Effect of Diterpenoid Alkaloid Thalatisamine (Aconitum Talassicum) and its Derivatives on Rat Liver and Heart Mito K+ Atp- Channels
Recently, the properties of mitochondrial ATP-dependent potassium (mitoK+ATP-channel) channels isolated from variousorgans have been deeply studied. Activation of the MitoK+ATP-channel leads to an increase in the size of mitochondria,acceleration of ATP synthesis, stimulation of respiration. In order to find new, effective modulators of mitoK+ATP-channel, theeffect of diterpenoid alkaloids thalatisamine and its derivative 14-O-benzoyltalatisamine isolated from Aconitum talassicumplant on mitoK+ATP-channel in rat liver and heart was studied. According to the results of the study, concentrations of 30 and50 μM of talatisamine, compared to the control, decreased the mitoK+ATP-channel of the liver by 72.1±2.1% and 107.7±2.0%,respectively, and the mitoK+ATP-channel of the heart by 37.1 ±2.4% and 88.5±2.9% reliable activation was found. Under theseconditions, concentrations of 30 and 50 μM of 14-O-benzoyltalatisamine decreased liver mitoK+ATP-channel by 145±2.3% and196±2.5%, respectively, and heart mitoK+ATP-channel by 116±3, 1% and activated by 149.7±2.5% reliable activation. Accordingto the obtained results, the effect of 14-O-benzoyltalatisamine on liver and heart mitoK+ATP-channel was found to be more activecompared to talatizamine alkaloid. Therefore, this alkaloid and its derivative require a wide range of studies as a cytoprotectorand cardioprotective agent protecting against ischemia by activating the mitoK+ATP-channel of the liver and heart.
Muratova DK¹*, Ergashev NA² and Asrarov MI²
¹Department of Biophysics, National University of Uzbekistan, Uzbekistan ²Institute of Biophysics and Biochemistry, National University of Uzbekistan, Uzbekistan
Introduction
Mitochondria is an organelle that synthesizes energy in living organisms, including mammalian cells, and is important in physiological and pathological conditions. Mitochondria are the “energy generators” of cells, transporting electrons generated in the Krebs cycle through the respiratory chain and producing ATP as a result of oxidative phosphorylation [1]. In addition to synthesizing ATP, mitochondria play an important role in various areas of cell physiology. One of the main properties of mitochondria is the presence of transmembrane potential generation in its inner membrane [1]. It is known from literature and experimental results that functional structures located in mitochondria and membranes are the main “target structure” for various pathological effects [2, 3, 4, 5]. Mitochondrial dysfunction leads to various pathologies ranging from minor changes in cell function to cell death [6]. MitoK+ATP-channels is a selective channel that plays an important role in the life of eukaryotic organisms, including cells. Recently, the properties of mitoK+ATP channels isolated from various organs have been deeply studied. It is known from many literatures that this channel is involved not only in physiological conditions, but also in pathological conditions such as ischemia, hypoxia, necrosis and apoptosis, in controlling the size of mitochondria, as well as in maintaining its integrity [7, 8].
Most modulators acting on the MitoK+ATP-channel are synthetic reagents that can either activate or inhibit the channel. At the same time, they can cause disruption of the regulation of cellular ion transport. Biologically active substances control the size of mitochondria by activating the mitoK+ATP-channel. At the same time, it participates in the acceleration of ATP synthesis, stimulation of respiration, protection of the myocardium in the case of ischemia, and prevention of apoptosis in oxidative stress [9].
In order to create effective drugs for the prevention and treatment of liver, cardiovascular, neurodegenerative and other diseases based on plant bioactive compounds, the mechanisms of action of diterpenoid alkaloids are also being thoroughly studied [10, 11, 12, 13]. Since plant bioactive compounds do not negatively affect the functional structures of mitochondria, finding new, effective modulators of mitoK+ATP-channel is one of the important tasks of biophysics and pharmacology. Based on the above, the effects of talatisamine [14] and its derivative 14-O-benzoyltalatisamine diterpenoid alkaloids isolated from Aconitum talassicum plant on the mitoK+ATP-channel of rat liver and heart were studied in vitro.
Materials and Methods
**Extraction and Separation of Alkaloids**
The aerial part of the plant was used as a source. Total alkaloids (3.55 g) of KSK was separated on a column of silica gel (sorbent: compound ratio 50:1) eluting with CHCl3; MeOH. Treatment with acetone and recrystallization from MeOH gave talatisamine (0.81 g) of the fractions obtained using CHCl3 and CHCl3; MeOH (100:1). It was determined by comparing TLC (Fluka Analytical, Germany), the mixt comparison of melting point and IR (Perkin-Elmer-2000) and PMR spectra with real sample. Elution by CHCl3; MeOH (50:1 and 25:1) isolated isotalatisine (0.17 g). Fractions eluted with CHCl3; MeOH (1:1) contained talatisine (0.08 g). The mother liquor of these fractions was subjected to rechromatography on a silica gel column (sorbent: compound ratio 20:1) and isotalatisine (0.08 g) and talatisine (0.03 g) were obtained. The resulting mother liquors after elution by CHCl3, it was rechromatographed on a silica gel column (sorbent: compound ratio 20:1) using CHCl3 and CHCl3; MeOH as eluents. Elution by CHCl3 is isolated 14-O-acetyltalatisamine (0.04 g). Further fractions eluted by CHCl3; MeOH (100:1, 50:1) afforded talatisamine (0.27 g). It contained talatisamine 0.18% of dry plant weight. Talatisamine (0.12% of dry plant weight) and isotalatisine and talatisine analogs were isolated method of separation from the total alkaloids of the aerial part of the plant collected during the flowering period. Diterpene alkaloid talatisamine, which was isolated from Aconitum talassicum (C24H39NO5) plants growing in the territory of the Republic of Uzbekistan. Figure 1 shows the structural formulas of talatisamine and 14-O-benzoyltalatisamine. The structural formulas of alkaloids were drawn by the ChemOffice 2002, Chem Draw Ultra 7.0 software (Figure 1).
Mitochondria Isolation
Mitochondria were isolated from the rat liver by differential centrifugation accordind to Schneider [15]. Briefly, rat liver was homogenized in a medium (in mM):
$$\text{sucrose} - 250, \text{Tris-HCl} - 10, \text{EDTA} - 1, \text{pH} 7.4 \text{ centrifuged at } 1500 \times \text{g for } 7 \text{ min} (0-2^{\circ}\text{C}).$$ Mitochondria were sedimented by centrifugation of supernatant at 6000×g for 15 min (0-2^{\circ}\text{C}). The final mitochondrial pellet was suspended in a small volume of medium containing sucrose - 250 mM, Tris-HCl - 10 mM, was kept on ice prior to experiments. Rat heart mitochondria were also isolated using differential centrifugation. Rat heart was homogenized in a medium (in mM): sucrose - 300, Tris-HCl - 10, EDTA - 2, albumin 0.2%, pH 7.4. To isolate a sufficient amount of mitochondria, 5 or 6 rat hearts were isolated and placed in chilled isolation medium. Laboratory equipment and petri dishes were stored in a freezer, and the procedure was carried out on ice. Rat hearts were cleaned of adipose tissue, blood and other large blood vessels in chilled 0.9% KCl solution. The cleaned heart was washed again, dried using filter paper, heart mass was determined and cut into small pieces using scissors. After that, the minced heart tissue was placed in a homogenizer and homogenized using a teflon pestle, poured with a 10:1 ratio of separation medium.
Because the heart tissue is composed of transverse muscles, it was more homogenized than the liver tissue. The homogenate was poured into a centrifuge tube, placed in the rotor and centrifuged at 1500 g for 7 min (0-2°C). At this stage, heavy aggregates settle. In the next step, centrifugation was performed at 6000 g for 20 min (0-2°C). Isolated mitochondria were washed in medium without EDTA and albumin. Mitochondria isolation procedures were carried out under cold conditions. Mitochondria were kept on ice during the experiments. The mitochondrial protein content was determined by the Lowry method by Peterson [16].
A Method for the Determination of ATP- Dependent K+-Channel Conductance in Mitochondria
ATP-dependent K+-channel conductance in mitochondria was determined by optical density at a wavelength of 540 nm. Incubation medium (IM, mM): KCl-25, HEPES-10, succinate-5, MgCl2 -1, K2HPO4 -2.5, KH2PO4 -2.5, rotenone -0.005 and oligomycin -0.001, pH-7.4 [17]. The amount of protein in the medium was 0.3-0.4 mg/ml.
Drugs and Chemicals
The following chemical reagents were used: EDTA, KCl, succinate, MgCl2, KH2PO4, K2HPO4, sucrose, (“Chemreaktivsnab”, Russia), Tris-HCl (“Serva”, Germany), HEPES, diazoxide, olygomycin, rotenone, ATP (“Sigma”, USA). Tris-HCl, albumin. All reagents are chemically pure.
Data Analysis
The results were analysed statistically using the Origin Pro 7.5 (Microsoft, USA). The data were evaluated using parametric Student’s t-test and are expressed as M ± m. The results that were deemed significant are indicated as follows: * p<0.05, p<0.01, and *-P<0.001.
Results
In these studies, the permeability of the mitoK+ATP channel in the absence of ATP in the incubation medium (IM) was taken as 100% (Figure 2). Addition of 200 μM ATP IM inhibited the liver mitoK+ATP-channel. The effect of 1-5 μM concentrations of talatisamine alkaloid on the liver mitoK+ATP-channel in the presence of ATP in the IM was hardly noticeable. During the experiment, when the liver mitoK+ATP-channel was exposed to a concentration of 10 μM of Talatisamine alkaloid, the liver mitoK+ATP-channel was activated by 27.6±1.7% compared to the condition inhibited by ATP (control). Talatisamine concentrations of 30 and 50 µM were found to reliably activate the liver mitoK+ATP- channel by 72.1±2.1% and 107.7±2.0%, respectively, compared to the control. Under these conditions, when compared with diazoxide, a classic activator of the channel, talatisamine had an activating effect on the liver mitoK+ATP- channel like diazoxide (Figure 2A).
It is known that in the presence of K+ ions in the environment, mitochondrial contraction is inhibited by ATP, the inhibitory effect of ATP is reactivated by mitoK+ATP- channel activators such as diazoxide, kromakalim, pinacidil [18]. As a result of activation of the MitoK+ATP-channel, the increase in matrix volume leads to stimulation of respiration and an increase in ATP synthesis [19, 20].
Therefore, talatisamine and other activators of the mitoK+ATP-channel led to an increase in the matrix volume, these properties of talatisamine play an important role in the creation of new antihypoxant or hepatoprotective drugs. Several scientific studies show that mitoK+ATP- channel in heart tissue plays a leading role in protecting the cell from ischemia-reperfusion [5, 7, 9, 21, 22]. Activation of K+ channels in the tissue before or during ischemia is directly associated with cytoprotection [23]. Since the activation of MitoK+ATP-channel protects the cell from ischemia and reperfusion state, the effect of talatisamine on heart mitoK+ATP-channel was studied in our next studies. In IM without ATP, heart mitoK+ATP-channel block was taken as a control (100%).
To determine the effect of talatisamine on the heart mitoK+ATP-channel, in the presence of 200 μM ATP in the IM, alkaloid talatisamine did not affect the channel activity at a concentration of 1 μM. In the experiment, it was found that 10 µM concentration of talatisamine activates heart mitoK+ATP-channel activity by 14.2±3.1% compared to the control. Talatisamine concentrations of 30 and 50 μM were found to reliably activate heart mitoK+ATP-channel activity by 37.1±2.4% and 88.5±2.9%, respectively, compared to the control. Thus, talatisamine alkaloid acts on heart mitoK+ATP- channel as an activator (Figure 2B).
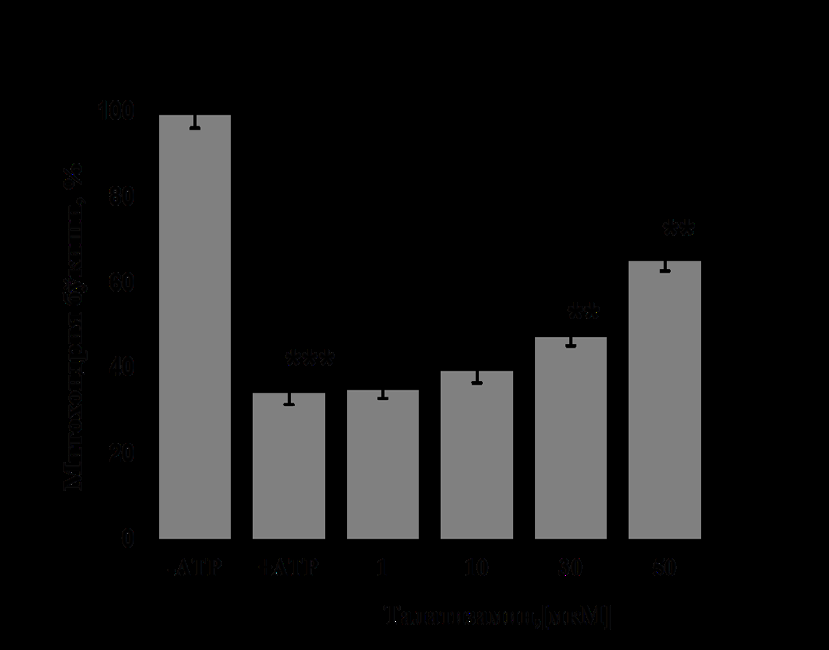
(On the ordinate axis, mitochondria staining is shown, and on the abscissa axis, different concentrations of the substance are shown. The reliability of statistical indicators under the influence of diazoxide (Dzx 10 μM) and talatisamine was obtained in comparison with the condition when 200 μM ATP was inhibited -*-P<0.05; -P<0.01; *-P<0.001; n=6). In subsequent experiments, the effect of 14-O-benzoyltalatisamine, a derivative of talatisamine alkaloid, on the liver mitoK+ATP-channel was determined. 14-O-benzoyltalatisamine was obtained by introducing a benzoyl group as a result of the acylation reaction of the hydroxyl group at the C-14 position of the alkaloid talatizamine carbon atom in the presence of benzoyl chloride.
In the presence of ATP in the IM, it was found that 1μM concentration of 14-O-benzoyltalatisamine activated it by 45±1.7% compared to the control (Figure 3A). A concentration of 10 μM of 14-O-benzoyltalatisamine was found to activate liver mitoK+ATP-channel activity by 76±2.1% compared to the control. Concentrations of 30 and 50 μM of 14-O-benzoyltalatisamine reliably activated liver mitoK+ATP-channel activity by 145±2.3% and 196±2.5%, respectively, compared to control. So, 14-O-benzoyltalatisamine alkaloid acts like an activator on the liver mitoK+ATP-channel (Figure 3A). In experiments on the effect of 14-O-benzoyltalatisamine alkaloid on heart mitoK+ ATP-channel, in the presence of 200 μM ATP in the IM, it was found that the activity of the channel was inhibited by 65±3.1% compared to the control (Figure 3B). 1μM concentration of 14-O-benzoyltalatisamine activated by 21.4±2.1%, 10 μM concentration by 48±2.8% in the inhibited state of heart mitoK+ ATP-channel. Concentrations of 30 and 50 μM of this alkaloid were found to reliably activate heart mitoK+ATP-channel activity by 116±3.1% and 149.7±2.5%, respectively, compared to the control (Figure 3B).
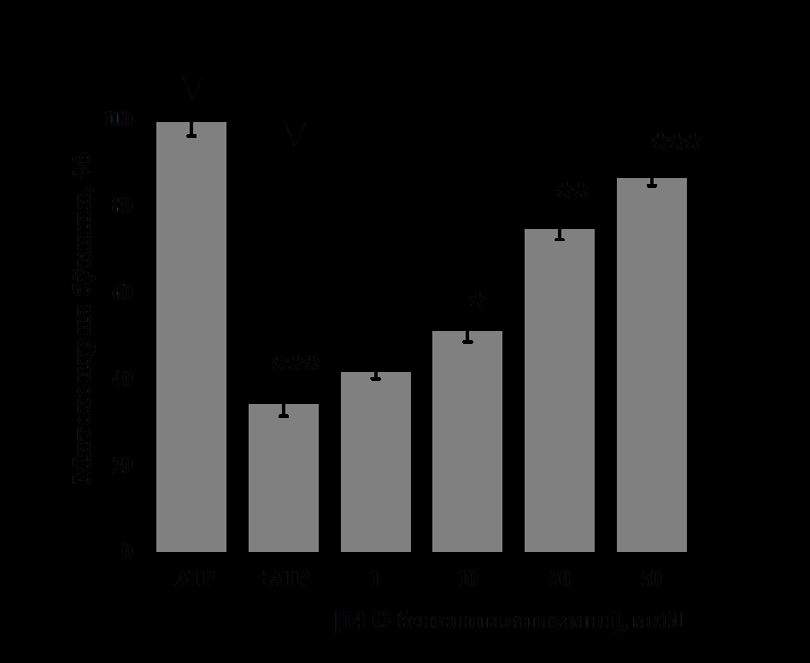
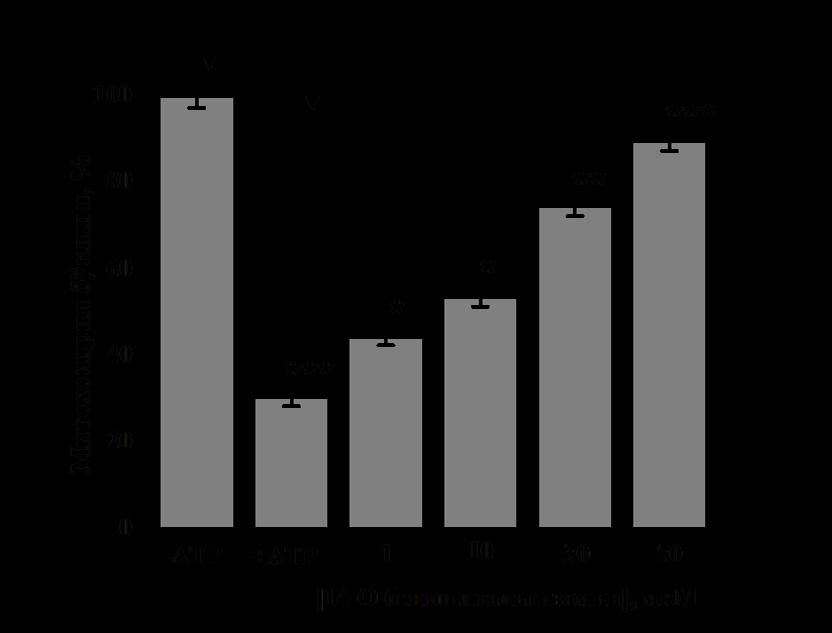
(Reliability of statistics on the effect of 14-O-benzoyltalatisamine was obtained relative to the condition when 200 μM ATP was inhibited- *-Р<0,05; - Р<0,01; *-Р<0,001; n=5). Thus, it was found that the effect of 14-O-benzoyltalatisamine on liver and heart mitoK+ATP-channel is much more active than that of alkaloid talatizamine. The introduction of the benzoyl group may have increased the activity and lipophilicity of the substance, as a result, the effect of 14-O-benzoyltalatisamine on the mitoK+ATP-channel showed a higher activity than that of the alkaloid talatisamine. Previous experiments have shown that the addition of a benzoyl group to the alkaloid talatisamine increases its antioxidant activity [13].
Conclusion
MitoK+ATP-channel is important in normal and pathological processes as it regulates the size and function of mitochondria. A. According to Szewczyk and L. Wojtczak, the effect of pharmacological drugs on mitochondria is divided into: drugs that modulate apoptosis, drugs that have protective effects that activate energetic processes in the cell (immunosuppressants, antiviral drugs, nonsteroidal anti-inflammatory drugs, anesthetics) [24]. Activation or inhibition of mitochondrial potassium channels by specific drugs is used to induce cytoprotection or control cell death [25]. Many studies have been conducted in cardiomyocyte mitoK+ATP-channel to study the properties of substances that activate them [26]. Such MitoK+ATP-channel activating agents have been found to depolarize heart mitochondria [27]. Therefore, the derivatives of diterpenoid alkaloids require a wide range of studies as cytoprotectors and cardioprotective agents protecting against ischemia by activating liver and heart mitoK+ ATP-channel.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
-
Logan DC (2006) The mitochondrial compartment. J Exp Bot 57(6): 1225-1243.
-
Szewczyk A, Wojtczak L (2002) Mitochondria as a pharmacological target. Pharmacol Rev 54(1): 101-127.
-
Szeto HH (2008) Mitochondria-targeted cytoprotective peptides for ischemia-reperfusioninjury. Antioxid Redox Signal 10(3): 601-619.
-
Smith RAJ, Hartley RC, Cochemé HM, Murphy MP (2012) Mitochondrial pharmacology. Trends in Pharmacological Sciences 33(6): 341-352.
-
Di LF, Canton M, Menabo R, Kaludercic N, Bernardi P (2007) Mitochondria and cardioprotection. Heart Fail Rev 12(3-4): 249-260.
-
Jacobson J, Duchen MR (2004) Interplay between mitochondria and cellular calcium signaling. Molecular and Cellular Biochemistry 256 (1-2): 209-218.
-
Akao M, Ohler A, O’Rourke B, Marbán E (2001) Mitochondrial ATP-sensitive potassium channels inhibit apoptosis induced by oxidative stress in cardiac cells. Circ Res 88(12): 1267-1275.
-
Costa ADT, Quinlan C, Andrukhiv A, West IC, Garlid KD (2006) The direct physiological effects of mitoKATP opening on heart mitochondria. Am J Physiol 290(1): 406-415.
-
Hausenloy DJ, Schulz R, Girao H, Kwak BR, Stefani DD, et al. (2020) Mitochondrial ion channels as targets for cardioprotection. J Cell Mol Med 24(13): 7102-7114.
-
Muratova DK, Ergashev NA, Shkinev AV, Asrarov MI, Kurbanov UH (2021) Effect of songorine on the activity of atp-dependent k+ channels and the state of megapore in rat liver mitochondria. Experimental and Clinical Pharmacology 84(4): 12-15.
-
Muratova DK, Ergashev NA, Sobirov JJ, Kurbanov UKH, Asrarov MI (2021) Effects of diterpene alkaloids on lipid peroxidation in mitochondria. Nova Biotechnol Chim 20(2): 1-8.
-
Ernazarov ZM, Pozilov MK, Asrarov MI, Zhurakulov SN (2023) Effects of dihydroquercetin, 1-aryltetrahydroisoquinoline, and conjugate on the functional condition mitochondrial membrane of the rat liver. Nova Biotechnologica et Chimica 22(1): 1-8.
-
Muratova DK, Ergashev NA, Asrarov MI (2023) Effect of Talatisamine and its Derivate 14-O-benzoyltalatisamine on Functional State of Rat Liver and Heart Mitochondria. Biomedical & Pharmacology Journal 16(4): 2333-2343.
-
Eshmatov ZM, Sultankhodzhaev MN, Nigmatullaev AM (2011) Accumulation dynamics of alkaloids in Aconitum talassicum. Chem Nat Comp 47: 149-150.
-
Schneider WC, Hogeboom GH (1951) Cytochemical studies of mammalion tissues: the isolation of cell components by differential centrifugation. Cancer Res 11(1): 1-22.
-
Peterson GL (1977) A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal Biochem 83(2): 346-356.
-
Vadzyuk OB, Kosterin SA (2008) Diazoxide-induced swelling of rat myometrial mitochondria as evidence of activation of the ATP-sensitive K+ channel. Ukr biochem Journal 80(5): 45-51.
-
Costa ADT, Quinlan C, Andrukhiv A, West IC, Garlid KD (2006) The direct physiological effects of mitoKATP opening on heart mitochondria. Am J Physiol 290(1): 406-415.
-
Korge P, Honda HM, Weiss JN (2005) K+-dependent regulation of matrix volume improves mitochondrial function under conditions mimicking ischemia- reperfusion. Am J Physiol Heart Circ Physiol 289(1): H66-H77.
-
Kaasik A, Safiulina D, Zharkovsky A, Veksler V (2007) Regulation of mitochondrial matrix volume. Am J Physiol Cell Physiol 292(1): C157-C163.
-
Garlid KD, Paucek P, Yarov‐Yarovoy V, Murray HN, Darbenzio RB, et al. (1997) Cardioprotective effect of diazoxide and its interaction with mitochondrial ATP‐sensitive K+ channels. Possible mechanism of cardioprotection. Circ Res 81(6): 1072‐1082.
-
Paggio A, Checchetto V, Campo A, Menabò R, Marco GD, et al. (2019) Identification of an ATP‐sensitive potassium channel in mitochondria. Nature 572(7771): 609‐613.
-
Nakai Y, Horimoto H, Mieno S, Sasaki S (2001) Mitochondrial ATP-sensitive potassium channel plays a dominant role in ischemic preconditioning of rabbit heart. Eur Surg Res 33(2): 57-63.
-
Szewczyk А, Wojtczak L (2002) Mitochondria as a Pharmacological Target. Pharmacol Rev 54(1): 101-127.
-
Wrzosek A, Augustynek B, Żochowska M, Szewczyk A (2020) Mitochondrial Potassium Channels as Druggable Targets. Вiomolecules 10(8): 1200.
-
Holmuhamedov EL, Jahangir A, Oberlin A, Komarov A, Colombini M, et al. (2004) Potassium channel openers are uncoupling protonophores: implication in cardioprotection. Febs Letters 568(1-3): 167-170.
-
Zhu HF, Dong JW, Zhu W, Ding HL, Zhou ZN (2003) ATP- dependent potassium channels involved in cardiac protection induced by intermittent hypoxia against ischemia-reperfusion injury. Life Sci 73(10): 1275-1287.
- Gallic and Citric Acid Present in the Peels of Tropical Fruits as an Alternative in the Fight against Cancer
- Treating the Forehead Lines with Combination of Forehead and Glabellar Botulinum Toxin Among Japanese Patients
- Clinical Evaluation of Patients Suffering from Breast Cancer & Determination of Treatment Therapies and Better Strategies Related to Breast Cancer
- Medieval Recipes by Al-Zahrāwī for Heart Palpitations Treatment
- Etiology and Prescription Errors of Myocardial Infarction in Different Health Care Systems of Azad Kashmir
- Early Diagnosis and Multidisciplinary Management of Turner Syndrome: A Paediatric Case Study