Assessing the Bioavailability and Bioaccessibility of Selenium from Selenium-Rich Algae
Many new selenium (Se) sources for supplementation use are emerging. The objective of this study was to test the Se bioaccessibility (BAC) and bioavailability (BA) of Se-rich algae. Algal samples were cultivated in media containing selenite or selenate then BAC and BA were compared to Se-salts and Se-amino acids from traditional supplementation. BAC was tested based on the Se solubility post in vitro digestion and BA by induction of cellular glutathione peroxidase activity (GPx) in a Se deficient Caco-2 cell model. Cells treated with algae grown in selenate and selenite had a mean GPx activity that was significantly less than the SeMet treatment (P
Introduction
Selenium (Se) is an essential trace mineral to human and animal health. It is known for its antioxidant and inflammatory properties, maintaining thyroid hormone metabolism, and even has protective properties against certain forms of cancer [1, 2, 3]. One of its most studied roles is as a cofactor of cellular glutathione peroxidase (GPx), one of the antioxidant enzymes. As all selenoproteins, GPx requires a selenocysteine (SeCys) at its catalytic site and is essential for removal of reactive oxygen species generated from aerobic metabolic reactions [4, 5]. Specifically, GPx catalyzes the reaction of a hydrogen peroxide and two reduced forms of glutathione molecules, resulting in an oxidized glutathione and water [6, 7]. The dose and which chemical form Se is consumed though has an influence on the level of GPx activity [6].
When assessing Se metabolism, it is important to distinguish bioaccessibility (BAC) from bioavailability (BA), and how the body metabolizes different chemical forms of this element. Absorbable fraction such as solubility of an element during gastrointestinal digestion is referred to as BAC. Once absorbed, an element can be distributed to cells for functional use, characterizing its BA [8, 9]. Selenomethionine (SeMet), an organic form, is often considered to have the greatest BAC and is less toxic compared to inorganic Se [10]. Inorganic selenite is typically shown to have lower BAC than the organic forms and cannot be stored in the body but inorganic selenate tends to have a BAC comparable to organic Se8. However, selenite has been shown to induce a higher cellular GPx activity compared to organic Se because it is immediately reduced to an available form for selenoprotein synthesis [11, 12]. Other studies suggest that organic selenized yeast has higher BAC than inorganic Se but also results in a lower GPx activity [13]. Overall, once Se is absorbed, it follows three main routes of metabolism: selenoprotein synthesis, incorporation into body proteins, or excretion [1, 14]. The mechanism of Se absorption is dependent upon the chemical form consumed. SeMet is absorbed to a greater extent throughout the small intestine, particularly in the jejunum using active transporters along with its sulfur analogues [15, 16]. Selenate is absorbed via Na+-dependent active transporters and increases toward the distal end of the small intestine [17]. Selenite on the other hand, utilizes passive diffusion and increases toward the distal end of the small intestine [18, 19]. Following intestinal epithelial uptake, inorganic Se is selectively taken up by the red blood cells and immediately reduced to hydrogen selenide (H2Se) and transported by albumin to the liver for selenoprotein synthesis or Se excretion [12, 20]. SeMet follows a longer trans-selenation pathway to generate H2Se, which is similar to the trans-sulfuration pathway taken by methionine [7, 14]. H2Se is then converted back to SeCys for co- translational incorporation into various selenoproteins, or methylated for excretion [1, 7, 14].
In supplementation, inorganic Se-salts and Se-rich yeasts are commonly used [21, 22]. Algae though are being recognized more for its economic advantage and its ability to hold high concentrations of Se [23]. Incorporating algae in the form of Chlorella sp. into dietary supplements is popular because it is a natural product containing a variety of vitamins (e.g. vitamin C and E), minerals (e.g. iron and calcium), and fiber [24, 25]. Over 50% of Chlorella sp. dry mass is as protein, providing all of the essential amino acids. Approximately 26% of the dry mass is lipids, providing polyunsaturated and monounsaturated fatty acids such as oleic acid and linoleic acid [26, 27].
Gross-Wen Technologies, LLC (GWT) developed a Revolving Algal Biofilm System (RAB) that produces quantities of algal biomass that are 10 times greater compared to other existing systems such as open-pond systems. The RAB system is cost-effective and has the ability to remove nutrients like Se from the effluent generated in food manufacturing [28]. However, no studies are reported to date the use of that algal biomass for nutritional studies, especially the BA of Se. Current methods for Se BA include animal and cell models. Solubility after in vitro digestion can be used as a surrogate for BAC since it is a prerequisite for any mineral to be absorbed. GPx activity is often used as a surrogate to Se BA in a cell model because it gives an accurate representation of the intestinal uptake and utilization of different forms of Se [29, 30]. The objectives of this study were to assess the BAC and BA of Se from the algae grown in different conditions with varying concentrations of selenite and selenate, and compare them to organic and inorganic Se often used in supplements.
Materials and Methods
Materials
L-Selenomethionine (SeMet), pancreatin and pepsin were purchased from Millipore Sigma (MA, USA); Sodium Selenite (Na2SeO3) from Alfa Aesar (MA, USA), and Sodium Selenate (Na2SeO4) were obtained from Fisher Scientific (MA, USA). For the cell culture, a human adenoma carcinoma cell line (Caco-2) was obtained from American Type Culture Collection (USA). For the media, fetal bovine serum (FBS) was purchased from Atlanta Biological (GA, USA); Advanced Dulbecco’s Modified Eagle Medium (DMEM), Gibco HEPES 1M, Gibco Antibiotic-Antimycotic 100x solution, and Gibco L-Glutamine 200 mM from Life Technologies, Thermo Fisher Scientific (CA, USA). Glutathione Peroxidase Assay Kits were purchased from Cayman Chemicals, (MI, USA). Phosphate Buffered Saline (PBS) and 0.5% Trypsin EDTA was purchased from Life Technologies, Thermo Fisher Scientific (CA, USA).
Preparation of Algal Samples
Se-rich algae production: Se removal from wastewater is of importance due to its toxicity to plant microbiota even at a concentration of several micrograms per liter [31]. The GWT established this Se-rich algae production research to test the feasibility of 1) using the RAB system to uptake Se from wastewater for Se-algae production, and 2) using the Se-algae as a Se supplement for humans. Throughout the project, chemically defined medium, i.e., Bold’s Basal Medium (BBM) with inorganic Se chemicals (Na2SeO3 and Na2SeO4) was blended in to mimic Se-containing wastewater. Polyculture form a consortium of local algal species in central Iowa was used as the algal seed. Selenium concentrations were tested in a range of 10-500 mg/L to compare the two types of algae culture systems, suspension- based bubble column reactors and RAB reactors. Besides
the Se chemical species, algae culture systems, and initial Se concentration in medium, some other factors including culture time and initial sulfur concentrations (controlled by adding sulfite) in medium were evaluated in affecting the Se BAC and BA in the dry algal biomass.
Se-rich algae culture in suspended system: The suspended algae cultivation was performed in glass bubble columns (Figure 1A). Each bubble column had a working volume of 1.2 L and was aerated with atmosphere air at 1.2 L/min. The initial cell culture had an optical density (OD) of 0.7 at 680 nm. The bubble columns were placed in an indoor laboratory with ambient temperature of 25°C and illuminated with fluorescent light bulbs at 110-120 μmol s-1 m-2. The weekly biomass harvesting was established through centrifugation at 1,860 X g for 4 minutes and freeze dried for further analyses.
Se-rich algae culture in RAB reactor: The design and operation of the RAB systems were described previously [32, 33] (Figure 1B). The RAB reactor included a liquid reservoir (1.2 L working volume) and a vertically oriented rotating belt with a surface area of 0.12 m2. During operation, the belt rotated at a speed of 1.2 rpm. The RAB reactor was placed in an indoor laboratory with ambient temperature of 25°C and illuminated with fluorescent light bulbs at 110-120 μmol s-1 m-2. The algal growth was initiated by inoculating the seed culture into the RAB reservoir. The RAB system was then operated for three weeks to establish a stable algae biofilm on the belt. During this inoculation period, the reservoir was supplemented with DI-water as necessary to compensate for water evaporative loss. The biomass harvesting was done on a weekly basis. After three weeks inoculation with BBM, Polyculture in RAB reactors started a 5-week continuous culture test with a hydraulic retention time (HRT) of 6 days, which was established by replacing 0.2 L of cell broth with fresh Se-containing BBM. The weekly Se-rich biomass harvesting was done by scraping the biofilm off the belt prior to freeze drying.

(A) (B) Figure 1: Production Se-rich algae using suspension-based glass bubble columns (A) and RAB reactor (B).
Algal biomass sample analysis: The dry algae biomass was digested with 5 ml concentrated nitric acid in a Multiwave Go microwave system (Anton Paar, Austria). The digested solution was diluted 10 times with deionized water and then measured for its Se concentration using inductively coupled plasma (ICP) optical emission spectrometer (Thermo Scientific, iCAP 7000 Series, USA). The analyzing program was controlled by Qtegra (Version 2.7.2425.65, Thermo Scientific). Nitric acid (2%) was used as rinse solution and Yttrium ICP standard (5 ppm) was used as the internal standard. The Se content in the dry algal biomass was determined based on the Se concentration in the solution and the weight of dry biomass used, resulting in a final Se percentage of dry mass.
In Vitro Simulated Human Digestion
SeMet, Na2SeO3, Na2SeO4, and Se-enriched algal biomass were subjected to simulated in vitro human digestion for assessing soluble Se, as well as Se cellular uptake for assessing BAC and BA, respectively. Each sample with 200 µg of Se was mixed with Mili-Q water to form slurry. The slurry was initially adjusted to pH 2 and incubated along with pepsin at 37°C for 1 hour while shaking at 300 rpm to simulate gastric digestion. After adjusting to pH 6 and adding pancreatin, samples were incubated again at 37°C for 1 hour while shaking at 300 rpm to simulate duodenal digestion. Enzyme deactivation was achieved by boiling the samples for 4 minutes immediately following second incubation as suggested for iron [34]. Finally, samples were centrifuged at 1070 x g for 15 minutes at 37°C before collecting supernatant and storage at-20°C. Se content was measured in duplicates using ICP-OES system as described to assess percent solubility and intestinal cellular uptake of Se. Although 46 algal samples in total went through in vitro digestion and solubility analysis, only the samples that had approximately 10% solubility were used for cellular uptake. Table 1 lists the algal samples that were used for the study, including a description of the names and designated label. The algae samples that were not used due to low Se values were not shown in the table.
| Selenite (ppm)a | ||||
|---|---|---|---|---|
| Sulfite (ppm) | Wkb | Se (dry mass %) | Label | |
| 30 | 0 | 1 | 0.51 | A1 |
| 30 | 0 | 3 | 1 | A2 |
| 30 | 0 | 2 | 0.56 | A3 |
| 200 | 200 | 3 | 0.16 | A4 |
| 50 | 0 | 3 | 0.25 | A5 |
| Selenate (ppm)a | ||||
| 200 | 0 | 5 | 0.58 | A6 |
| 200 | 0 | 3 | 0.5 | A7 |
| 200 | 0 | 2 | 0.31 | A8 |
| 100 | 0 | 3 | 0.3 | A9 |
| 50 | 0 | 5 | 0.25 | A10 |
| 100 | 0 | 5 | 0.4 | A11 |
| 100 | 0 | 4 | 0.23 | A12 |
| 50 | 0 | 2 | 0.29 | A13 |
| 50 | 0 | 1 | 0.13 | A14 |
| 50 | 0 | 3 | 0.24 | A15 |
Table 1: Description of Se-rich algal samples grown in either a selenite or selenate-rich medium. aAll samples were grown in the
Table 1: Description of Se-rich algal samples grown in either a selenite or selenate-rich medium. aAll samples were grown in the Revolving Algal Biofilm system with BBM bWeek of harvest Cell Culture Caco-2 cells were grown at 37°C with 5% CO2 and DMEM containing 10% FBS. The cells were maintained by weekly passage by washing with PBS and then with 0.5% Trypsin EDTA to disassociate the cells after reaching 90% confluency. For the cell uptake experiments, cells were passed to 6-well plates in DMEM with 10% FBS at a cell count of 190,000 cells/mm3 per plate well.
Because serum used in the culture media contains trace concentrations of Se, the cells were depleted with Se to assure optimal uptake of Se. This was achieved by growing them with DMEM supplemented with 10% FBS for 2 days, followed by 5% FBS for 2 days, and then 0% FBS for 6 days. At the end of 6 days, the cells became Se-deficient as measured by GPx activity. We used these Se-deficient cells for Se uptake experiments.
For BA experiments, immediately after the serum reduction period, the cells were washed with PBS. They were then treated with 4 mL of serum free DMEM and 1 mL of in vitro digest supernatants containing 100 nmol/L of Se either from SeMet, Na2SeO3, Na2SeO4, or Se-enriched algal biomass, with duplicates of each treatment. Treatment remained on the cells for 24 h to induce GPx activity prior to harvesting with 1mL of cold buffer (50mM Tris-HCl, pH 7.5, 5mM ethylenediamine tetraacetic acid, and 1mM dithiothreitol) and then sonicated for 4 seconds on ice. The samples were stored at -80°C prior to being thawed and centrifuged at 10,000 x g for 15 minutes at 4°C for protein analysis using the PierceTM Coomassie Bradford Protein Assay Kit (Thermo Fisher Scientific CA, USA) and GPx analysis. Two uptake measurements were made from each duplicate digestion and the values were corrected to the total concentration of Se in the supernatant after digestion.
Statistical Analysis
The GPx activity (mean + SD) between Se-deficient and Se-sufficient cells was determined using Student t-test. Data of BAC (solubility) and BA (GPx activity) are shown as mean ± SD and the differences among groups were determined by ANOVA with Tukey multiple comparison test for BAC and BA.
Duplicate wells of each treatment were used in the cell model as well as duplicates for solubility and GPx testing. Differences among the treatments were considered significant at P<0.05.
Results and Discussion
The BAC is determined by solubility following in vitro digestion. The solubility (%) of the Se-containing samples was calculated based on the fraction of 200 µg of Se that was soluble following in vitro digestion. Figure 2A compares the solubility of the algae samples grown in various concentrations of selenite, SeMet and Na2SeO3. No significant differences were seen in mean solubility between SeMet, and Na2SeO3 (p>0.05) with 84.3% and 74.3%, respectively. The solubility of all Se-rich algae samples was significantly lower (p<0.05) compared to SeMet and Na2SeO3. The algae sample A1 had the highest mean solubility of 30.7%, which was significantly different from the other four algae samples (p<0.05), although the Se concentration is 50% of A2 (Table 1). Its solubility was approximately 36% of SeMet and its growth conditions consisted of 30 ppm selenite in media without added sulfite, and harvested at week 1. Compared to the other samples with 30 ppm selenite and 0 ppm sulfite, the growth conditions for A1 had a shorter cultivation time.
Figure 2B compares the solubility of the algae samples grown in various concentrations of selenate, SeMet and Na2SeO4 supplement. Again, Se solubility of SeMet and Na2SeO4, were not significantly different (p>0.05) with 84.3% and 87.1%, respectively. The Se-rich algae samples all had a much lower solubility and were significantly different compared to SeMet, and Na2SeO4, (p<0.05) (Figure 2).
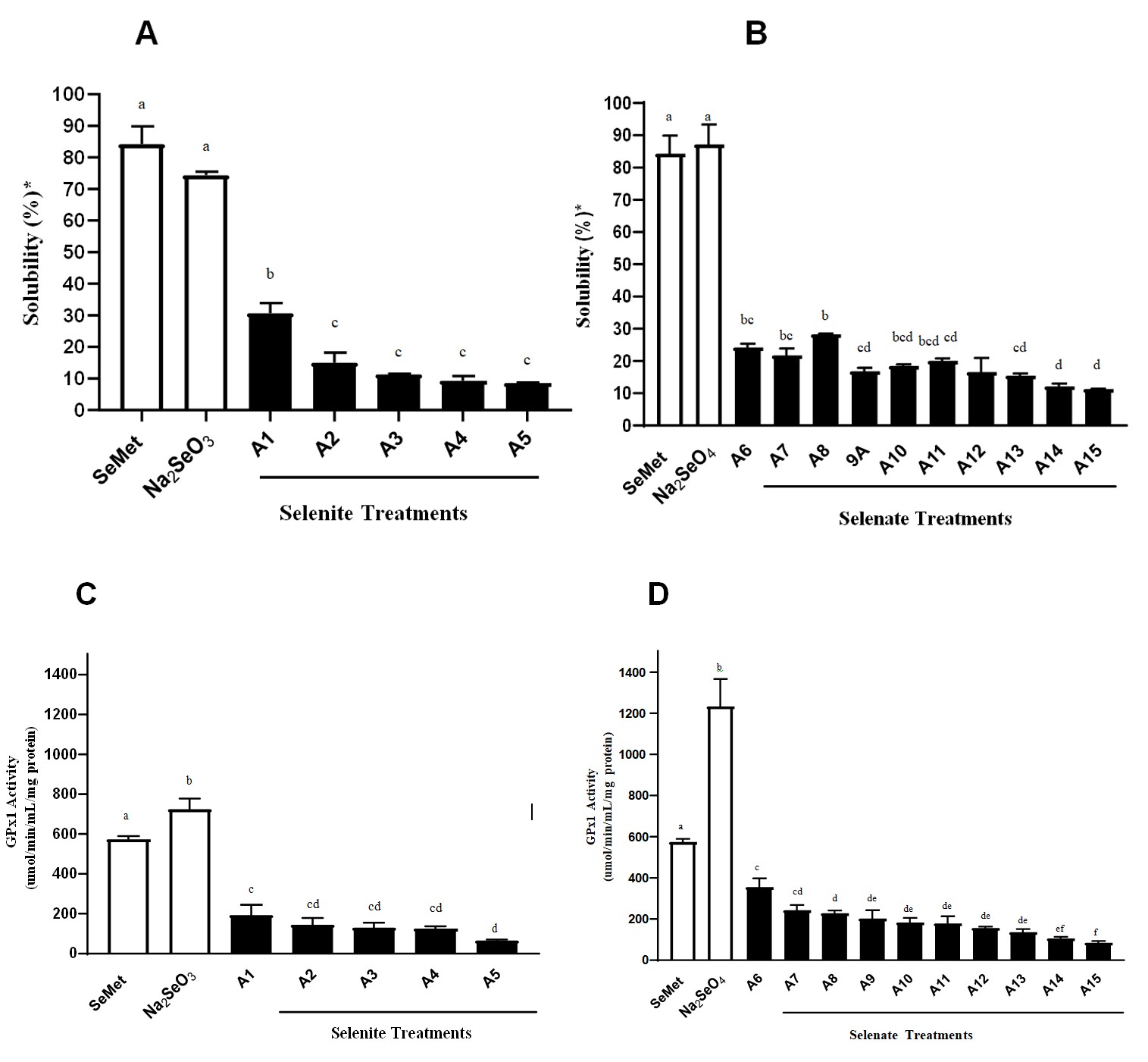
Figure 2: Bioaccessibility and bioavailability following in vitro digestion. Percent solubility (BAC) following in vitro digestion of algal samples grown in selenite (A) and selenate medium (B) compared to solubility of SeMet and their respective inorganic salts. The GPx activity (BA) of algal samples grown in selenite (C) and selenate (D) medium compared to GPx activity of SeMet and their respective inorganic salts. Data represent mean ± SD of duplicates for solubility and 3-4 replicates for GPX activity. Values not sharing the same letter are considered significantly different (P<0.05) based on ANOVA with Tukey multiple comparison test.
Sample A8 grown with 200 ppm selenate in media without sulfite, and harvested at week 2 of growth with 0.31% Se in its dry mass had the greatest average solubility (28.3%) and was significantly higher compared to sample A9 and A12-A15 (p<0.05). Its solubility was approximately 34% of the SeMet. These results could be explained by the higher concentration of selenate added to the growth medium, the shorter cultivation time compared to the other samples, and having no sulfite in the medium. Based on several previous studies, SeMet is recognized to be more soluble and therefore, have greater BAC compared to most other forms of Se, particularly selenite and elemental Se1 [29]. It is also recognized as the best form of Se to raise overall Se status in the body, as it is able to be stored, methylated, and can increase selenoprotein synthesis. Selenate, although inorganic, is comparable in BAC to organic Se, based on several published studies [1, 8, 19]. Selenite has lower BAC compared to organic forms of Se such as SeMet, SeCys, and methylselenocysteine [35].
Algae can successfully absorb larger quantities of inorganic Se and can metabolize it to organic forms, including selenoproteins [36, 37]. Other studies have reported that algae can generate methylated forms of Se [22, 38]. Currently it is not clear what type and how much organic Se is formed following uptake of inorganic Se, creating challenges for assessing the BA37. The composition of algae may play a major role in how Se is metabolized and utilized. Fiber and sulfur content, for example, could inhibit Se absorption, particularly organic Se(17). Lastly, pH of the simulated digestion could also might play a role in Se BA assessment. The pH 6 that was used in our in vitro digestion protocol was reflective of duodenal pH, which is commonly used for minerals [39] but, Se is absorbed to the greatest extent in the jejunum [16, 40]. Perhaps using a slightly higher pH to mimic further down the small intestine could increase absorption of all the treatments, but a different cell line has to be used for this. However, our results for solubility with the SeMet and inorganic Se-salts are consistent with current literature [1, 13, 29].
To assess the BA of the Se-rich algae, we utilized GPx activity in the cell extracts after Se was taken up by the cell. Figure 3 shows the Se-deficient cells had approximately a 70% reduction in GPx activity, which was significantly lower compared to the Se-sufficient cells (P<0.05). These results were slightly different from a previous published study that showed an 80% reduction in GPx activity in their Se- deficient model [29]. The slight differences in methodology might be responsible for the results. It was assumed that not all forms of Se, although having similar BAC, would induce GPx to the same extent. Concentration and chemical form are key factors regarding the ability for Se to induce GPx activity. Since GPx activity might vary with concentration of Se, use of same concentration of 100 nmol/L of Se in each cell treatment in our study allowed for better comparison among the samples.
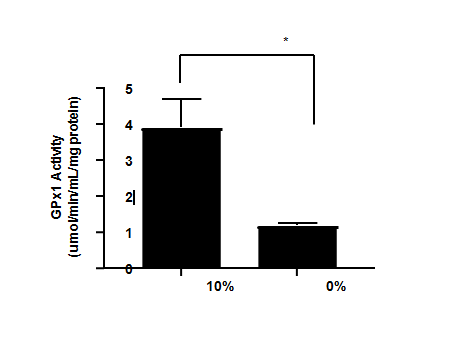
Figure 2C compares the GPx activity of the algae samples, cultivated in selenite under different conditions as described in the methods. Compared to SeMet and Na2SeO3, Na2SeO3 had the highest GPx activity 725 mU/mg cell protein which was 126% significantly higher (p<0.05) than SeMet (574.1 mU/mg cell protein). Cells treated with algae grown in selenite had a mean GPx activity that was 11-34% (p<0.05) of the SeMet treatment and all of the algae samples had a significantly lower (p<0.05) GPx activity compared to Na2SeO3, therefore, it was not possible to identify any of the algae samples as superior. Sample A1 did appear to have a better overall GPx activity but it was only significantly different (p<0.05) in GPx activity to sample A5 (lowest BA), which was cultivated in a higher concentration (50 ppm) of selenite and harvested at 3 weeks.
Figure 2D compares the GPx activity of the algae samples grown in selenate to SeMet and Na2SeO4. The GPx activity of Na2SeO4 was 214% higher than SeMet (p<0.0001). Cells treated with algae grown in selenate had a mean GPx activity that was 15-62% (p<0.05) of the SeMet treatment and all of the algae samples had a significantly lower (p<0.05) GPx activity compared to Na2SeO4. Sample A6 (grown in 200 ppm selenite without sulfite, and harvested at week 5 of growth with 0.58% Se in dry mass) had the highest GPx activity (354 mU/mg cell protein) and was significantly different (p<0.05) compared to the other algae samples, except for sample A7 which had the same growth conditions but was harvested at week 3. In general, the algae grown in the higher concentration of selenate (200 ppm) with no sulfite appeared to have a better BA than all other algae samples.
It is clear that Se solubility may not be the best predictor of the Se BA. The inorganic selenite and selenate showed higher GPx activity compared to SeMet which could be explained by the direct reduction to H2Se before incorporation into selenoproteins, compared to the longer pathway that organic Se uses [11, 41, 42]. Organic Se is not directly transformed to H2Se and may go through other routes such as incorporation into body proteins in place of methionine [43]. Regardless of concentration or the form of Se, BA from Se-rich algal samples was very low compared to organic and inorganic Se. Based on the difference in BA between organic and inorganic Se, it is possible that algae may metabolize and absorb the forms of Se. As with solubility, considering factors such as algal composition could further provide the understanding of its BA. Algal biomass could simply be more difficult to digest and finding other ways to improve the digestion could improve the BAC, and perhaps the BA.
RAB reactors generally showed significantly higher Se removal efficiency and higher Se content in the biomass than bubble column. Such an advantage could be attributed to the extracellular polymer substances (EPS) produced in the algal biofilm. Composed of polysaccharide, protein, and other organic compounds, EPS is reported to be capable of binding significant quantities of metals [44, 45, 46]. It is very likely that EPS-bound Se may account for a large percentage of the total Se bound to the biofilm in RAB reactor, which might be in inorganic form. This Se adsorbed to the algal biofilm may be tightly bound to the EPS and may not be released during digestion. This could explain the algae having low BAC and BA compared to their respective salts. Some biomass pretreatment strategies including grinding may be helpful to improve the Se-rich algae digestion. In addition, it is very intriguing that the highest BA and BAC was received when the biofilm was cultured under low concentration (10 ppm) of Se from sodium selenite or high concentration (200 ppm) of Se from sodium selenate. Optimization of selecting the form of Se salts and concentrations in the algal culture medium and understanding of the mechanisms of Se assimilation would be necessary to enhance the BAC and BA of algal-based Se.
In addition to providing Se by algal biomass, Chlorella sp. has much to offer for nutritional benefits because it contains essential amino acids, monounsaturated and polyunsaturated fatty acids, fiber, vitamins, and minerals [24, 26]. It can have a positive environmental impact based on its ability to remove nutrients that are toxic at higher concentrations [31]. Because of how well algae can be cultivated with one or more particular nutrients such as Se, it is promising to use for biotechnological purposes, such as creating a natural supplement for humans or animals. In addition, a Se-rich alga is emerging in research for its potential to remove Se from wastewaters and areas in the environment that have high Se concentration.
In conclusion, Se deficiency can be induced in the Caco-2 cells by gradual depletion of Se in the media and was found to be a good model to assess BA. It appears that a higher concentration of selenate needs to be added to the media compared to the selenite to produce Se-rich algae. Although, the BA of the algae grown in selenate or selenite had significantly less than the SeMet treatment, selenate treated samples performed better than selenite samples. In general, the algae grown in the higher concentration of selenate (200 ppm) with no sulfite appeared to have a better BA than all other algae samples.
Although this study did not show the BAC and BA of Se- rich algae nearly as good as traditional forms of Se used in supplementation, our study nonetheless provides an insight for future research for improving BA of Se from this novel source.
Author Information
The authors declare no conflict of interest
Funding Sources
This study was funded by the United States Department of Agriculture
References
-
Finley JW (2006) Bioavailability of selenium from foods. Nutr Rev 64(3): 146-151.
-
Daniels LA (1996) Selenium metabolism and bioavailability. Biol. Trace Elem. Res 54(3): 185-199.
-
Rayman MP (2013) Stranges, S. Epidemiology of selenium and type 2 diabetes: can we make sense of it? Free Radic Biol Med 65: 1557-1564.
-
Tan M, Li S, Swaroop M, Guan K, Oberley LW, Sun Y (1997) Transcriptional activation of the human glutathione peroxidase promoter by p53. J Biol Chem 274(17): 12061-12066.
-
Sonet J, Bierla K, Bulteau AL, Lobinski R, Chavatte L (2018) Comparison of analytical methods using enzymatic activity, immunoaffinity and selenium- specific mass spectrometric detection for the quantitation of glutathione peroxidase 1. Anal Chim Acta 1011: 11-19.
-
Forman HJ, Zhang H, Rinna A (2009) Glutathione: overview of its protective roles, measurement, and biosynthesis. Mol Aspects Med 30(1-2): 1-12.
-
Bodnar M, Szczyglowska M, Konieczka P, Namiesnik J (2016) Methods of selenium supplementation: bioavailability and determination of selenium compounds. Crit Rev Food Sci Nutr 56(1): 36-55.
-
Thiry C, Ruttens A, De Temmerman L, Schneider YJ, Pussemier L (2012) Current knowledge in species- related bioavailability of selenium in food. Food Chem 130(4): 767-784.
-
Fernández-Martínez A, Charlet L (2009) Selenium environmental cycling and bioavailability: a structural chemist point of view. Rev Environ Sci Bio 8(1): 81-110.
-
Dimkovikj A, Hoewyk D Van (2014) Selenite activates the alternative oxidase pathway and alters primary metabolism in _Brassica napus_ roots: evidence of a mitochondrial stress Response. BMC Plant Biol.
-
Suzuki KT, Shiobara Y, Itoh M, Ohmichi M (1998) Selective uptake of selenite by red blood cells. Analyst 123: 63-67.
-
Cases J, Vacchina V, Napolitano A, Caporiccio B, Besançon P, et al. (2001) Selenium from selenium-rich spirulina is less bioavailable than selenium from sodium selenite and selenomethionine in selenium-deficient rats. J Nutr 131(9): 2343-2350.
-
Bierla K, Szpunar J, Yiannikouris A, Lobinski R (2012) Comprehensive speciation of selenium in selenium-rich yeast. TrAC - Trends Anal Chem 4: 122-132.
-
Suzuki KT, Kurasaki K, Ogawa S, Suzuki N (2006) Metabolic transformation of methylseleninic acid through key selenium intermediate selenide. Toxicol Appl Pharm 215 (2): 189-197.
-
Soriano-García JF, Torras-Llort M, Ferrer R, Moretó M (1998) Multiple pathways for L- methionine transport in brush-border membrane vesicles from chicken jejunum. J Physiol 509(2): 527-539.
-
Nickel A, Kottra G, Schmidt G, Danier J, Hofmann T, et al. (2009) Characteristics of transport of selenoamino acids by epithelial amino acid transporters. Chem Biol Interact 177(3): 234-241.
-
Thiry C, Ruttens A, Pussemier L, Schneider YJ (2013) An in vitro investigation of species- dependent intestinal transport of selenium and the impact of this process on selenium bioavailability. Br J Nutr 109(12): 2126-2134.
-
McConnell KP, Cho GJ (1965) Transmucosal movement of selenium. Am J Physiol 208(6): 1191-1195.
-
Vendeland SC, Butler JA, Whanger PD (1992) Intestinal absorption of selenite, selenate, and selenomethionine in the rat. J Nutr Biochem 3(7): 359-365.
-
Mehdi Y, Hornick JL, Istasse L, Dufrasne I (2013) Selenium in the environment, metabolism and involvement in body functions. Molecules 18(3): 3292- 3311.
-
Suhajda Á, Hegóczki J, Janzsó B, Pais I, Vereczkey G (2000) Preparation of selenium yeasts I. Preparation of selenium-enriched Saccharomyces cerevisiae. J Trace Elem Med Biol 14(1): 43-47.
-
Bodnar M, Szczyglowska M, Konieczka P, Namiesnik J (2016) Methods of selenium supplementation: bioavailability and determination of selenium compounds. Crit Rev Food Sci Nutr 56(1): 36-55.
-
Rajashree K, kumar TM (2013) Preparation of selenium tolerant yeast saccharomyces cerevisiae. J Microbiol Biotechnol Res 3(3): 46-53.
-
Wells ML, Potin P, Craigie JS, Raven JA, Merchant SS, et al. (2017) Algae as nutritional and functional food sources: revisiting our understanding. J Appl Phycol 29(2): 949-982.
-
Mason R (2001) Chlorella and spirulina green supplements for balancing the body. J Altern Complement Med 7(3): 161-165.
-
Petkov G, Garcia G (2007) Which are fatty acids of the green alga _Chlorella_? Biochem Syst Ecol 35(5): 281-285.
-
Pratoomyot J, Srivilas P, Noiraksar T (2005) Fatty acids composition of 10 microalgal species. J Sci Technol 27(6): 179-1187.
-
Gross M, Henry W, Michael C, Wen Z (2013) Development of a rotating algal biofilm growth system for attached microalgae growth with in situ biomass harvest. Bioresour Technol 150: 195-201.
-
Zeng H, Botnen JH, Johnson LK (2008) A selenium- deficient caco-2 cell model for assessing differential incorporation of chemical or food selenium into glutathione peroxidase. Biol Trace Elem Res 123(1-3): 98-108.
-
Pavlata L, Pechová A, Illek J (2000) Direct and indirect assessment of selenium status in cattle - a comparison. Acta Vet Brno 69: 281-287.
-
Staicu LC, Van Hullebusch ED, Rittmann BE, Lens PNL (2017) Industrial selenium pollution: Sources and biological _Treatment Technologies_. Bioremediation of selenium contaminated wastewater, pp: 75-101.
-
Gross M, Wen Z (2014) Yearlong evaluation of performance and durability of a pilot-scale revolving algal biofilm (RAM) cultivation system. Bioresour Technol 171: 50-58.
-
Gross M, Mascarenhas V, Wen Z (2015) Evaluating algal growth performance and water use efficiency of pilot‐ scale revolving algal biofilm (RAB) culture systems. Biotechnol. Bioeng 112(10): 2040-2050.
-
Proulx AK, Reddy MB (2006) Iron bioavailability of hemoglobin from soy root nodules using a caco-2 cell culture model. J Agric Food Chem 5(4): 1518-1522.
-
Takahashi K, Suzuki N, Ogra Y (2017) Bioavailability comparison of nine bioselenocompounds In vitro and in vivo. Int J Mol Sci 18(3): 506.
-
Doucha J, Lívanský K, Kotrbáček V, Zachleder V (2009) Production of chlorella biomass enriched by selenium and its use in animal nutrition: A review. Appl Microbiol Biotechnol 83: 1001-1008.
-
Schiavon M, Ertani A, Parrasia S, Vecchia FD (2017) Selenium accumulation and metabolism in algae. Aquat. Toxicol 189: 1-8.
-
Rayman MP (2004) The use of high-selenium yeast to raise selenium status: how does it measure up? Br J Nutr 92(4): 557-573.
-
Swain JH, Tabatabai LB, Reddy MB (2002) Histidine content of low-molecular-weight beef proteins influences nonheme iron bioavailability in caco-2 cells. J Nutr 132(2): 245-251.
-
Korpela H (1989) Effect of ascorbic acid on the bioavailability of sodium selenite in selenium-depleted rats as assessed by serum selenium concentration and glutathione peroxidase activity. Nutr Res 9(8): 937-940.
-
Nancharaiah YV, Lens PNL (2015) Selenium biomineralization for biotechnological applications. Trends Biotechnol 33(6): 323-330.
-
Birringer M, Pilawa S, Flohé L (2002) Trends in selenium biochemistry. Nat Prod Rep 19(6): 693-768.
-
Burk RF (1991) Molecular biology of selenium with implications for its metabolism. FASEB J 5(9): 2274- 2279.
-
Sheng G, Xu J, Luo HW, Li WW, Li WH, et al. (2013) Thermodynamic analysis on the binding of heavy metals onto extracellular polymeric substances (EPS) of activated sludge. Water Res 47(2): 607-614.
-
Liu Y, Lam MC, Fang HH (2001) Adsorption of heavy metals by EPS of activated sludge. Water Sci Technol 43(6): 59-66.
-
Vijayaraghavan K, Padmesh TVN, Palanivelu K, Velan M (2006) Biosorption of nickel (II) ions onto Sargassum wightii: Application of two-parameter and three- parameter isotherm models. J Hazard Mater 133(1): 304-308.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends