT4 mRNAs) was measured on day 16 in an experiment set separately using the same procedure. The results are shown in Figure 3. Taxifolin (10 µg/mL) significantly inhibited C/EBPα mRNA expression, but not PPARγ mRNA expression. The mRNA expression of adiponectin (P < 0.001) and GLUT4 (P < 0.001) was also significantly inhibited by taxifolin (2 and/ or 10 µg/mL). Taxifolin (10 µg/mL) tended to inhibit CD36 mRNA expression, although it was insignificantly different (P = 0.11).
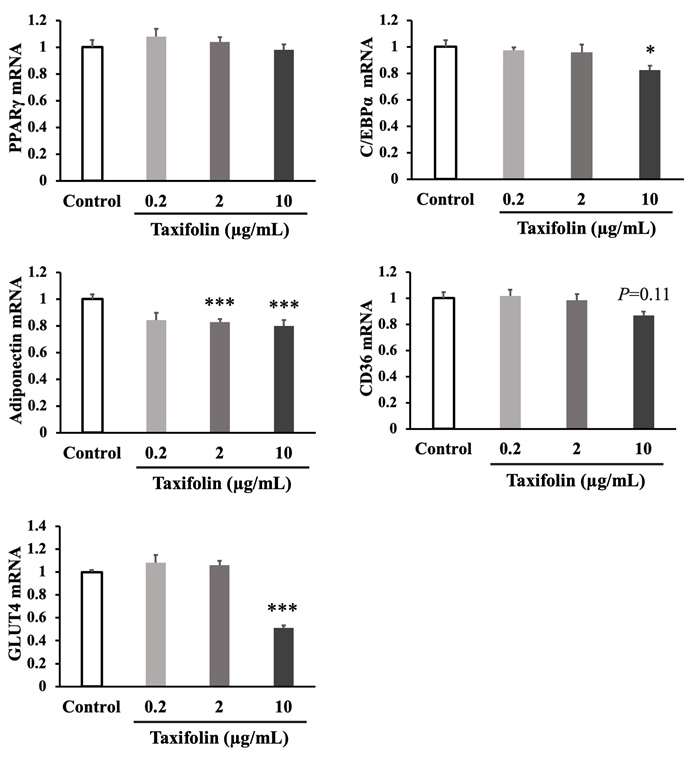
Figure 3: Effects of taxifolin on the expression of differentiation-related genes (PPARγ, C/EBPα, adiponectin, CD36, and GLUT4 mRNAs) in adipocyte differentiation. Confluent preadipocytes were cultured for 16 days in the differentiation medium (control) or medium containing various taxifolin concentrations (0.2, 2, and 10 µg/mL). The quantification of each gene expression in the adipocytes was measured on day 16 using a real-time PCR system. The gene expression level was expressed relative to the control after normalization using GAPDH. Each data represents the mean ± SEM (n = 6). *P < 0.05 vs control, ***P < 0.001 vs control: statistical significance was assessed using Dunnett’s test following one-way ANOVA.
Effect of taxifolin on TNF-α-induced increase in IL-6
The effect of taxifolin on TNF-α-induced increase in inflammatory cytokine IL-6 was assessed by incubating mature adipocytes on day 16 of differentiation for three hours in the differentiation medium (control) or the medium containing TNF-α and TNF-α + taxifolin. After incubation, the IL-6 concentration in the medium was measured, and the results are shown in Figure 4. TNF-α stimulation significantly increased IL-6 concentration in the medium. Co-treatment with taxifolin (20 µg/mL) significantly inhibited the TNF-α- induced increase in IL-6 level.
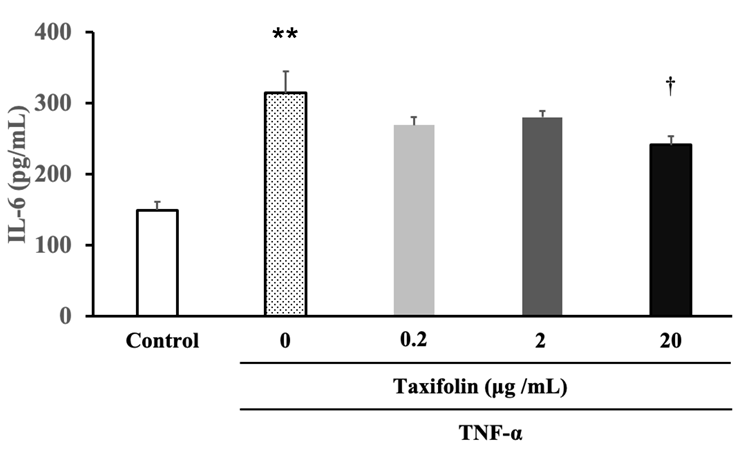
Figure 4: The inhibitory effect of taxifolin on TNF-α-induced increase in inflammatory cytokine IL-6. Mature adipocytes on day 15 of differentiation were incubated for three hours in the differentiation medium (control) or the medium containing TNF-α (final concentrations: 5 ng/mL), TNF-α + various concentrations of taxifolin (final concentrations: 0.2, 2, and 20 µg/mL). After incubation, the IL-6 concentrations in the medium were measured. Each data represents the mean ± SEM (n = 6). **P < 0.01 vs control, †P < 0.05 vs TNF-α: statistical significance was assessed by Dunnett’s test following one-way ANOVA.
Discussion
This study investigated the taxifolin effect on adipocytes using non-cytotoxic concentrations, and demonstrated that taxifolin has anti-differentiation and anti-inflammatory effects in cultured human adipocytes.
Differentiation or maturation of preadipocytes to adipocytes is characterized by the appearance of intracellular lipid droplets and the generation of adiponectin [30]. Therefore, in this study the differentiation process was evaluated by morphological observation and quantification of intracellular lipid droplets stained with Oil Red O and adiponectin generation as the differentiation markers. The number of adipocytes containing lipid droplets and the lipid concentrations increased with increasing days of differentiation. Adiponectin production rapidly increased until reaching mature adipocytes on the 16th day of differentiation but decreased conversely on the 37th day when many swollen adipocytes were observed. This result (Figure 1) is consistent with previous reports showing that adiponectin production is reduced in hypertrophic adipocytes [15, 17]. Therefore, we decided to verify the taxifolin effect on differentiation through the differentiation period for 16 d. Intracellular lipid accumulation was significantly inhibited in adipocytes cultured in a taxifolin- containing differentiation medium than the control medium. Cell morphological observations showed that the number of mature cells in the taxifolin-treated group was lower compared to the control group. Additionally, the adiponectin concentration, an adipocyte-specific cytokine, in the medium on day 16 was significantly lower in the taxifolin group than in the control group. These results induced by treating taxifolin (Figure 2) suggest that taxifolin may suppress preadipocyte- to-adipocyte differentiation.
Adipocyte differentiation and adipogenesis are regulated by transcriptional factors, including CCAAT/ Enhancer-Binding Protein (C/EBP) family members (C/ EBPα, C/EBPβ, and C/EBPδ) and PPARγ [24, 31]. In early adipose differentiation, the expression of transcription factors such as C/EBPβ and C/EBPδ is temporarily increased to induce differentiation. Both transcription factors increase the expression of PPARγ and C/EBPα transcription factors, thereby increasing the expression of adipocyte-specific genes, including fatty acid transport proteins (CD36 and FABP4) and adiponectin, as well as GLUT4. Consequently, intracellular lipid accumulation and adiponectin production are promoted and differentiated to mature adipocytes [24, 31]. Therefore, to further verify the inhibitory effect of taxifolin on adipocyte differentiation, the expression of differentiation-related genes such as PPARγ, C/EBPα, adiponectin, CD36, and GLUT4 mRNAs was measured on day 16 of differentiation. Among the two master regulators of adipocyte differentiation, taxifolin significantly inhibited C/EBPα mRNA expression, but not PPARγ mRNA expression. Taxifolin also significantly inhibited adiponectin and GLUT4 mRNAs and tended to inhibit CD36 mRNA expression (Figure 3). These results further showed that taxifolin suppressed adipocyte differentiation. Although more detailed investigation may be needed, taxifolin may have suppressed the coordination between PPARγ and C/EBPα by significantly reducing C/EBPα mRNA expression. It was speculated that this imbalance between PPARγ and C/EBPα may have suppressed mRNA expression of adiponectin, CD36, and GLUT4, resulting in the suppression of fat accumulation and adiponectin production. The excessive numbers of adipocytes and increased size of adipocytes in adipose tissue contribute to the obesity induction [32]. Theobromine, a cocoa bean-derived methylxanthine, is thought to prevent obesity by suppressing adipocyte differentiation or adipogenesis by suppressing C/EBPβ expression [24]. Therefore, the inhibitory effect of taxifolin on adipocyte differentiation may also be an effective strategy for preventing and treating obesity. The development of chronic inflammatory is closely associated with an increase in infiltration of M1 macrophages into adipose tissue because adipocyte hypertrophy and macrophage infiltration were observed in the adipose tissue of patients with obesity [33, 34]. TNF-α secreted from M1 macrophages that infiltrate adipose tissue activates NF-κB via TNF receptors expressed on the surface of adipocytes. Activated NF-κB translocates into the nucleus, binds to the inflammatory gene promoter region, and ultimately promotes the expression and secretion of proinflammatory cytokines, such as IL-6 and MCP-1 [13, 14, 16]. A thiazolidine derivative, which is a PPARγ agonist, has been reported to suppress the NF-κB pathway via adiponectin produced through the promotion of the PPARγ-adiponectin pathway and exerts an anti-inflammatory effect [13, 14, 35].Therefore, in this study, TNF-α was added to the medium of cultured mature adipocytes on day 16 of differentiation instead of macrophage infiltration to examine taxifolin effect on TNF- α-induced inflammatory cytokines. TNF-α stimulation significantly increased IL-6 concentration in the medium. Taxifolin significantly suppressed the TNF-α-induced increase in IL-6 level, suggesting that taxifolin has anti- inflammatory effects (Figure 4). Taxifolin has been reported to be an Adipo2 receptor agonist [36]. Additionally, pine bark extract containing taxifolin has been reported to inhibit NF-κB activation in macrophages [37] and vascular endothelial cells [38, 39]. These findings suggest that the IL-6 inhibitory effect of taxifolin may be due to a direct inhibition of NF-κB activity and/or an indirect inhibition of NF-κB activity mediated by the Adipo2 receptors, although future verification is necessary.
In conclusion, it was demonstrated here that taxifolin inhibited adipocyte differentiation and exerted anti- inflammatory effects in cultured human adipocytes. This result will be a useful information for elucidating the molecular mechanism of taxifolin for adipocytes in the future. Furthermore, combined with previous results on macrophages [29], taxifolin is expected to have the potential as a therapeutic drug for obesity and metabolic syndrome.
Conclusion
It was first demonstrated in this study that taxifolin had anti-differentiation and anti-inflammatory effects in cultured human adipocytes. Taxifolin is expected to have the potential as a therapeutic drug for obesity and metabolic syndrome.
Acknowledgements
The authors would like to thank Enago (www.enago.jp) for the English language review.
Conflicting interests
This work was supported in part by a research grant from DHQ Co., Ltd (Saitama, Japan).
References
-
Huang PL (2009) A Comprehensive Definition for Metabolic Syndrome. Dis Model Mech 2(5-6): 231-237.
-
Matsuzawa Y (1997) Pathophysiology and Molecular Mechanisms of Visceral Fat Syndrome: The Japanese Experience. Diabetes Metab Rev 13(1): 3-13.
-
Flier JS (2004) Obesity Wars: Molecular Progress Confronts an Expanding Epidemic. Cell 116(2):337-350.
-
Friedman JM (2000) Obesity in the New Millennium. Nature 404(6778): 632-634.
-
Longo M, Zatterale F, Naderi J, Parrillo L, Formisano P, et al. (2019) Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int J Mol Sci 20(9): 2358.
-
Bray GA, Clearfield MB, Fintel DJ, Nelinson DS (2009) Overweight and Obesity: The Pathogenesis of Cardiometabolic Risk. Clin Cornerstone 9(4): 30-40.
-
Chawla A, Chawla R, Jaggi S (2016) Microvasular and Macrovascular Complications in Diabetes Mellitus: Distinct or Continuum? Indian J Endocrinol Metab 20(4): 546-551.
-
Reaven GM (2011) Insulin Resistance: The Link between Obesity and Cardiovascular Disease. Med Clin North Am 95(5): 875-892.
-
Westphal SA (2008) Obesity, Abdominal Obesity, and Insulin Resistance. Clin Cornerstone 9(1): 23-29.
-
Kadowaki T, Yamauchi T, Kubota N, Hara K, Ueki K, et al. (2006) Adiponectin and Adiponectin Receptors in Insulin Resistance, Diabetes, and the Metabolic Syndrome. J Clin Invest 116(7): 1784-1792.
-
Matsuzawa Y (2006) The Metabolic Syndrome and Adipocytokines. FEBS Lett 580(12): 2917-2921.
-
Christiansen T, Richelsen B, Bruun JM (2005) Monocyte Chemoattractant Protein-1 is Produced in Isolated Adipocytes, Associated With Adiposity and Reduced After Weight Loss in Morbid Obese Subjects. Int J Obes (Lond) 29(1): 146-150.
-
Suganami T, Nishida J, Ogawa Y (2005) A Paracrine Loop between Adipocytes and Macrophages Aggravates Inflammatory Changes: Role of Free Fatty Acids and Tumor Necrosis Factor Α. Arterioscler Thromb Vasc Biol 25(10): 2062-2068.
-
Suganami T, Tanimoto Koyama K, Nishida J, Itoh M, Yuan X, et al. (2007) Role of the Toll-Like Receptor 4/NF-Κb Pathway in Saturated Fatty Acid-Induced Inflammatory Changes in the Interaction between Adipocytes and Macrophages. Arterioscler Thromb Vasc Biol 27(1): 84- 91.
-
Tilg H, Moschen AR (2006) Adipocytokines: Mediators Linking Adipose Tissue, Inflammation, and Immunity. Nat Rev Immunol 6(10): 772-783.
-
Zoico E, Garbin U, Olioso D, Mazzali G, Fratta Pasini AM, et al. (2009) The Effects of Adiponectin on Interleukin-6 and MCP-1 Secretion in Lipopolysaccharide-Treated 3T3-L1 Adipocytes: Role of the NF-Κb Pathway. Int J Mol Med 24(6): 847-851.
-
Lau DC, Dhillon B, Yan H, Szmitko PE, Verma S (2005) Adipokines: Molecular Links between Obesity and Atherosclerosis. Am J Physiol Heart Circ Physiol 288(5): 2031-2041.
-
Hammarstedt A, Andersson CX, Rotter Sopasakis V, Smith U (2005) The Effect of PPARγ Ligands on the Adipose Tissue in Insulin Resistance. Prostaglandins Leukot Essent Fatty Acids 73(1): 65-75.
-
Okuno A, Tamemoto H, Tobe K, Ueki K, Mori Y, et al. (1998) Troglitazone Increases the Number of Small Adipocytes without the Change of White Adipose Tissue Mass in Obese Zucker Rats. J Clin Invest 101(6): 1354- 1361.
-
Tao C, Sifuentes A, Holland WL (2014) Regulation of Glucose and Lipid Homeostasis by Adiponectin: Effects on Hepatocytes, Pancreatic Beta Cells and Adipocytes. Best Pract Res Clin Endocrinol Metab 28(1): 43-58.
-
Bhat IA, Kabeer SW, Reza MI, Mir RH, Dar MO (2020) Adiporon: A Novel Insulin Sensitizer in Various Complications and the Underlying Mechanisms: A Review. Curr Mol Pharmacol 13(2): 94-107.
-
Okada Iwabu M, Yamauchi T, Iwabu M, Honma T, Hamagami K, et al. (2013) A Small-Molecule Adipor Agonist for Type 2 Diabetes and Short Life in Obesity. Nature 503(7477): 493-499.
-
Fredholm BB, Lindgren E (1984) The Effect of Alkylxanthines and other Phosphodiesterase Inhibitors on Adenosine-Receptor Mediated Decrease in Lipolysis and Cyclic Amp Accumulation in Rat Fat Cells. Acta Pharmacol Toxicol (Copenh) 54(1): 64-71.
-
Mitani T, Watanabe S, Yoshioka Y, Katayama S, Nakamura S, et al. (2017) Theobromine Suppresses Adipogenesis Through Enhancement of CCAAT-Enhancer-Binding Protein Β Degradation by Adenosine Receptor A1. Biochim Biophys Acta Mol Cell Res 1864(12): 2438- 2448.
-
Yang P, Xu F, Li HF, Wang Y, Li FC, et al. (2016) Detection of 191 Taxifolin Metabolites and Their Distribution in Rats Using HPLC-ESI-IT-TOF-Msn. Molecules 21(9): 1209.
-
Sunil C, Xu B (2019) An Insight into the Health-Promoting Effects of Taxifolin (Dihydroquercetin). Phytochemistry 166: 112066.
-
Ding T, Wang S, Zhang X, Zai W, Fan J, et al. (2018) Kidney Protection Effects of Dihydroquercetin on Diabetic Nephropathy Through Suppressing ROS and NLRP3 Inflammasome. Phytomedicine 41: 45-53.
-
Sun X, Chen RC, Yang ZH, Sun GB, Wang M, et al. (2014) Taxifolin Prevents Diabetic Cardiomyopathy in Vivo and In Vitro by Inhibition of Oxidative Stress and Cell Apoptosis. Food Chem Toxicol 63: 221-232.
-
Muramatsu D, Uchiyama H, Kida H, Iwai A (2020) In Vitro Anti-Inflammatory and Anti-Lipid Accumulation Properties of Taxifolin-Rich Extract from The Japanese Larch, Larix Kaempferi. Heliyon 6(12): e05505.
-
Fu Y, Luo N, Klein RL, Garvey WT (2005) Adiponectin Promotes Adipocyte Differentiation, Insulin Sensitivity, and Lipid Accumulation. J Lipid Res 46(7): 1369-1379.
-
Lefterova MI, Zhang Y, Steger DJ, Schupp M, Schug J, et al. (2008) Pparγ And C/EBP Factors Orchestrate Adipocyte Biology Via Adjacent Binding on a Genome-Wide Scale. Genes Dev 22(21): 2941-2952.
-
Gesta S, Tseng YH, Kahn CR (2007) Developmental Origin of Fat: Tracking Obesity to Its Source. Cell 131(2): 242-256.
-
Xu H, Barnes GT, Yang Q, Tan G, Yang D, et al. (2003) Chronic Inflammation in Fat Plays A Crucial Role in the Development of Obesity-Related Insulin Resistance. J Clin Invest 112(12): 1821-1830.
-
Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, et al. (2003) Obesity is Associated with Macrophage Accumulation in Adipose Tissue. J Clin Invest 112(12): 1796-1808.
-
Achari AE, Jain SK (2017) Adiponectin, A Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int J Mol Sci 18(6): 1321.
-
Sun Y, Zang Z, Zhong L, Wu M, Su Q, et al. (2013) Identification of Adiponectin Receptor Agonist Utilizing A Fluorescence Polarization Based High Throughput Assay. PLOS ONE 8(5): e63354.
-
Cho KJ, Yun CH, Yoon DY, Cho YS, Rimbach G, et al. (2000) Effect of Bioflavonoids Extracted from the Bark of Pinus Maritima on Proinflammatory Cytokine Interleukin-1 Production in Lipopolysaccharide Stimulated RAW 264.7. Toxicol Appl Pharmacol 168(1): 64-71.
-
Peng Q, Wei Z, Lau BH (2000) Pycnogenol Inhibits Tumor Necrosis Factor-Α-Induced Nuclear Factor Κb Activation and Adhesion Molecule Expression in Human Vascular Endothelial Cells. Cell Mol Life Sci 57(5): 834-841.
-
Ohkita M, Kiso Y, Matsumura Y (2011) Pharmacology in Health Foods: Improvement of Vascular Endothelial Function by French Maritime Pine Bark Extract (Flavangenol). J Pharmacol Sci 115(4): 461-465.
- Gallic and Citric Acid Present in the Peels of Tropical Fruits as an Alternative in the Fight against Cancer
- Treating the Forehead Lines with Combination of Forehead and Glabellar Botulinum Toxin Among Japanese Patients
- Clinical Evaluation of Patients Suffering from Breast Cancer & Determination of Treatment Therapies and Better Strategies Related to Breast Cancer
- Medieval Recipes by Al-Zahrāwī for Heart Palpitations Treatment
- Etiology and Prescription Errors of Myocardial Infarction in Different Health Care Systems of Azad Kashmir
- Early Diagnosis and Multidisciplinary Management of Turner Syndrome: A Paediatric Case Study