Phenolic Acids in Red Wine Interact Directly with the Protein Fraction of Saliva
Wine-evoked astringency is usually associated with interactions between proanthocyanidins and salivary proteins. Limited information exists about interaction of other wine phenolic compounds with saliva. This study assessed the ability of single phenolic acids and mixes of phenolic acids to interact with salivary protein. Protein diffusion on cellulose membranes and protein precipitation assays were conducted. Single phenolic acids (PA), excepting tannic acid (T), showed concentrationdependent reduction of the area of diffusion of the diffusible salivary protein. PAs were weak precipitants of the salivary protein but strong inducers of soluble PA-salivary protein complexes. T precipitated non-diffusible salivary proteins without effect on the diffusible protein. Compared to single PAs, mixes of gallic acid (G) and PAs showed similar effects regarding inhibition of protein diffusion and formation of soluble PA-protein complexes. Contrarily, T interfered with both effects of PAs. It is concluded that PAs interact directly and diversely with the salivary protein and thence may cause astringency
Introduction
Phenolic acids are non-flavonoid polyphenols that are widely spread throughout the plant kingdom. They are particularly abundant in flesh and skins of grape berries (around 50-200 mg/kg) and in red wine (150 mg/L) [1, 2, 3]. Benzoic acids (C6-C1) and cinnamic acids (C6-C3) are the two most common groups of phenolic acids. In wine, vanillic acid, protocatechuic acid and gallic acid are well-known abundant benzoic acids while ferulic acid, p-coumaric acid and caffeic acid are typical examples of cinnamic acids [4]. Some authors also include tannic acid in the group of wine phenolic acids because of its very high contents of gallic acid and ellagic acid residues that are sterified to one or several OH groups of a single molecule of glucose and because it is a common component of wood-aged wines [5, 6, 7]. Phenolic acids have been shown to contribute diversely and positively to wine quality by acting as color stabilizing copigments, antioxidants and free radical scavengers [8, 9, 10, 11]. However, these polyphenols may be also potentially detrimental to wine aroma and flavor following microbial degradation and to wine color through their enzymatic or chemical oxidation leading to hazing [1, 12].
On the other hand, the major and diverse family of polyphenols has been widely associated to a number of sensory features of grape and wine, including astringency [13]. Since Bate-Smith [14], abundant evidence has accumulated that polyphenol-evoked astringency is associated with physicochemical interactions between particular families of polyphenols (proanthocyanidins and hydrolizable tannins) and some families of salivary proteins (proline-rich proteins and histidin-rich proteins or histatins) [15, 16]. Hydrogen bonds between numerous OH groups on the polyphenols and lateral chains of certain aminoacid residues, such as proline, on the salivary proteins, together with hydrophobic interactions involving stacking of aromatic rings from both types of molecules are core mechanisms underlying protein-polyphenol interactions [15]. Those interactions would result in extensive alteration in the salivary pellicle covering the oral surfaces, which in turn is transduced by oral mechanoreceptors and sensorially perceived as tightness, dryness and roughness on the oral surfaces [13, 17, 18, 19, 20, 21, 22, 23, 24, 25].
Certainly, such molecular interactions that are putatively responsible for triggering astringency would not be applicable to a number of astringent molecules lacking OH groups, such as alums and inorganic acids. Accordingly, the quest for further insights into physicochemical mechanisms accounting for astringency still remains as an active research focus since three decades ago [13]. Thus, apart from the hydrophobic interactions between aromatic rings being part of salivary proteins and polyphenols as well as hydrogen bonding in local hydrophobic molecular environments, a number of studies have documented a major influence of pH on polyphenol-protein interactions and thence on polyphenol-provoked astringency [13, 26, 27, 28, 29, 30]. In this regard, pH would influence the number of hydrogen-bond forming sites by inducing conformational changes in the molecular species forming hydrogen bonds [31, 32].
In the case of wine-evoked astringency, such general effect of pH is assumed to be provided by the abundant and diverse fraction of non-phenolic acid species of wine, including tartaric, malic and citric acids. In fact, strong acids lacking OH groups, such as hydrochloride acid, have been shown to be powerful inducers of astringency (relative to its sour taste) when compared to weak organic acids [33]. Altogether, considering that phenolic acids are a quantitatively significant component of the wine matrix and that they display several independent structural characteristics of relevance for astringency, such as acidic character (pKa around 4-5), presence of aromatic C-bound OH groups with the ability to form hydrogen bonds, and presence of at least one hydrophobic aromatic ring, the wine phenolic acids may well have the ability to interact in a physicochemical manner with proteins. In fact, recent studies have shown that some wine phenolic acids of the hydroxybenzoic and hydroxycinnnamic families tested either individually or as mixtures do interact directly and differentially with a model pure proline-rich peptide (IB712), thus providing strong support to the hypothesis that other members of the wine matrix, apart from proanthocyanidins and hydrolyzable tannins, should be considered in the molecular study models addressed to understand polyphenol-evoked astringency [34, 35, 36].
Accordingly, those interactions of phenolic acids with single well-defined peptides should be now stepped up and complemented by the assessment of their interactions with the bulk of human salivary protein. Human saliva is a complex dynamically structured supramolecular matrix. Hundreds of proteins and mucins secreted by a diversity of major and minor salivary glands form a salivary pellicle that covers all hard and soft tissues in the mouth [37, 38]. Such would be the physicochemically interactive semi-solid proteinaceous structure participating through a barely known mechanism in evoking the mouth-feeling sensation of astringency [37]. In the present study we have compared the ability of a group of phenolic acids to interact in vitro with the protein component of a standard and expectedly invariable sample of human saliva. The protein component of undiluted or water-diluted whole human saliva placed on a cellulose membrane shows a biphasic mode of diffusion comprising a diffusible and a non- diffusible component that can be revealed by staining with a high affinity protein dye, such as Coomassie blue [30]. In the present study, individual or dual mixtures of phenolic acids were tested for their abilities to interfere either with the diffusion of the protein component of saliva on a cellulose membrane or with its solubility [7]. Well-known astringent grape and wine polyphenols (hydrolyzable tannins and proanthocyanidins) have been shown to interfere with both parameters [39]. The eventual interaction of phenolic acids with fractions of salivary proteins is expected to shed further lights on their participation in astringency production.
Materials and Methods
Materials
Gallic acid (G), protocatechuic acid (P), ferulic acid (F), vanillic acid (V), caffeic acid (CA), p-coumaric acid (CU), tannic acid (T), bovine serum albumin (BSA) and Coomassie blue R-250 were acquired from Sigma-Aldrich (Saint Louis, MO, USA). Whatman (R) grade 1 cellulose sheets were obtained from Whatman Ltd (Maidstone, England). Proanalysis grade trichloacetic acid, ethanol and acetic acid were obtained from Merck KGaA (Darmstadt, Germany).
Saliva Collection
Unstimulated whole saliva was freshly collected from a 24-year old female volunteer, without history of smoking, alcoholism or medication consumption, with no evidence of disease and displaying both normal saliva flow (over 1mL/ min) and normal salivary protein profile [40]. Immediately before the experiments (always between 9:00 and 11.00
A.M. to minimize eventual diurnal variations) and following a mouth rinse with water, the saliva accumulated for 1 minute in the oral cavity was collected in a sterile Falcon tube. After spinning the tube in a refrigerated centrifuge (4°C) at 650 g for 3 min, the supernatant representing saliva was kept in an ice-bath during the experiment. An aliquot of saliva was routinely saved at -80°C to measure protein concentration by the method of Bramhall using bovine serum albumin as reference [41].
Albumin Working Solution
Bovine serum albumin (BSA) (100 mg) was dissolved in distilled water (80 mL) at 35-40°C with constant mechanical stirring for 24 hours. Following centrifugation (650 g x 5 min), the supernatant was passed through a membrane disc filter, 0.45 µm pore size and 25 mm diameter. The absorbance of the filtrate at 280 nm was adjusted to 0.7 AU (1 mg/mL) with distilled water [7]. The resulting solution was stored at -20°C as 1-ml aliquots. Single aliquots were thawed and used just once in the study.
Phenolic Acid (PA) Solutions
Stock solutions (5 mg/mL) of each PA were prepared in 30% (v/v) ethanol. pH was not adjusted. Right before the assays, stock solutions were diluted with the same hydroalcoholic solvent to produce the corresponding working solutions.
Interaction of Phenolic Acids with the Salivary Protein Fraction
Diffusion and precipitation assays of mixes of phenolic acids with freshly collected saliva (or BSA, for reference) were conducted as described elsewhere [7]. For the diffusion assay, 150 µL of a series of solutions of each phenolic acid (range 0-3 mg/mL) were mixed with 150 µL of whole saliva (or BSA) using mechanical stirring on a Vortex mixer for 15 sec (binary mixes). Twenty-µL aliquots of the resulting mixtures were transferred with a precision micropipette on a point of a horizontally suspended cellulose membrane and allowed to freely diffuse. The borders of the wet diffusion area were demarcated with graphite. The air-dried membrane was fixed, stained for protein with Coomassie blue and revealed [7]. Roughly circular blue spots represented the distribution areas of the salivary protein (or BSA). The membrane was imaged and the areas of the blue spots were measured by using Image J v1.45 software [42]. In this diffusion assay, compared to the control condition (absence of phenolic acid in the mixture), decreased diffusion area of individual spots is taken as indicative of PA-protein interaction. For the precipitation assay, each binary PA-saliva mixture (or PA-BSA mixture) used in the diffusion assay was spun in a refrigerated centrifuge at 650 g x 5 min and 20-µL aliquots of the corresponding supernatants were spotted on a cellulose membrane. This was fixed, stained and processed as above. In this precipitation assay, compared to the control condition, reduced staining of individual spots with Coomassie blue is taken as indicative of lower amount of protein in the supernatant, that is, occurrence of protein precipitation. Interaction between PAs and the salivary protein fraction was assessed by mixing saliva with either one PA (binary mixes) or with one PA plus either gallic acid or tannic acid (ternary mixes).
Statistics
ANOVA analysis with Infostat® Statistical Software (v 2013) was initially used. In case of significant differences between samples, a Tukey’s multiple range test at the 0.05 level of significance was conducted.
Results
Individual Phenolic Acids affect Diffusion of the Protein Fraction of Saliva on Cellulose Membranes
A 1:1 dilution of saliva with 30% ethanol placed on a cellulose membrane and revealed with a selective protein- staining dye displayed a biphasic mode of diffusion consisting of a non-diffusible protein fraction (NDF) surrounded by a weaker stained strip of freely diffusible protein fraction (DF) (Figure 1), control. When growing amounts of phenolic acids in the 0-3 mg/mL range of concentrations were mixed with saliva, the areas of diffusion of the diffusible protein fraction were progressively and significantly reduced (anti-diffusive effect) to as much as 65% with respect to the area of diffusion displayed by the salivary protein alone (control condition) (Tukey test, p < 0.05) (Figure 1). Contrarily, the area of the non-diffusible salivary protein fraction was unaffected by the presence of any of the PAs. Comparatively, the anti-diffusive effects produced by the series of concentrations of the whole group of PAs were highly similar to each other in such a manner that the diffusible fraction could be consistently noticed up to concentrations of 1.2-1.4 mg/mL. Above those concentrations, single PAs provoked full disappearance of the diffusible salivary protein that was accompanied by a noticeable increase in the staining intensity of the non- diffusible protein fraction. Unlike the rest of PAs, tannic acid showed neither anti-diffusive effect on the diffusible salivary protein fraction nor increase in the staining intensity of the non-diffusible salivary protein (Figure 1).
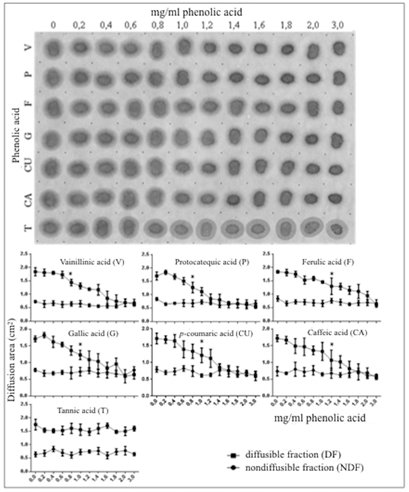
Figure 1: Effect of growing concentrations of phenolic acids on diffusion of the salivary protein on cellulose membranes. Series of 1:1 v/v phenolic acid/saliva mixes were spotted on a cellulose membrane and assayed for protein diffusion as described under Materials and Methods. Upper panel: Numbers on top represent concentrations of phenolic acids before mixing with saliva. Letters on the left stand for individual phenolic acids: V, vanillic acid; P, protocatechuic acid; F, ferulic acid; G, gallic acid; CU, p-coumaric acid; CA, caffeic acid and T, tannic acid. Note the progressive reduction of the total diffusion area of the salivary protein with increasing concentrations of phenolic acids, excepting tannic acid. Lower panel: Planimetric assessment of the anti-diffusive effect of phenolic acids on the diffusible protein fraction (DF) and lack of effect on the non-diffusible protein fraction (NDF) of saliva. Each point on the curves represents average ± standard deviation (triplicates). The asterisk on each curve represents the minimal concentration of phenolic acid resulting in a significant difference respecting the control condition (solvent with no phenolic acid).
For comparison, the diffusion assay was reproduced by replacing saliva with a pure highly diffusible model hydrophilic protein, bovine serum albumin (“albumin”). Unlike saliva, a solution of pure albumin behaves as a homogeneously diffusing protein on cellulose membranes (Figure 2). In this study, the series of growing concentrations of PA solutions was shown to interfere with albumin diffusion even at the lowest tested concentration (0.2 mg/mL). Such anti-diffusive effect was progressive in direct relation with the PA concentration. Only minor quantitative differences between the anti-diffusive effects of different PAs could be appreciated. By contrast, in this assay with albumin the effect of tannic acid differed drastically from the one observed with the salivary protein. In effect, at a concentration as low as 0.4 mg/mL, tannic acid provoked an almost complete suppression of albumin diffusion on the cellulose membrane (Figure 2).
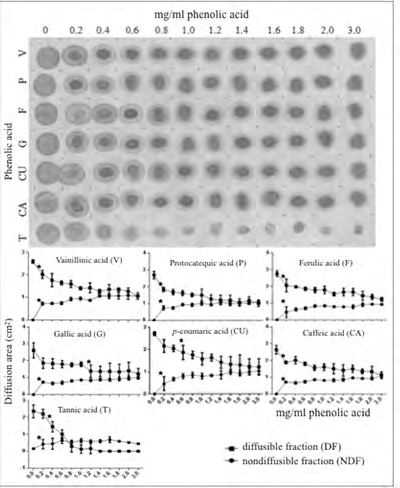
Figure 2: Effect of growing concentrations of phenolic acids on diffusion of albumin on cellulose membranes. The experiment is identical to the one of Figure 1 except that saliva was replaced by albumin in solution. Symbols in this figure have the same meaning as those given in the corresponding legend to Figure 1. Note in the upper panel the progressive but somewhat diverse reduction of the diffusion area of albumin and the appearance of a non-diffusible protein fraction with increasing concentrations of different phenolic acids. Note also the intense anti-diffusive effect of tannic acid on the protein. In the lower panel a planimetric assessment of the diffusion areas for each experimental condition is shown.
Effect of Individual Phenolic Acids on the Solubility of the Salivary Protein
When aliquots taken from supernatants (600 g x 5 min) of the series of PA-saliva reaction mixtures described in the experiment of Figure 1 were spotted on a cellulose membrane, protein staining remained usually intense, thus indicating that the PAs in the study are weak precipitating agents for the salivary protein fraction (Figure 3). In this precipitation assay, the diffusion area of the salivary protein remaining in the supernatant was also progressively reduced as the amount of PA mixed with saliva was increased (Figure 3). Altogether, such observations are consistent with interaction of PAs with the salivary protein and with occurrence of soluble PA-salivary protein complexes in the supernatants. Most of the PAs in the study showed similar effects. In contrast, under the experimental conditions in the study, tannic acid at concentrations above 1.2 mg/mL was able to fully precipitate the non-diffusible salivary protein fraction (Figure 3). Accordingly, both the insoluble and the soluble PA-salivary protein complexes in this precipitation assay would point to physicochemical interactions between PAs and salivary protein components.
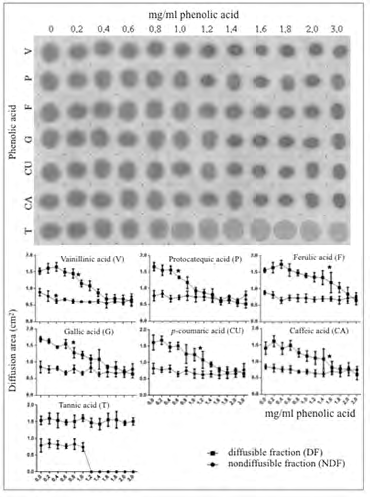
Figure 3: Effect of growing concentrations of phenolic acids on the precipitation of salivary protein. Samples of supernatants from the series of 1:1 v/v phenolic acid/saliva mixes were spotted on a cellulose membrane and assayed for the presence of protein as described under Materials and Methods. Symbols in this figure have the same meaning as those given in the previous legends to figures. Note in the upper panel the continuous presence of non-precipitated salivary protein together with a progressive reduction of its diffusion area in direct relation with increasing concentrations of phenolic acids. Unlike the other phenolic acids, medium concentrations of tannic acid fully precipitated the non-diffusible salivary protein fraction. As in previous figures, the planimetric assessment of the anti-diffusive effect of phenolic acids on the diffusible protein fraction (DF) together with their lack of significant effect on the non-diffusible protein fraction (NDF) of saliva are shown in the lower panel.
Crosstalk in the Interaction of some Phenolic Acids with the Salivary Protein Fraction
In order to examine whether the observed effects of individual PAs on the salivary protein fraction (binary mixes) could be influenced by the presence of other PAs, both diffusion and precipitation tests were conducted in presence of mixes of two PAs plus saliva (ternary mixes). Two different series of ternary mixes were tested. In the first series, saliva was mixed at 1:1 (volume/volume) ratios with working solutions of each one of the whole group of PAs previously mixed at 1:1 (weight/weight) ratios with gallic acid (G). The second series of ternary mixes was identical to the first one except that gallic acid was replaced by tannic acid (T). Comparisons were performed on the basis of similar total concentrations of PAs (range of 0-2 mg/mL) in the assays. Thus, the effects of each one of the PAs on the salivary protein fraction were compared with those produced by mixes of those PAs with either G or T. As shown in Figure 4, the diffusion test revealed that at any single concentration, each PA displayed an anti-diffusive effect on the salivary protein that was undistinguishable from the one produced by the corresponding two-component mix of PAs including G. For instance, at a concentration of 2 mg/mL, diffusion of the diffusible salivary protein was almost fully suppressed both by any pure PA and by the corresponding PA/G mix. By contrast, over the whole range of concentrations, T alone showed no effect on salivary protein diffusion but it did suppress the anti-diffusive effect produced by G alone. Accordingly, T displayed a sort of dominance on G concerning their differential effects on salivary protein diffusion. In a strict parallel experiment in which the two-component PA solutions comprised PA and T, with no exception T was shown to significantly reduce in a concentration-dependent manner the anti-diffusive effect displayed by each one of the PAs on the salivary protein (Figure 5).

Figure 4: Interaction effect of single phenolic acids (PA) with gallic acid (G) on both diffusion on cellulose membranes and precipitation of salivary protein. PA/saliva and (1:1 PA + G)/saliva mixes representing three different total concentrations of phenolic acids (0.5, 1.0 and 2 mg/ mL) were spotted on a cellulose membrane and assayed for protein diffusion (left panel). Supernatants from the same mixes were used to assess precipitation of the salivary protein (right panel) also using cellulose membranes as described under Materials and Methods. Numbers on top of the panels stand for total concentrations of phenolic acids before being mixed with saliva. Concentration zero represents saliva correspondingly diluted with the hydroalcoholic solvent used to dissolve phenolic acids. As in previous figures, letters on the left of the membrane represent individual phenolic acids. Note similar decreases in the diffusion area of the salivary protein in direct relation with the increasing total concentrations of either single phenolic acids or their mixtures with gallic acid (additive effect), excepting tannic acid. This one displayed no anti-diffusive effect on the salivary protein and a dominant suppressive effect over the anti-diffusive activity of gallic acid. The precipitation assay, which is focused on the assessment of the supernatants for each experimental condition, closely mirrored the outcome of the diffusion assay.
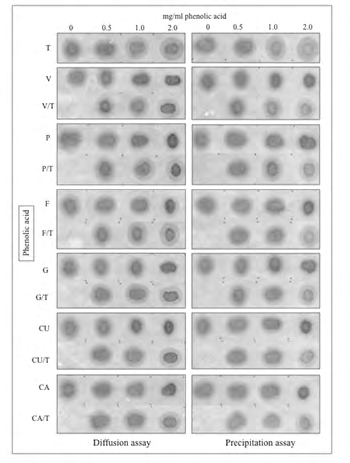
Figure 5: Interaction effect of single phenolic acids (PA) with tannic acid (T) on both diffusion on cellulose membranes and precipitation of salivary protein. PA/ saliva and (1:1 PA + T)/saliva mixes representing three different total concentrations of phenolic acids (0.5, 1.0 and 2 mg/mL) were spotted on a cellulose membrane and assayed for protein diffusion (left panel). Supernatants from the same mixes were used to assess precipitation of the salivary protein fraction (right panel) also using cellulose membranes. Letters on the left of the membranes represent individual phenolic acids, as described in the legends to previous figures. As in figure 4, note similar decreases in the diffusion area of the salivary protein in direct relation with the increasing total concentrations of every single phenolic acid, excepting tannic acid. By contrast, being part of mixes with any other phenolic acid, tannic acid partially reverted the anti-diffusive effect of the partner phenolic acid. Again, the precipitation assay, which is focused on the analysis of supernatants for each experimental condition, showed the low power of individual phenolic acids to precipitate the salivary protein and the appreciable suppressive effect of tannic acid on the anti-diffusive effect displayed by phenolic acids on the diffusible salivary protein.
Given the significant interaction effects between PAs concerning diffusion on cellulose membranes and solubility of the salivary protein, experiments were designed to assess whether such interactions could be observed using albumin as a reference pure protein. The representative Figure 6, involving protocatechuic acid (P), gallic acid (G) and tannic acid (T), shows that at either concentration in the study (i.e. 0.5, 1.0 or 2.0 mg/mL), the G/P mixes interfered with albumin diffusion to a similar extent as with G alone or P alone. Contrarily, when in this study T was used instead of G, a different interaction was observed. Thus, T alone displayed an anti-diffusive effect on albumin that was significantly stronger than the ones produced by G alone or P alone, whereas the T/G and T/P mixes, at similar total concentrations, showed even stronger anti-diffusive effects than the G/P mixes and similar to T alone. Assessments of interactions between PAs concerning their effects on albumin by means of the precipitation test showed identical results than the diffusion test, that is, T was a significantly more powerful precipitant for albumin compared to G or P, whereas the partial precipitating effect of the G/P mixes on albumin was roughly similar to that of G or P alone. At identical total concentrations, the T/G and T/P mixes were weaker precipitants for albumin than T alone. However, the still abundant protein remaining in the supernatants under the conditions of the experiment displayed a variably reduced ability to diffuse, thus suggesting the occurrence of of soluble PA-BSA complexes (Figure 6).

Figure 6: Interaction effect of a single representative phenolic acid (protocatechuic acid, P) with either gallic acid (G) or tannic acid (T) on both diffusion on cellulose membranes and precipitation of albumin as reference protein. Three different total concentrations (0.5, 1.0 and 2 mg/mL) of either single PAs (T, G and P) or 1:1 mixes of PAs (T + G, T + P and G + P) were mixed at a 1:1 v/v ratio with albumin working solution. Aliquots of the final mixes were spotted on a cellulose membrane and assayed for protein diffusion (left panel). Supernatants from the same final mixes were used to assess precipitation of albumin (right panel) also using cellulose membranes. Numbers on top of the panels stand for total concentrations of phenolic acids (PA) before being mixed with BSA. Concentration zero represents albumin correspondingly diluted with the hydroalcoholic solvent used to dissolve phenolic acids. Note similar mild anti-diffusive effects of P, G and P + G and a markedly stronger anti-diffusive effect of tannic acid (T). The anti-diffusive effects of T + P and T + G mixes resembled the one of T alone. In the precipitation test, G, P and G + P showed a similar weak effect, but the non-precipitated albumin became part of less diffusible protein aggregates. By contrast, T alone was able to almost fully precipitate albumin but in presence of G or P such precipitating activity was noticeably reduced. However, such non-precipitated albumin became part of still less diffusible protein aggregates.
Discussion
The results of the present study support the hypothesis that some common wine phenolic acids interact physicochemically with the protein component of saliva. In the study, the salivary protein was revealed by selective staining with a protein-binding dye on unreactive cellulose membranes [43]. In addition, interaction between phenolic acids and the salivary protein fraction was represented by either a decreased area of distribution of the protein on cellulose membranes (diffusion test) or by loss of the protein from the supernatant following centrifugation of the phenolic acid-saliva mixture (precipitation test) [7]. Individual phenolic acids were found to interfere with the diffusion on cellulose membranes displayed by the diffusible protein component of saliva in a concentration-dependent manner. Although exhibiting some minor quantitative differences, most of the tested phenolic acids showed highly similar anti- diffusive effects with the remarkable exception of tannic acid. In addition, most of phenolic acids were also shown to form soluble phenolic acid-protein complexes with a fairly poor ability to precipitate the salivary protein component. Again, tannic acid was a remarkable exception due to its ability to fully precipitate the non-diffusible salivary protein component, a distinct fraction which is characterized by its nil diffusion when whole saliva is placed on cellulose membranes. Most of the experiments in the study consisted in testing in vitro interactions between a wide range of concentrations of phenolic acids (0-3 mg/mL) and the salivary protein. Such range was chosen in the understanding that in-mouth interactions between proteins from whole saliva and wine polyphenols underlie astringency perception, that the overall content of polyphenols in wine is about 1.5 g/L and that the design of the in vitro study comprised an hydroalcoholic medium (ethanol 15%), with no presence of any other polyphenolic wine component, excepting the phenolic acid being tested. Comparison of eventual interactions of a list of phenolic acids with the salivary protein by assaying only usual concentrations of phenolic acids in wine (around 0.1-0.2 mg/L) might disregard the eventual influence of a more complex organic environment on the interactions. Naturally, phenolic acids are part of a much more complex organic wine matrix. Accordingly, by testing a wide range of concentrations of individual phenolic acids left open the possibility that some concentration- dependent effects could be experimentally observed. As a matter of fact, some interactions between phenolic acids and salivary proteins were concentration-dependent and were observed even at the lowest concentrations of phenolic acids in the study.
On the other hand, free-forms of phenolic acids were used throughout the study even though in grapes and wines different hydroxycinnamic acids (e.g. caffeic, p-coumaric and ferulic acid) can be also esterified to different extents with tartaric acid (to form caftaric, coutaric or fertaric acids, respectively) or with the glucose moieties of anthocyanin monoglucosides. Quite likely, the compositional profiles of these various molecular forms involving phenolic acids may vary extensively among grapes and wines because of the genetic background of grape varieties, vitivinicultural practices and edaphoclimatic conditions. Since physicochemical reactivities of those different molecular forms towards salivary proteins could also differ from each other, extrapolation from the present findings with pure compounds to their effects as part of a more natural and complex matrix calls for an appropriate degree of caution.
Another main feature of the study design to highlight concerns the source of saliva used in the various assays. It is well known that protein composition profiles and amount of saliva vary with individuals. However, most of the attributes of this biological fluid are shared by saliva from most of healthy individuals, including protein concentration (below 1 mg/mL), pH (around 6.8), hypotonic fluid, low sodium and chloride and high calcium and potassium concentrations compared to plasma, profiles of relative concentrations of various salivary protein families (acidic and basic proline-rich proteins, alpha-amylase, histatins), and so on. Consequently, all the assays in the study were carried out with saliva taken from a single subject and always under exactly the same experimental conditions so that the independent variable was not saliva, or its quality, but the various phenolic acids. Over that proteinaceous substrate the study was addressed to characterize interactions of phenolic acids with the whole salivary protein fraction (which is revealed by its high affinity for Coomassie blue R-250, a protein-binding dye) and not with a selected single protein or synthetic model polypeptide. Interestingly, unlike other mammals, whole human saliva displays a biphasic mode of protein diffusion on cellulose membranes comprising both a diffusible and a non- diffusible component [30]. In the present study, diffusion of the protein salivary component was significantly affected (reduced) by most of phenolic acids in the study (diffusion assay) whereas the non-diffusible salivary component was reproducibly precipitated (precipitation assay) by only one of the phenolic acids in the study (tannic acid, a well-known astringent compound). Such differences (and similarities) between the effect of different phenolic acids on fractions of salivary proteins, also observed in the study when saliva was replaced by a soluble reference protein (serum albumin), seem to be a reflection of the complexities involved in the interactions between polyphenols and human saliva and, possibly, in astringency triggering mechanisms.
Saliva is a highly complex supramolecular dynamically structured functional proteinaceous complex that coat as a film both hard and soft surfaces in the mouth [17, 37]. As such, saliva represents the first contact site for foods and drinks in the organism right before evoking a variety of sensory perceptions, including astringency [13, 17, 44]. In mouth, transient changes in the organization of the salivary pellicle would derive from its marked dilution and interaction with food components at the start of a signaling process towards brain domains where sensory processing takes place. In the present study, in vitro interactions of phenolic acids with diffusible and non-diffusible complex protein fractions of whole saliva can represent at least partly those interactions evoking sensory perceptions. In this same regard, independent comprehensive sensory studies addressed to learn about differences in perception of phenolic acids in relation with saliva features are highly desirable [33]. In our experience, the widely recognized perceived astringency of red wines and proanthocyanidin polyphenols correlate well with physicochemical interactions assessed by these methods [30, 45, 46]. In this regard, both hydroxybenzoic and hydroxycynnamic acids exhibit several of the structural aspects of both condensed and hydrolyzable tannins that have been associated with their abilities to interact with some salivary proteins as an initial step leading to the sensation of astringency [15]. Firstly, hydroxyl groups attached to the aromatic hydrocarbon domain of phenolic acids might form hydrogen bonds with hydrogen acceptor atoms in the side chains of some amino acid residues of polypeptide chains (e.g. Pro). In addition, phenolic acids could also participate in hydrophobic interactions through the stacking of their aromatic ring against the pyrrolidone ring of either proline or other aromatic amino acid residues (e.g. Phe, Tyr, Trp). In mouth (around pH 7), phenolic acids in the study (pKa 4-4.64) could also participate by means of their negatively charged carboxyl groups in ionic associations with positively charged sites in some amino acid residues with high isoelectric points (e.g. Lys, Arg, His). Using saturation-transfer difference nuclear magnetic resonance spectroscopy (STD-NMR) and molecular dynamics simulations, a recent study has provided solid evidence that some individual wine phenolic acids, tested either individually or as mixtures, do interact directly and differentially with a model pure proline-rich peptide (IB712) [34]. Certainly, to move the assessment of protein- ligand interactions from using single peptides to the use of full-sequence isolated proteins and thence to the salivary protein matrix poses today just another major challenge to the understanding of astringency triggering (Charlton, Baxter, Lilley, Haslam, McDonald & Williamson [47].
In the present study it has been shown that several common wine phenolic acids do interact with the protein fraction of saliva. In addition, the observed interactions of the salivary protein with phenolic acids revealed several outstanding features that may be of value for an eventual algorithm associated with wine astringency. In first place, marked quantitative differences in the interaction displayed by different phenolic acids, all of them with similar pKa values, would suggest that proton dissociation is not the plain mechanism accounting for the observed effects. Secondly, interactions of most of the phenolic acids are dependent on their concentrations, thus supporting the view that pH is not sufficient to account for their physicochemical effects. In third place, different proteins or complex mixtures of proteins can interact differently with a single phenolic acid. For instance, under the experimental conditions in the study, tannic acid interfered with albumin diffusion on cellulose membranes but not with that of the diffusible salivary protein fraction. In this case, an apparent much higher affinity of tannic acid for the non-diffusible protein fraction of saliva would have led to sequestration of the phenolic acid from the diffusible salivary protein component, and thence to unaltered free diffusion of the latter one on the cellulose membranes. Such observation seems to be representative of the structural and eventually functional complexity of the human salivary matrix respecting physicochemical interactions accounting for astringency [15, 16, 48, 49]. Last but not least, diffusion and precipitation assays in the study using ternary mixes, (i.e. saliva plus two different phenolic acids) have prompted the view that some phenolic acids (e.g. tannic acid) are “dominant” over other phenolic acids (e.g. gallic and protocatechuic acids) as to their ability to interact with the salivary protein. Apart from indicating that those direct and diverse interactions depend not only on the capability of the phenolic acid to release protons but on the conjugate base or on the prevailing undissociated phenolic acid structure, the study also lends support to the view that those interactions are additionally mediated by their structural role in the also highly complex supramolecular wine matrix [50]. Likewise, these physicochemical findings may well underlie both co-astringency effects and synergistic effects observed in sensory studies with phenolic mixtures involving phenolic acids [34]. Thus, phenolic acids should be taken as active direct participants in oral sensations mediated by their interaction with mouth and salivary proteins, and not only as non-specific secondary elements providing a suitable acid oral environment for physicochemical interactions of a constellation of taste- and texture-associated food compounds.
Acknowledgements
This study was supported by grants Fondecyt–Chile 1180975 and Enlace 22 VID-UChile (EOS).
References
-
Croft K (1998) The chemistry and biological effects of flavonoids and phenolic acids. Ann N Y Acad Sci 854: 435-442.
-
Souquet J, Labarbe B, Le Guernevé C, Cheynier V, Moutounet M (2000) Phenolic composition of grape stems. J Agric Food Chem 48(4): 1076-1080.
-
Vivas N, Nonier M (2003) Sur l’estimation et le quantification des composés phenoliques des vins. Bulletin de la Office International de la Vigne et du Vin 76: 281-303.
-
Câmara C, Bortoloti J, Scarminio I, Ballus C, Meinhart A, et al. (2013) Optimization of electrophoretic separations of thirteen phenolic compounds using single peak responses and an interactive computer technique. Journal of the Brazilian Chemical Society 24(11): 1744- 1753.
-
Bedran A, Yoo K, Ema K, Pashley D (2009) Mechanical properties of tannic-acid-treated dentin matrix. J Dent Res 88(9): 807-811.
-
Zhentian L, Jervis J, Richard F (2001) Use of methanolysis for the determination of total ellagic and gallic acid contents of wood and food products. J Agric Food Chem 49(3): 1165-1168.
-
Obreque Slier E, Mateluna C, Peña Neira A, López Solís R (2010) Quantitative determination of interactions between tannic acid and a model protein using diffusion and precipitation assays on cellulose membranes. J Agric Food Chem 58(14): 8375-8379.
-
Robbins RJ (2003) Phenolic acids in foods: an overview of analytical methodology. J Agric Food Chem 51(10): 2866-2887.
-
Flanzy C (2003) Enología: fundamentos científicos y tecnológicos. 2nd (Edn.), Madrid: Ediciones Mundi- Prensa.
-
Pannala AS, Chan T, O’Brien P, Rice C (2001) Flavonoid B-ring chemistry and antioxidant activity: fast reaction kinetics. Biochem Biophys Res Commun 282(5): 1161- 1168.
-
Chen CH, Wu MC, Hou CY, Jiang CM, Huang CM, et al. (2009) Effect of phenolic acid on antioxidant activity of wine and inhibition of pectin methyl esterase. Journal of the Institute of Brewing 115(4): 328-333.
-
Capello MS, Zapparoli G, Logrieco A, Bartowsky E (2017) Linking wine lactic acid bacteria diversity with wine aroma and flavour. Int J Food Microbiol 243: 16-27.
-
Bajec MR, Pickering GJ (2008) Astringency: Mechanisms and perception. Crit Rev Food Sci Nutr 48(9): 858-875.
-
Bate-Smith EC (1954) Flavonoid compounds in foods. Advances in Food Research 5: 261-300.
-
Bennick A (2002) Interaction of plant polyphenols with salivary proteins. Crit Rev Oral Biol Med 13(2): 184-196.
-
Sarni Manchado P, Canals Bosch JM, Mazerolles G, Cheynier V (2008) Influence of the glycosylation of human salivary proline-rich proteins on their interactions with condensed tannins. J Agric Food Chem 56(20): 9563-9569.
-
Gibbins HL, Carpenter GH (2013) Alternative mechanisms of astringency-What is the role of saliva? Journal of Texture Studies 44(5): 364-375.
-
(1989) ASTM, Standard Terminology to Sensory Evaluation of Materials and Products. American Society of Testing and Materials: Philadelphia.
-
Obreque Slier E, Peña Neira A, López Solís R, Zamora Marín F (2010) Tannin-protein interaction is more closely associated to astringency than tannin-protein complex precipitation: experience with two oenological tannins and gelatin. International Journal of Food Science and Technology 45: 2629-2636.
-
Pires MA, Pastrana LM, Fuciños P, Cristiano Abreu CS, Oliveira SM (2020) Sensorial perception of astringency: oral mechanisms and current analysis methods. Foods 9(8): 1124.
-
Poncet Legrand C, Cartalade D, Putaux JL, Cheynier V, Vernhet A (2003) Flavan-3-ol aggregation in model ethanolic solutions: Incidence of polyphenol structure, concentration, ethanol content and ionic strength. Langmuir 19(25): 10563-10572.
-
Poncet Legrand C, Doco T, Williams P, Vernhet A (2007) Inhibition of grape seed tannin aggregation by wine mannoproteins: Effect of polysaccharide molecular weight. American Journal of Enology and Viticulture 58: 87-91.
-
Soares SI, Gonçalves RM, Fernandes I, Mateus N, de Freitas V (2009) Mechanistic approach by which polysaccharides inhibit α-amylase/procyanidin aggregation. J Agric Food Chem 57(10): 4352-4358.
-
Soares S, Brandão E, Mateus N, de Freitas V (2017) Sensorial properties of red wine polyphenols: astringency and bitterness. Crit Rev Food Sci Nutr 57(5): 937-948.
-
Sun B, de Sá M, Leandro C, Caldeira I, Duarte FL, et al. (2013) Reactivity of polymeric proanthocyanidins toward salivary proteins and their contribution to young red wine astringency. J Agric Food Chem 61(4): 939-946.
-
Baxter N, Lilley T, Haslam E, Williamson M (1997) Multiple interactions between polyphenols and a salivary proline-rich protein repeat in complexation and precipitation. Biochemistry 36(18): 5566-5577.
-
Gawel R (1998) Red wine astringency: a review. Australian Journal of Grape and Wine Research 4(2): 74- 95.
-
Bacon J, Rhodes M (1998) Development of a competition assay for the evaluation of the binding of human parotid salivary proteins to dietary complex phenols and tannins using a peroxidase-labeled tannin. J Agric Food Chem 46(12): 5083-5088.
-
Jöbstl E, O Connel J, Fairclough J, Williamson M (2004) Molecular model for astringency produced by polyphenol-protein interactions. Biomacromolecules 5(3): 942-949.
-
Obreque Slier E, Peña Neira A, López Solís R (2012) Interactions of enological tannins with the protein fraction of saliva and astringency perception are affected by pH. International Journal of Food Science and Technology 45(1): 88-93.
-
Yang S, Zhang Y, Zhang X, Xu J (2007) The influence of pH on a hydrogen-bonded assembly film. Soft Matter 3: 463-469.
-
Patronov A, Salamanova E, Dimitrov I, Flower DR, Doytchinova I (2014) Histidine hydrogen bonding in MHC at pH 5 and pH 7 modeled by molecular docking and molecular dynamics simulations. Curr Comput Aided Drug Des 10(1): 41-49.
-
Thomas CJ, Lawless HT (1995) Astringent subqualities in acids. Chemical Senses 20(6): 593-600.
-
Ferrer Gallego R, Hernández Hierro JM, Brás NF, Vale N, Gomes P, et al. (2017) Interaction between wine phenolic acids and salivary proteins by saturation-transfer difference nuclear magnetic resonance spectroscopy (STD-NMR) and molecular dynamics simulations. J Agric Food Chem 65(31): 6434-6441.
-
García Estévez I, Pérez Gregorio R, Soares S, Mateus N, de Freitas V (2017) Oenological perspective of red wine astringency. OENO One 51(3): 237-249.
-
García Estévez I, Ramos Pineda AM, Escribano Bailón MT (2018) Interactions between wine phenolic compounds and human saliva in astringency perception. Food & Function 9(3): 1294-1309.
-
Grassl N, Kulak NA, Pichler G, Geyer PE, Jung J, et al. (2016) Ultra-deep and quantitative saliva proteome reveals dynamics of the oral microbiome. Genome Medicine 8: 44.
-
Messana I, Inzitari R, Fanali C, Cabras T, Castagnola M (2008) Facts and artifacts in proteomics of body fluids. What proteomics of saliva is telling us? J Sep Sci 31(11): 1948-1963.
-
López Solís R, Muñoz Romero D, Medel Marabolí M, Obreque Slier E (2020) Diverse interaction of commercial enological tannins with the protein fraction of saliva. Association with astringency. Food Structure 26: 100163.
-
Morales Bozo I, Urzúa Orellana B, Domínguez P, Aguilera S, López Solís RO (2006) Patterns and variability in electrophoretic polypeptide profiles of human saliva in a healthy population. J Physiol Biochem 62(3): 179-188.
-
Atkinson JC, Dawes C, Ericson T, Fox PC, Gandara BK, et al. (1993) Guidelines for saliva nomenclature and collection. Annals of the New York Academy of Sciences 694(1): 11-12.
-
López Solís R, Duarte Venegas C, Meza Candia M, Del Barrio Galán R, Peña Neira A, et al. (2017) Great diversity among commercial inactive dry-yeast based products. Food Chem 219: 282-289.
-
Durham JP, López Solís RO (1979) Effect of sodium hydroxyde upon protein binding to filter paper disks and its significance in the determination of protein in colorimetric and radioisotopic assays. Anal Biochem100(1): 98-99.
-
Gawel R, Oberholster A, Francis IL (2000) A mouth-feel wheel: terminology for communicating the mouth-feel properties of red wine. Australian Journal of Grape and Wine Research 6(3): 203-207.
-
Obreque Slier E, Peña Neira A, López Solís R (2010b) Enhancement of both salivary protein-enological tannin interactions and astringency perception by ethanol. J Agric Food Chem 58(6): 3729-3735.
-
Vazallo Valleumbrocio G, Medel Marabolí M, Peña Neira A, López Solís R, Obreque Slier E (2017) Commercial enological tannins: Characterization and their relative impact on the phenolic and sensory composition of Carménère wine during bottle aging. LWT-Food Science and Technology 83: 172-183.
-
Charlton AJ, Baxter NJ, Lilley TH, Haslam E, McDonald CJ, et al. (1996) Tannin interactions with a full-length human salivary proline-rich protein display a stronger affinity than with single proline-rich repeats. FEBS Letters 382(3): 289-292.
-
Soares S, Vitorino R, Osorio H, Fernandes A, Venancio A, et al. (2011) Reactivity of human salivary proteins families toward food polyphenols. J Agric Food Chem 59: 5535-5547.
-
Huang R, Xu C (2020) An overview of the perception and mitigation of astringency associated with phenolic compounds. Compr Rev Food Sci Food Saf 20(1): 1036- 1074.
-
Ferrer Gallego R, Hernández Hierro JM, Rivas Gonzalo JC, Escribano Bailón MT (2014) Sensory evaluation of bitterness and astringency sub-qualities of wine phenolic compounds: synergistic effect and modulation by aromas. Food Research International 62: 1100-1107.
- The Role of Podocyte Apoptosis and the Involvement of SIRT1 in Diabetic Nephropathy
- Dealcoholization of Beer by Osmotic Distillation for the Beverage Industry
- Biopolymer-Based Edible Packaging- Biomaterials, Methods, and Applications in Food Industry: An Updated Review
- Influence of Bioprocessing Methods on 'China Rice' (Gawal R1), and Soyabean Supplementation on the Quality of Complementary Food
- Cassava (Manihot esculenta) Varietal Growth, Yield and Cyanide Content Performance in Three Sites in the South- Eastern Semi Arid Regions of Kenya
- Food Waste Treatment, Recycling, Management and Production of Value-Products-An Update on Methodologies and Current Trends