Resurrection of the Fern "Cheilanthes albomarginata Clarke" Involves Utilization of Trehalose as Energy Source and Accumulation of Protective Stress Metabolites
<p>Most plants are unable to survive desiccation to an air-dried state, but a small group of plants known as resurrection plants can tolerate extreme desiccation, and regain normal function after rehydration. Cheilanthes albomarginata is one such resurrection fern, which has been analyzed in this study for metabolite changes during resurrection. In this study, we have tested C. albomarginata fronds from ten different geographical regions of Himachal Pradesh for the presence of trehalose, proline, and glycine betaine. The profile of trehalose sugar during resurrection was investigated using chromatographic techniques and enzymatic assays. Analysis of extracted sugars by High performance liquid chromatography (HPLC) showed that trehalose levels were highest in dehydrated stage of C. albomarginata (46 %), which reduced to 28% upon rehydration. Fourier Transform InfraRed spectroscopy (FTIR) analysis of intracellular sugars in crude extracts of C. albomarginata indicated the closest match of the sugar in the samples with trehalose rather than sucrose. Quantitative estimation of extracted trehalose sugar of C. lbomarginata using crude trehalase further confirmed the utilization of trehalose during resurrection. Thus, trehalose appears to be the major sugar accumulating during dehydrated condition, with an active role in resurrection of C. albomarginata. Proline and Glycine betaine were observed to accumulate during dehydration stage in C. albomarginata, and remained largely unaltered during rehydrated state, indicating their protective role during resurrection. </p>
Introduction
Elevation in the emissions of greenhouse gases, increased world population and worldwide climate change are some of the major key issues from the last few decades [1, 2, 3]. The ultimate result of these issues is global water shortage which now is one of the another major challenge, affecting livelihood and agriculture worldwide to larger extent [4]. Due to the global water shortage, sufficient amount of water is not available to the every corner of world and as a result, drought has become a major threat from the last few decades to agriculture globally as water scarcity contributes to the highest percentage death rate of many important crops. To feed every person in this planet, the requirements for valuable crops with high productivity are needed. But during heavy drought, most of plants do not survive and dies out. Exceptionally, a few plants known as resurrection plants, can survive in these extreme conditions [5]. Thus, from the few last decades, lots of efforts have been made by many researchers to understand the physiology, development and life cycle of these plants at cellular, biochemical and molecular level. Resurrection plants are characterized by the ability to revive or becoming alive after seeming to be dead. Upon dehydration, resurrection plants shrivel up and fold their leaves, until water is available [6, 7, 8]. Thus, it is important to understand the mechanism that how these plants adapt to the water stress and how they survive during the extreme drought. Since metabolites play a key role during drought stress and revival upon rehydration [9]. Therefore, analysis of metabolomics of these plants can help the researchers to find out the mysteries behind the survival in the extreme conditions. Cheilanthes albomarginata, a resurrection fern undergo its dehydration and rehydration cyles according to the environmental conditions [10]. Thus vegetative desiccation tolerance in this fern is seasonally regulated. The leaves of C. albomarginata culred up during summer season and revived again when water is available_. The RWC values in this fern showed that fern is able to survive even in 8% of remaining water and can also able to recover the chlorophyll content and metabolic activity at full rehydration. Many physiological factors such as light, temperature (data not published), water and developmental stage appear to have an enhancement effect on the resurrection activity of _C. albomarginata fronds [10]. In this study, an effort has been made to understand the metabolomics involved during the resurrection using various techniques and assays. The samples of C. albomarginata were collected and analysed during resurrection. The fresh stage of sample is considered as primary control and purified standards as secondary control for comparing and analyzing the metabolic changes in dry and rehydrated stages. Finally, the correlation statistics is applied on the results obtained in each analysis with relative water content, time for complete resurrection and altitude. To our knowledge, this is the first study to provide insight into the resurrection activity in C. albomarginata and its metabolic characterization.
Materials and Methods
Collection and Identification of Fern Samples
The fronds of C_. albomarginata_ fern were collected during rainy season (July-August) from Solan and adjoining areas, Himachal Pradesh, India [10]. The fronds were first washed with 1% (v/v) H2O2 and then with distilled water and used immediately upon collection. The remaining plants were stored at -80 °C. All samples were identified from herbarium of Punjab University, Chandigarh India, University of Horticulture and Forestry, Nauni, Solan, H.P., and other resources [11].
Thin Layer Chromatography (TLC) Analysis of Sugars during Resurrection of C. albomarginata
Ethanolic crude extracts of powdered fronds of C. albomarginata were prepared, weighed and re-suspended in ethanol to a concentration of 1 mg/ mL, and used for TLC analysis of sugars. For the analysis of sugars, standards of trehalose (Sigma-Aldrich, USA) and glucose (Himedia Labs, Mumbai) were spotted along with ethanolic extracts (1 mg/ mL) of fronds of fresh, dry and rehydrated stages of C. albomarginata onto pre-cast silica gel TLC plates (Merck, Millipore India Pvt. Ltd., Bangalore) and developed with a solvent system containing butanol : pyridine : water (in a ratio of 15 : 30 : 20, v/v) as the mobile phase. After drying at 50 °C for 10 min in an hot air oven, and the spots were detected by spraying with potassium permanganate solution.
Analysis of Sugars in the Crude Extracts of C. albomarginata by FTIR
For FTIR analysis, dry ethanolic crude extracts of fresh, dry and rehydrated stages of C. albomarginata fronds were used and analysed against the standards of sucrose and trehalose sugars. FTIR spectra were recorded using an IR-spectrometer (Carry 650 FTIR, Agilent Technologies) and mercury cadmium telluride (MCT) detector.
Qualitative Analysis of Sugars by HPLC
HPLC analysis was carried out using a gradient modular pump, Agilent 1200 series injector fitted with a 25 µL loop. For detection, differential refractometer (RI) detector was used and chemstation software (version 6.0) was used. An L-Amine 210 (YMC Co. ltd, Japan) column of 50 x 50 cm was used along with 45 % acetonitrile/water (v/v) with 0.1 M ammonium acetate as the mobile phase at a flow rate of 1.0 mL /min. The run time was 60 min. Qualitative determination was done by comparison of sugars extracted from fresh, dry and rehydrated stages of samples of C. albomarginata fronds (as described under TLC section) against the standard mixtures of glucose, sucrose and trehalose sugars.
Extraction of Trehalose from the Fronds of C. albomarginata by Hot Ethanol Method
Trehalose was extracted using the modified hot ethanol method of Vázquez‐Ortíz, et al. [12]. The extracts obtained were re-suspended in 100 µL sterile distilled water and stored at -20 ⁰C until use. Trehalase Enzyme Assay: Preparation of crude trehalase enzyme from Sacchromyces cereviseae. The wild type strain of Sacchromyces cereviseae was used to prepare crude trehalase according to the method given by Rossouw, et al. [13] and stored at 4°C until use. The protein content in the crude trehalase preparation was determined by the Bradford method [14] using bovine serum albumin (BSA) as a standard. Trehalase Assay: The trehalose extracted from C. albomarginata (as described above) or the commercial trehalose [2.5 µM (limiting substrate concentration) and 10 µM (saturating substrate concentration)] was incubated with crude trehalase enzyme in assay buffer. The reaction mixture was incubated at 30°C for 1 h and the reaction was stopped by incubating at 90°C for 2 min in a dry bath. The assay mixture was subsequently used for estimating the amount of glucose released by 3,5- Dinitrosalicylic acid (DNS) method [15]. The trehalase assay was performed in two sets: limiting amount of trehalase enzyme (340 µg total protein) and substrate (50 µL of extracted trehalose); saturating amount of enzyme (850 µg) and substrate (100 µL) along with negative controls.
Estimation of Proline during Resurrection of C. albomarginata Fronds
The proline content was quantified according to the method of Bates, et al. [16]. The assay was performed with standard proline (50 µmol) and a standard curve was generated. The amount of proline in the test samples was calculated from the standard curve.
Quantitation of Glycine-betaine (GB) during Resurrection of C. albomarginata Fronds
The estimation of GB was done according to the method of Greive and Grattan [17]. The assay was also performed with standards of GB (50-200 mg/mL) and the standard curve was generated. The amount of GB in the samples of C. albomarginata was determined from the standard curve.
Correlation Analysis of Experimental Data
To study the correlation of resurrection phenomenon with the different experimental factors, the results were analyzed using Microsoft Excel and Statistical Packages for Social Sciences (SPSS 20) statistical analysis software (IBM Corporation, US, version 20) using two replicates of trehalose content, proline content and GB content of C. albomarginata resurrection samples. Paired correlation analysis was performed between relative water content (RWC), resurrection time and altitude during different stages of resurrection (fresh, dry and rehydrated stage). Statistical correlative analysis was performed by bivariate correlation using "Pearson correlation coefficients" for two-tailed test of significance with least significant difference (LSD) less than p value of 0.05.
Results
Qualitative Analysis of Sugars Present in Different Stages of Resurrection by FTIR Spectroscopy
C. albomarginata is widely distributed in Himachal Pradesh. In our previous study, we reported the resurrection activity of C. albomarginata collected from ten different districts of Himachal Pradesh, India. These locations vary in their climatic conditions and altitude (372 m-2455 m) [10]. Trehalose and sucrose are the major sugars that accumulate during desiccation in plants. Therefore, the FTIR spectra of ethanolic extracts of C. albomarginata were compared with those of standard trehalose and sucrose. The position of the major IR peaks in the spectra of fresh, dry and rehydrated stage samples seemed to match with each other (Figure 1 A, B and C). The first pre- dominant IR peak in all the 3 spectra (Figure 1 A-C) was found as a broad peak centered around 3300 cm-1, which corresponds O-H or N-H group. The next five IR peaks were positioned around 2925 cm-1 (corresponding to C- H), 2854 cm-1 (corresponding to C-H), 1700 cm-1 (corresponding to C=O or C-C), 1600 cm-1 (corresponding to C=C) and 1020 cm-1 (corresponding to C-N or C-X) (Figure 1 A, B and C). Some of the peaks were observed as small peaks in the IR spectra, indicating the presence of some other interacting functional groups in ethanolic extracts (Figure 1A). The ethanolic extracts of different stages of resurrection showed no IR peaks at 2995 cm-1, which was only present in the sucrose standard (Figure 1 A-E).
Comparison of the FTIR profiles of the resurrection samples with that of trehalose revealed similarities at positions of 3250 cm-1, 2100 cm,-1and 1350 cm-1 respectively (Figure 1 A-D). These results indicate the possible presence of trehalose, and absence of sucrose in the extracts of fresh, dry and rehydrated stages of fronds of C.albomarginata.
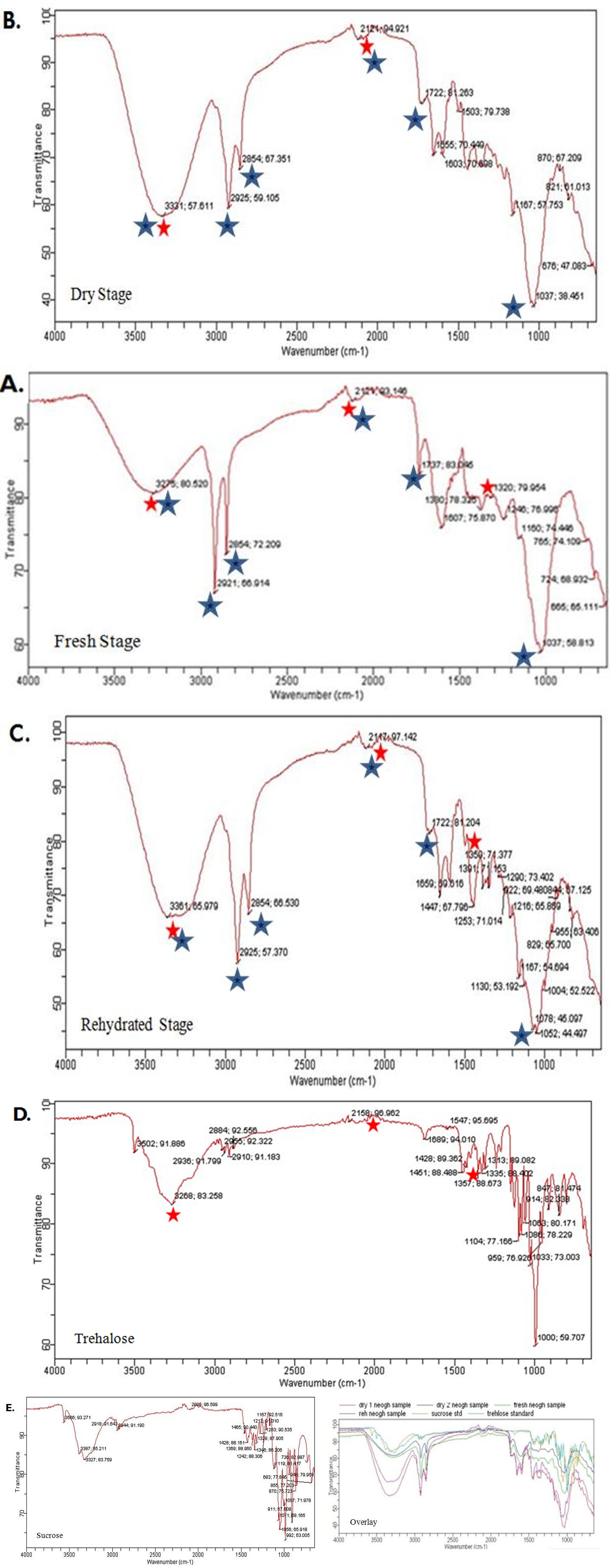
Characterization of Sugars in Different Stages of Resurrection in C. albomarginata by TLC
To understand the role of sugars in resurrection, trehalose was analyzed in ethanolic extracts during fresh, dry and rehydration stages. In order to detect variation in trehalose content during different stages of resurrection, TLC analysis of the ethanolic extracts of the three stages of resurrection were performed along with trehalose as a standard. TLC analysis was done and developed using KMnO4. As shown in Figure 2, a prominent spot corresponding to the mobility of the trehalose was observed in fresh and dry stages, but not in rehydrated stage of resurrection in C. albomarginata ethanolic extracts. Moreover, the intensity of the spot (Rf = 0.84) corresponding to mobility of the trehalose was almost double in the dry stage (Rf = 0.87) as compared to that of fresh stage (Rf = 0.86).
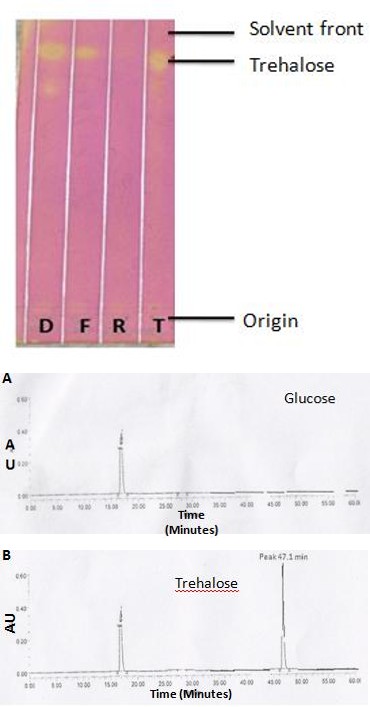
corresponding to trehalose in the rehydrated stage (Figure 2). Thin layer chromatography analysis of crude ethanolic extracts of C. albomarginata samples during fresh (F), dry (D) and rehydrated (R) stages, developed with KMnO4. Yellow spots of sugars were visible in a pink background. The trehalose (T) loaded as reference is indicated.
Qualitative Analysis of Trehalose during Resurrection of C. albomarginata by HPLC
The analysis of sugars profile by TLC favor the possibility that trehalose is accumulated by C. albomarginata during desiccation/dry condition, and metabolized during rehydration. However, the possible role for other stress sugars like sucrose is still not eliminated. Therefore, a more sensitive analysis was performed by HPLC using trehalose, glucose and sucrose standards. Comparison of the HPLC profiles of ethanolic extracts of C_. albomarginata_ fronds from fresh, dry and rehydrated stages with the standards revealed the presence of trehalose in all the three stages. On the other hand, glucose was detected only in fresh and rehydrated stages (Figure 3; Table 1). Several other peaks were present in the ethanolic extracts of C_. albomarginata_, whose identity could not be established (Figure 3 D-F). Quantitative analysis of trehalose content from chromatogram showed that trehalose begins to accumulate during fresh stage (43.6 %); accumulates further at dry stage (46 %); and its level declines during rehydration stage (28 %), possibly due to its utilization as an energy source. Moreover, it was evident from the HPLC chromatograms that sucrose was undetectable during all stages, indicating the role of trehalose as desiccation protectant molecule (Figure 3; Table 1). Consistent with the possible metabolism of trehalose to release glucose, a slight increase in glucose content was observed during the rehydration stage (1.87 %).
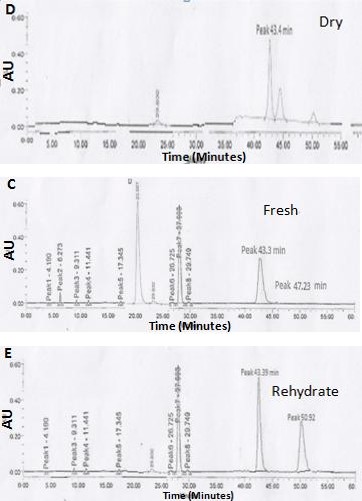
| S. No. | Metabolites | Retention time (min) | (%) area | ||
| S. No. | Metabolites | Retention time (min) | Fresh | Dry | Rehydrate |
| 1 | Glucose | 17 | 1.3 | -* | 1.87 |
| 2 | Trehalose | 43 | 43.6 | 46 | 28 |
Table 1: List of Sugars, their retention time and percentage area analyzed by HPLC during fresh, dry and rehydrated stages of C.
Quantitation of Trehalose Content during different Stages of Resurrection of C. albomarginata by Trehalase Assay
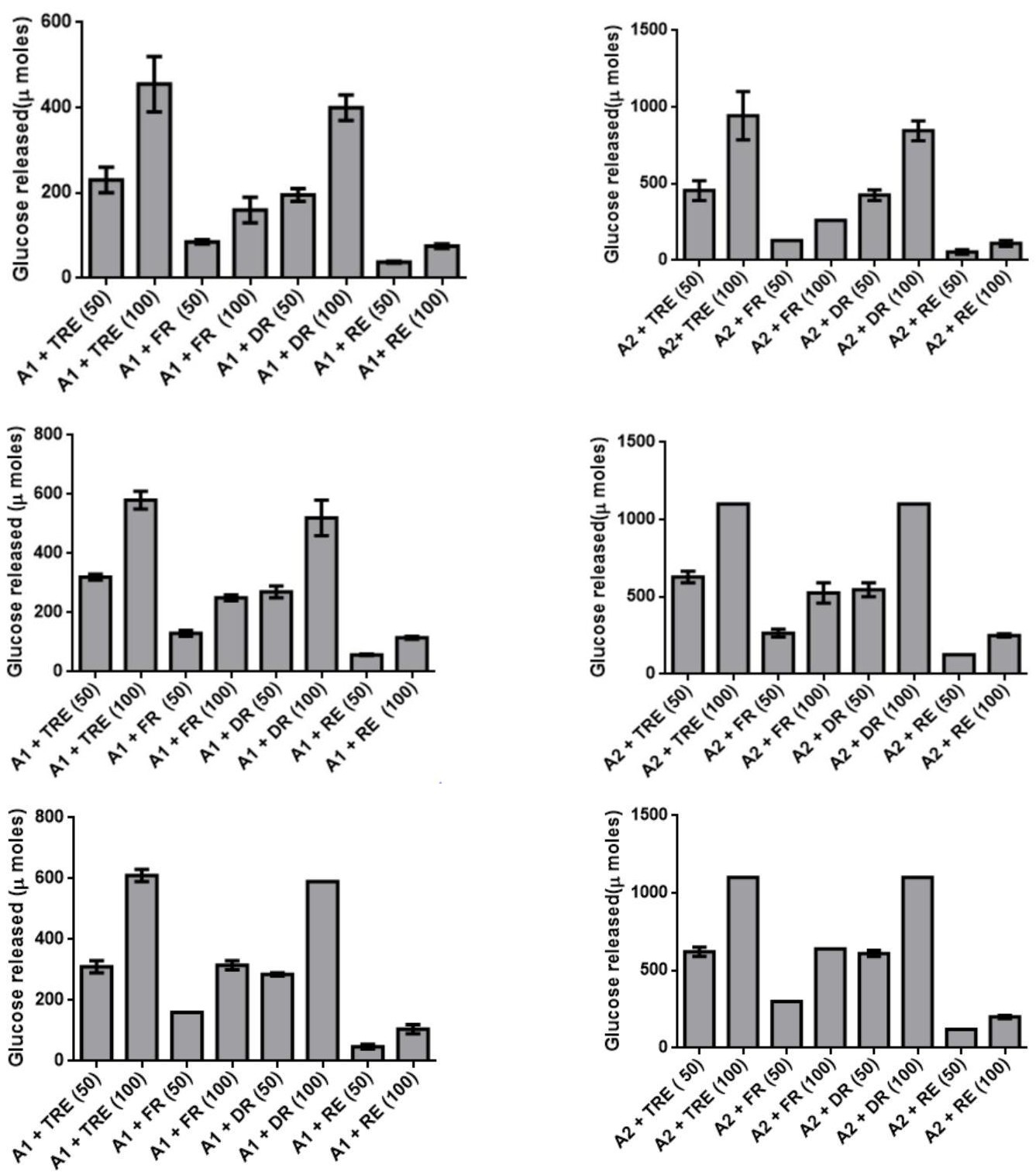
The changes in the trehalose content during desiccation and resurrection indicated its role as desiccation protectant in C_. albomarginata.To quantitate and further validate the amount of trehalose during different stages of resurrection, an enzymatic assay was designed. The conversion of trehalose extracted from different stages of resurrection into glucose was assayed by using a crude preparation of trehalase enzyme from the yeast _Saccharomyces cerevisiae. The specificity of the assay was confirmed by using appropriate controls (enzyme alone and extracted trehalose alone). The commercial trehalose was used as a positive control for the enzyme assay. The key results from these assays are summarized below: 1) The presence of trehalase in crude enzyme was confirmed by the dose dependent release of glucose upon incubation with purified trehalose (5µM and 10 µM) in all the experiments [A1/A2 + 50/100 (TRE) data in Figure 4 A - J and a – j]. 2) The release of glucose was specific to trehalase, since no activity was observed without trehalase. 3) A dose dependent increase in the release of glucose was observed with increasing amounts of enzyme [340 µg and 850 µg; (Figure 4 A-J and a-j)] as well as with increasing amounts of trehalose (50 and 100 µL) extracted from fresh, dry and rehydrated stages of C. albomarginata fronds (Figure 4 A-J and a-j). 4) A uniform pattern of trehalose utilization, i.e., amount of glucose released during the three stages of resurrection (fresh, dry and rehydrated)was observed in all the 10 samples (Figure 4 A-J and a-j). It was observed that during rehydration stage, the amount of glucose released was reduced to approximately half as compared to dry stage sample of C. albomarginata. Similar results were observed for all the 10 samples of C. albomarginata (Figure 4 A-J and a-j). Together, these results confirm the role of trehalose in the resurrection activity of C. albomarginata and its utilization during rehydration. 5) The amount of glucose released from trehalosein dry stage of all the 10 samples of C. albomarginata was almost double the amount when compared to fresh stage of respective sample (Figure 4). These results indicate the accumulation of trehalose during dry stage, a feature observed in many resurrection plants during desiccation. Thus, qualitative (TLC, FTIR and HPLC) and quantitative (trehalase assays) experiments strongly indicate a role for trehalose in resurrection of C. albomarginata fronds.
A. Bajhol a. A1 - 340 µg crude trehalase A2 – 850 µg crude trehalase
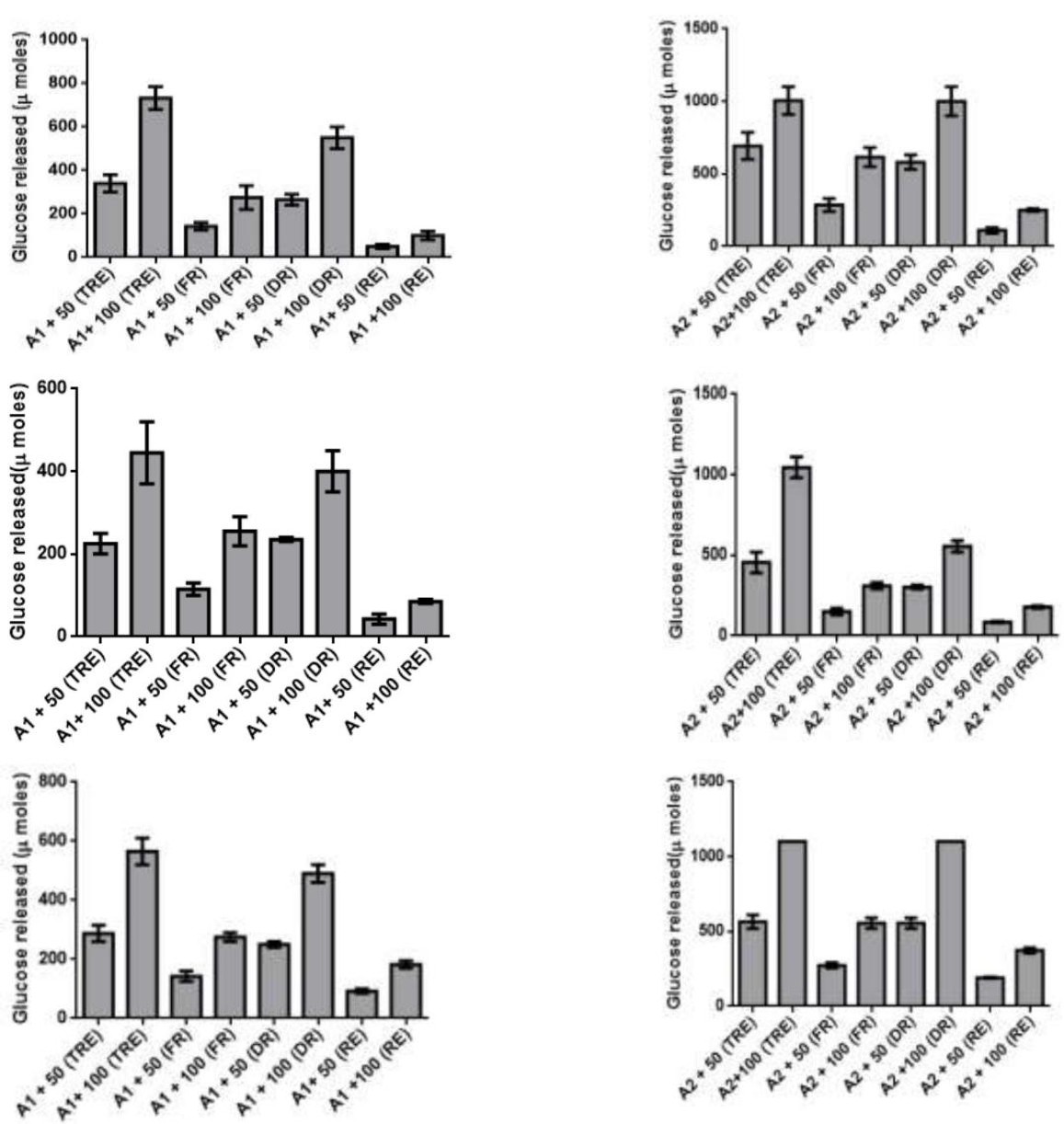
B. Chail b.
C. Dharampur c.
D. Kasauli d. A1- 340 µg crude trehalase A2– 850 µg crude trehalase
E. Mandi e.
F. Nalagarh f.
G. Shimla g. A1- 340 µg crude trehalase A2- 850 µg crude trehalase
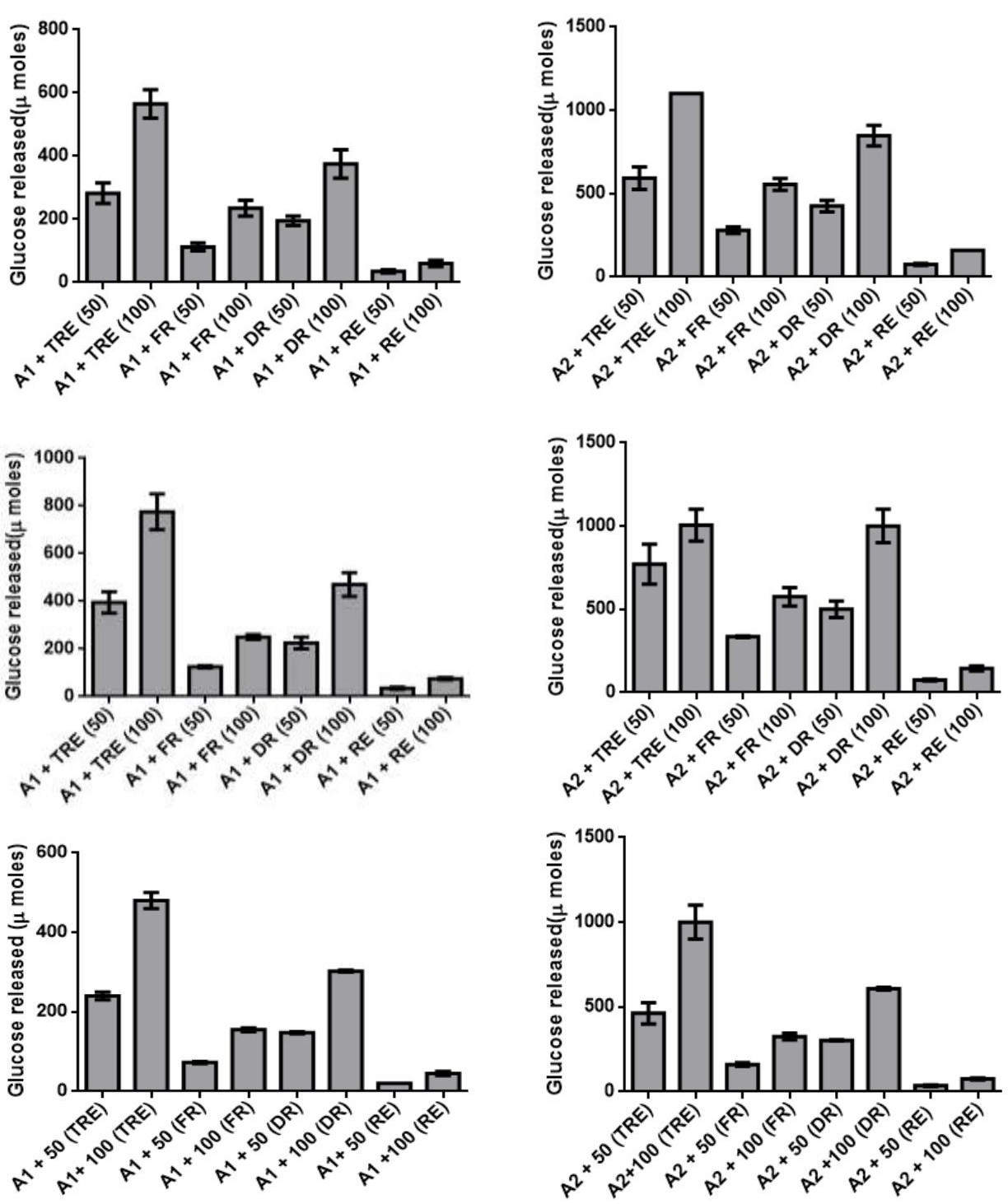
H. Shoghi h.
I. Sirmour i.
J. Subathu j. A1- 340 µg crude trehalase A2– 850 µg crude trehalase
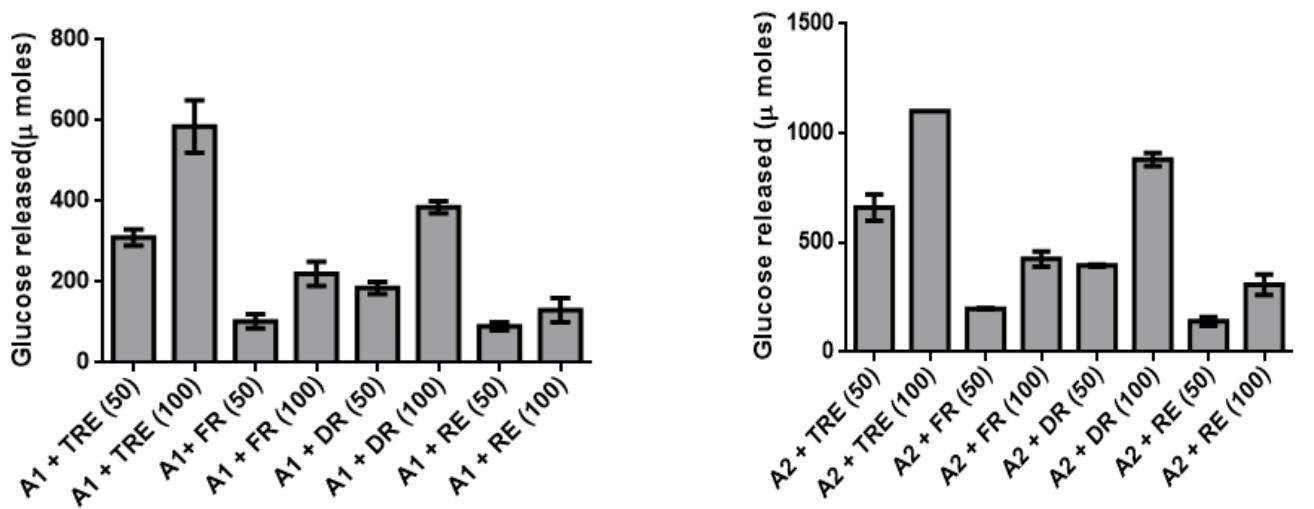
Quantitative Analysis of Proline Content during Resurrection of C. albomarginata
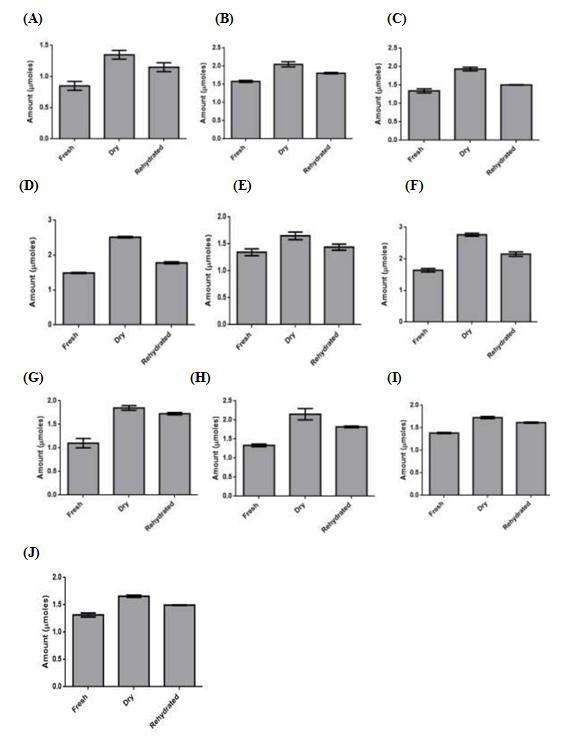
The amino acid proline has been found to accumulate in the tissues of several plants during desiccation [18]. To study the role of proline in the resurrection of C. albomarginata, proline content was measured in all the 10 samples during different stages of resurrection as described above. Proline was detected in the fresh stage of C. albomarginata fronds of all the 10 locations, ranging from 1.1 µmol/g in Shimla to 1.85 µmol/g in Bajhol sample (Figure 5A-J). An increase in proline content (59 – 137 %) was observed in the dry stage, respectively of C. albomarginata fronds of all the ten locations relative to the fresh stage (Figure 5A-J; fresh versus dry). These results indicate that proline accumulates in response to desiccation stress. Interestingly, amount of proline in rehydrated stages of all the samples locations of C. albomarginata was only marginally reduced (62-156 %) relative to the proline content in their respective dry stages (Figure 5A-J; dry versus rehydrated). These results suggest that proline accumulates during dry stage and its levels are maintained during rehydration, indicating its role as an osmoprotectant during rehydration and desiccation. The pattern of proline accumulation during different stages of resurrection of C. albomarginata collected from ten different locations was consistent with each other (Figure 5A-J).
Quantitative analysis of Glycine Betaine (GB) Content during Resurrection of C. albomarginata
Glycine betaine is known to accumulate as osmolyte in several plant species during desiccation [19]. Extreme dryness or desiccation is one form of stress that can induce GB production and accumulation, and in some cases, accumulation is considered to be an adaptive response [19]. GB accumulation was analysed during resurrection of C. albomarginata. The amount of GB in different stages of resurrection of C. albomarginata samples collected from 10 different locations was determined.
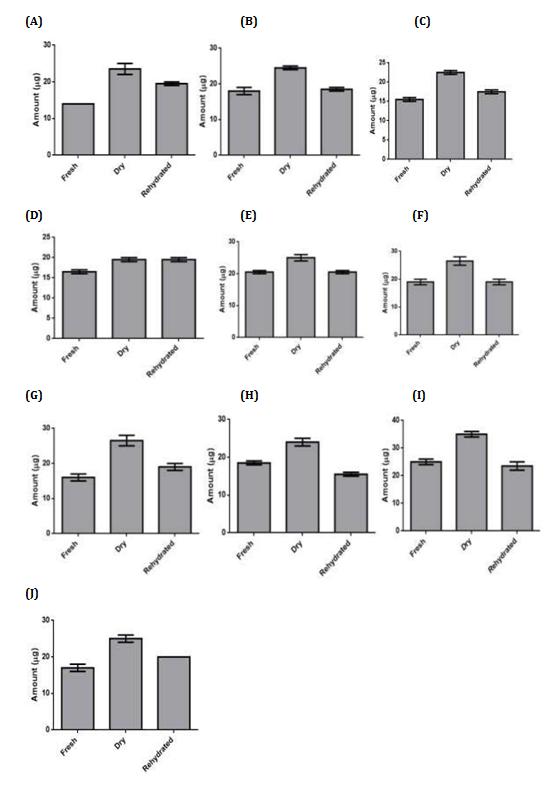
Glycine betaine could be detected in all the three stages of resurrection in C. albomarginata collected from the ten locations (Figure 6, A-J). The amount of GB during the fresh stage was lowest (14 µg) in the Bajhol sample, while highest in (25 µg) in Sirmour sample (Figure 6). Upon drying, a significant increase in the levels of GB was observed for all the ten samples, which ranges from 121 to 192 % relative to the levels in the fresh stage (Figure
6). These results indicate that GB accumulates in response to dryness in the fronds of C. albomarginata. Similar to the proline, most of the GB (80 %) content present in the dry stage was retained during rehydration stage of C. albomarginata collected from the ten different locations (Figure 6). These results suggest that GB accumulates during dryness/desiccation and retained during rehydration of C. albomarginata fronds. Together both proline and GB appear to serve as osmoprotectant during resurrection of C. albomarginata.
Correlation Analysis of Physical and Biological Factors with Resurrection of C. albomarginata
To study the relationship between resurrection phenomenon and the physiological parameter involved, paired correlation analysis was performed between accumulation of metabolites such as trehalose, proline and glycine betaine, and physiological factors such as relative water content, time taken for resurrection, temperature and altitude. Three types of correlations were observed: positive, negative or no correlation (Table 2). The trehalose content in fresh stage of C. albomarginata had no correlation with either of the physiological parameters tested (Table 2). On the other hand, trehalose content in dry stage showed inverse correlation with RWC, and the altitude of the location. Similarly, during rehydration condition, trehalose content negatively correlated with RWC (the capacity to obtain full turgidity).
| S. No. | Parameters | RWC | Resurrection time | Temp | Altitude |
| A. | Trehalose content | ||||
|---|---|---|---|---|---|
| 1 | Fresh stage | NS* | NS | NS | NS |
| 2 | Dry stage | -S* | NS | NS | -S |
| 3 | Rehydrated stage | -S | NS | NS | NS |
| B. | Proline content | ||||
| 1 | Fresh stage | NS | NS | NS | NS |
| 2 | Dry stage | +S* | NS | NS | NS |
| 3 | Rehydrated stage | NS | -S | NS | NS |
| C. | Glycine-Betaine content | ||||
| 1 | Fresh stage | NS | NS | NS | NS |
| 2 | Dry stage | NS | NS | NS | NS |
| 3 | Rehydrated stage | NS | NS | NS | NS |
Table 2: Correlation analysis between various physiological parameters and metabolite accumulation during different Table 2: Corr
Table 2: Correlation analysis between various physiological parameters and metabolite accumulation during different Table 2: Correlation analysis between various physiological parameters and metabolite accumulation during different stages of resurrection of C. albomarginata. *(NS): Not significant, i.e., no significant correlation. *(+S): Positive Significant value, i.e., positive correlation. *(-S): Negative Significant value, i.e., negative correlation.
The correlation analysis of stress amino acid proline did not indicate any correlation with any of the parameter during fresh stage, whereas during dry stage, accumulation of proline showed a direct correlation with RWC (Table 3). During rehydration stage, proline content showed a negative correlation with resurrection time (Table 3). GB content did not show any correlation during fresh stage, dry and rehydrated stages with any of the parameters studied (Table 3).
| Metabolite | Stage of resurrection | Qualitative analysis* | ||
|---|---|---|---|---|
| Presence/ Absence | Accumulation /utilization | |||
| 1 | Trehalose | Fresh | (+) | (+++) |
| 1 | Trehalose | Dry | (+) | (+++) |
| 1 | Trehalose | Rehydrated | (+) | (--) |
| 2 | Proline | Fresh | (+) | (+++) |
| 2 | Proline | Dry | (+) | (+++) |
| 2 | Proline | Rehydrated | (+) | (+++) |
| 3 | Glycine Betaine | Fresh | (+) | (+++) |
| 3 | Glycine Betaine | Dry | (+) | (+++) |
| 3 | Glycine Betaine | Rehydrated | (+) | (+++) |
Table 3: Qualitative analysis of metabolites during resurrection activity in C. albomarginata.
Discussion
Resurrection is the ability of an organism to survive the loss of most ~ 95% of its cellular water for extended periods and to recover full metabolic competence upon rehydration. The plant kingdom has many unique groups of plant species, ranging from lower to higher groups. Among others, only a few can tolerate extreme water loss (desiccation) of down to 5% of cellular water content for longer periods without dying, commonly referred to as resurrection plants [5, 20, 21, 22, 23, 24].
Desiccation tolerance (DT) in resurrection plants is a multi-genic and multi-factorial phenomenon and is associated with prevention of damage caused by oxidation. Resurrection plants produce a high diversity of metabolites for defense against desiccation and are required for growth and developmental processes during lethal conditions e.g. osmoprotectants, such as sugars (trehalose, sucrose and fructan), amino acids (tryptophan and proline) and ammonium compounds (polyamines and glycinebetaine) [25]. These molecules accumulate in these plants under lethal conditions arise due desiccation as adaptive mechanism and provide stress tolerance. Thus, metabolites play a key role during desiccation stress and revival upon rehydration [9]. Thus, to characterize the metabolites essential for resurrection in C. albomarginata, quantitative and qualitative analysis of various metabolites such as trehalose, proline and glycine betaine during dehydration/desiccation and rehydration were performed.
In this study, techniques like TLC, FTIR and HPLC were used as the major techniques for the characterization of sugars, while enzymatic and colorimetric assays were used for the quantitative estimation of trehalose, proline and glycine betaine during resurrection.
In TLC analysis, the appearance and disappearance of spot corresponding to trehalose was observed in the crude dehydrated and rehydrated samples, respectively. The disappearance of trehalose during rehydration might indicate its utilization by the plant as an energy source during rehydration process. Quantitative estimation of extracted sugars by HPLC analysis from fresh, dehydrated and rehydrated stages of C. albomarginata showed that trehalose levels were highest in dehydrated stage (46 %), which reduced to 28% upon rehydration. Thus, HPLC data further support the TLC results. FTIR analysis of intracellular sugars in crude extracts of C. albomarginata during fresh, dehydrated and rehydration stages indicated the closest match of the sugar in these samples with trehalose standard rather than sucrose standard. Quantitative estimation of extracted trehalose sugar from the fronds of fresh, dry and rehydrated stages of C. albomarginata was done using trehalase assay. Interestingly, the pattern of trehalose accumulation was consistent with results obtained from TLC and HPLC analysis, in all C. albomarginata samples collected from ten different locations (Fig 4). Thus, trehalose appears to be the major sugar accumulating during dehydrated condition, and might have a protective role in C. albomarginata during desiccation.
Proline plays multiple roles as stress protectant, viz, stabilizing enzymes, membranes, proteins and free radical scavengers. Besides acting as an excellent osmolyte, it also acts as a metal chelator, an antioxidative defense molecule and a signaling molecule. Proline accumulation is a common phenomenon observed in response to environmental stress in many organisms including plants [26]. In plants, intracellular proline levels have been found to increase by > 100-fold during stress [27]. Proline accumulation in plants occurs during exposure to various stresses, including salt [28], drought [29], UV radiation [30], heavy metal ions [31], pathogens [32], and oxidative stress [33].
The level of proline accumulation during dehydration stage varied significantly amongst the 10 samples of C. albomarginata (Figure 5). The highest proline accumulation was observed in Nalagarh sample (2.7 µmol), followed by Kasauli sample (2.5 µmol) and Shoghi sample (2.1 µmol), respectively. The lowest proline accumulation was observed in Bajhol sample (1.35 µmol).
GB protects cells from stresses by maintaining an osmotic balance with the surrounding environment [34] and by stabilizing the quaternary structures of complex proteins. In most plants, Glycine betaine acts as an osmolyte and provides protection to the plant by scavenging of reactive oxygen species (ROS) to restore redox metabolism, and mediates preservation of cellular turgor by restitution of osmotic balance.
In this study, GB content was estimated in all the collected samples of C. albomarginata during fresh, dehydrated and rehydrated stages of resurrection, to understand the role of GB. Similar to proline, GB accumulated during dehydration stage in all the 10 samples of C. albomarginata, and remained largely unaltered during rehydrated state, indicating a protective role for GB during resurrection (Figure 6). The level of GB accumulated during dry stage varied significantly amongst the 10 samples of C. albomarginata; the highest GB accumulation was observed in Sirmour sample (35
µg), followed by Nalagarh sample (26.5 µg) and Shimla sample (26.5 µg), respectively. The lowest GB accumulation was observed in Kasauli sample (19.5 µg).
Correlative analysis between trehalose accumulation and the RWC indicated that in fresh stage, there was no significant correlation of the trehalose content with RWC content and resurrection time, while in dehydrated stage, the accumulation of trehalose showed inverse relation with RWC. In addition, accumulation of trehalose sugar during dehydrated stage had a significant inverse correlation with altitude of the place of collection of C. albomarginata fronds. No such reports are available in literature for comparison. The correlation analysis of proline content did not show any correlation with any of the factors during fresh stage, whereas during dehydrated stage, accumulation of proline showed a positive correlation with RWC, i.e., lesser the water content in the cell or higher the RWC, more is the synthesis and accumulation of enzyme/membrane/protein stabilizers like proline during dehydrated conditions. During rehydration stage of resurrection of C. albomarginata, proline content showed an inverse correlation with resurrection time; hence more the proline content during the rehydrated stage, faster is the resurrection of the plant. The accumulation of GB did not show any correlation with RWC, resurrection time and altitude in either of the three stages of resurrection. The GB has generalized protective role again oxidative stress in all the plants. Further studies are required to confirm its role during resurrection.
Conclusion
From the present study, it can be concluded that various metabolites accumulated during dehydrated stage of C. albomarginata in response to physiological conditions and protect the plant from the detrimental effects of the desiccation. In addition, during rehydration, few metabolites were used up (example, trehalose) for resurrection activity, whereas others were maintained as osmoprotectants (proline and GB). This study has revealed that the fern C. albomarginata is sensitive to dehydration stress, and relies on readily inducible protection to combat desiccation. A detailed characterization in future of the genomics and proteomics of resurrection phenomenon should help in better understanding of the adaptive mechanisms developed by resurrection plants and identify the role of metabolites involved in stress tolerance.
References
-
Kiriga WJ, Yu Q, Bill R (2016) Breeding and genetic engineering of drought-resistant crops. Int J Agr Crop Sci 9(1): 7-12.
-
Tilman D, Balzer C, Hill J, Befort BL (2011) Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci USA 108(50): 20260- 20264.
-
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, et al. (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol Manag 259(4): 660-684.
-
Vörösmarty CJ, Green P, Salisbury J, Lammers RB (2000) Global water resources: vulnerability from climate change and population growth. Science 289(5477): 284-288.
-
Challabathula D, Bartels D (2013) Desiccation tolerance in resurrection plants: new insights from transcriptome, proteome and metabolome analysis. Front Plant Sci 4: 482.
-
Lambers H, Chapin FS, Pons TL (2008) Photosynthesis. In: Plant physiological ecology Springer, New York, NY, pp: 11-99.
-
Farrant JM, Moore JP (2011) Programming desiccation-tolerance: from plants to seeds to resurrection plants. Curr Opin Plant Biol 14(3): 340- 345.
-
Dinakar C, Bartels D (2012) Light response, oxidative stress management and nucleic acid stability in closely related _Linderniaceae_ species differing in desiccation tolerance. Planta 236(2): 541–555.
-
Lamichhane S, Yde CC, Forssten S, Ouwehand AC, Saarinen M, et al. (2014) Impact of dietary polydextrose fiber on the human gut metabolome. J Agric Food Chem 62(40): 9944-9951.
-
Sharma A, Shruti K, Dev K, Sourirajan A (2015) A report on resurrection activity of the fern ‘_Cheilanthes_ _albomarginata clarke_’ from various regions of Himachal Pradesh, India. Int J Life Sci Biotechnol Pharma Res 5(4): 1-7.
-
Olsen Sue (2007) Encyclopedia of garden ferns. Timber Press.
-
Vázquez‐Ortíz FA, Valenzuela‐Soto EM (2004) HPLC Determination of Trehalose in _Selaginella lepidophylla_ Plants. J Liq Chromatogr Relat Technol 27(12): 1937- 1946.
-
Rossouw D, Heyns EH, Setati ME, Bosch S, Bauer FF (2013) Adjustment of trehalose metabolism in Wine _Saccharomyces cerevisiae_ strains to modify ethanol yields. Appl Environ Microbiol 79(17): 5197-5207.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1): 248-254. _15._ Miller GL (1959) Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal Chem 31(3): 426-428.
-
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39(1): 205-207.
-
Grieve CM, Grattan SR (1983) Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 70(2): 303-307.
-
Verbruggen, Nathalie, Christian H (2008) Proline accumulation in plants: a review. Amino acids 35(4): 753-759.
-
Ashraf M, Foolad MR (2007) Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ Exper Bot 59(2): 206-216.
-
Gaff DF (1977) Desiccation tolerant vascular plants of Southern Africa. Oecologia 31(1): 95-109.
-
Farrant JM, Brandt W, Lindsey GG (2007) An overview of mechanisms of desiccation tolerance in selected angiosperm resurrection plants. Plant Stress 1(1): 72-84.
-
Farrant JM, Cooper K, Nell H (2012) Desiccation tolerance. Plant stress physiology, Publisher: CAB International, pp: 238-265.
-
Gechev TS, Dinakar C, Benina M, Toneva V, Bartels D (2012) Molecular mechanisms of desiccation tolerance in resurrection plants. Cell Mol Life Sci 69(19): 3175-3186.
-
Gaff DF, Oliver M (2013) The evolution of desiccation tolerance in angiosperm plants: a rare yet common phenomenon. Funct Plant Biol 40(4): 315-328.
-
Rosenthal GA (1991) The biochemical basis for the deleterious effects of L-canavanine. Phytochemistry 30(4): 1055-1058.
-
Okuma E, Soeda K, Tada M, Murata Y (2000) Exogenous proline mitigates the inhibition of growth of _Nicotiana tabacum_ cultured cells under saline conditions. J Soil Sci Plant Nutr 46(1): 257-263.
-
Rentsch D, Hirner B, Schmelzer E, Frommer WB (1996) Salt stress-induced proline transporters and salt stress repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease-targeting mutant. Plant Cell 8(8): 1437-1446.
-
Thakur PS, Rai VK (1982) Dynamics of amino acid accutnulation in two differentially drought resistant _Zea mays_ cultivars in response to osmotic stress. Env Exp Bot 22(2): 221-226.
-
Munns R (2005) Genes and salt tolerance: bringing them together. New Phytol 167(3): 645-663.
-
Larcher WA (1981) Effects of low temperature stress and frost injury on plant productivity. Physiological processes limiting plant productivity 253-269.
-
Fougère F, Le Rudulier D, Streeter JG (1991) Effects of salt stress on amino acid, organic acid, and carbohydrate composition of roots, bacteroids, and cytosol of alfalfa (_Medicago sativa_ L.). Plant Physiol 96(4): 1228-1236.
-
Rajendra kumar CSV, Suryanarayana T, Reddy AR (1997) DNA helix destabilization by proline and betaine: possible role in the salinity tolerance process. FEBS Letter 410(2-3): 201-205.
-
Singh TN, Aspinall D, Paleg LG (1972) Proline accumulation and varietal adaptability to drought in barley: a potential metabolic measure of drought resistance. Nature New Biol 236(67): 188-190.
-
Robinson SP, Jones GP (1986) Accumulation of glycinebetaine in chloroplasts provides osmotic adjustment during salt stress. Funct Plant Biol 13(5): 659-668.
- An Efficient and Affordable Method for Isolating Bone Marrow- Derived Mesenchymal Stem Cells from Swiss Albino Mice
- Superposition of Cryo-EM and AlphaFold Predictions of Dengue Antigen-Antibody Complexes
- Jugular-Applied Coherent Low-Level Laser Therapy Enhances Systemic Mitochondrial Metabolic Function and Antioxidant Response
- Role of OMC32 Polypeptide in Acrosin-Mediated Exocytosis during the Bovine Sperm Acrosome Reaction
- Association of Galectin-3 but not Laminin in Tamoxifen-Induced Growth Suppression in Breast Cancer MCF-7 Cells
- Effect of Different Wavelengths of Light on the Rate of Photosynthesis