Preliminary Observations on the Ability to Adapt To Salinity Changes in Nursehound Scyliorhinus Stellaris (Linnaeus, 1758) and Small Spotted Catsharks Scyliorhinus Canicula (Linnaeus, 1758)
Nursehound Scyliorhinus stellaris and small spotted catsharks Scyliorhinus canicula are frequently hosted in public aquaria and their husbandry is improving. The reproduction of this species in captive conditions is not difficult and juveniles are easily maintained in laboratory or in thematic tanks showed to the public. Data on juveniles are easily acquired in order to understand different aspects of their natural life and physiology. In two stages, during 2006 and 2010, six S. stellaris and three S. canicula juveniles were tested at different salinity levels under controlled conditions in order to understand their responses in terms of respiration rate to this stress. In fact this is a benthic species living at constant water quality parameters at approximately 70-100 meters deep.
Introduction
Scyliorhinus canicula (Linnaeus, 1758) is a species of shark easy to manage in an aquarium and easy to observe. Much research about its endocrinology, nervous system, reproduction, physiology, etc., have been performed since 1904 [1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16] and it is considered as an “emerging chondrichthyan model” [10]. The aim of this study was to evaluate the ability of immature specimens of Scyliorhinus canicula and of Scyliorhinus stellaris (Linnaeus, 1758) to adapt to changes in salinity, by measuring the consumption of oxygen of three small spotted catshark specimens and six nursehound specimens. During aquarium husbandry of juvenile nursehounds, it was observed that they were able to resist low salinity conditions; infact, in the oviparous elasmobranchs, the ability of the embryo to regulate salts and urea is present at the earliest stage of development [17]. The need for elasmobranch embryos to regulate osmolytes was reduced or delayed as viviparity evolved [17]. Observations were made in order to test their oxygen consumption at low salinity conditions. This stress can occur in fishes as a consequence or reaction to different environmental conditions. It is known as, the General Adaptative Syndrome (G.A.S.), in which biochemical and physiological changes occur, such as the production of ACTH and corticosteroids with a subsequent reduction of Na+ and Cl- and loss of K+, an increase of blood sugars and blood pressure, and also an increase in oxygen consumption [18]. The mechanism of gill ventilation in Scyliorhinidae has been shown to be fundamentally the same as that found in teleost fishes [19, 8, 9]. The flow across the gills is maintained partly as a result of the increased pressure in of gill resistances but also because of the suction pump action of the parabrachial cavities [20]. Previous studies, performed in captivity, indicate that, in experimental conditions, Scyliorhinus canicula consumes 20-25 cm3/kg/hr of oxygen at a temperature of 12°C. These measurements were made after relaying the specimens into cylinders, where it was regulated direct water flow to the gills. The oxygen consumption was calculated in different gradients of hydrostatic pressure (Δp) and pressure (PO 2) through the gills. A ventilation volume of 0.43 l/min/kg for small spotted catsharks, Scyliorhinus stellaris was measured by Piiper and Schumann (1967). The experiments were designed to test if differences appeared in oxygen consumption at different salinities, and to obtain preliminary data on the osmotic stress to the fish to see if this could influence the respiratory processes. Observations were made in 2006 for Scyliorhinus stellaris and in 2010 for Scyliorhinus canicula.
Materials and methods
Test Scyliorhinus stellaris (2006)
The observations were made in the Argentario Mediterraneum Aquarium. The six tested specimens were maintained in four glass tanks, three of them with a volume of 13.5 litres (l), the last with a total volume of 30l. One specimen was maintained in each tank of 13.5 l and three specimens were placed in the 30l. All tanks were equipped with an inner filter and an air diffuser, sand in the bottom, and some small rocks in an attempt to reproduce a natural environment. All juveniles were approximately one year old. Water parameters (T°, pH, Salinity, Nitrites) were measured daily. Tests were performed two times per week for three weeks. Nine tanks of 6.2 l were used for the experiments, six of them with animals and three without to compare diffused oxygen level differences in the sea water during the test. The sea water, arriving directly by pipeline from the sea after 50 µm filtration, was prepared the day before at a fixed salinity rate (adjusted using freshwater) and oxygenated all night. The dimensions of the experimental tanks were 18 cm high, with a water level of 13 cm, 30 cm long, 16 cm wide, with a volume of 6.24 l; an air diffuser was used before experiment, no filter system. The experiments were performed between 10:00 hrs to 15:00 hrs. The starting oxygen level was 8 mg/l for each experiment. The specimens were maintained in the test tanks for five hours and data on oxygen levels were obtained ever 30 min with field oximeter calibrated daily (Hanna HI 9142, company, place made) [21]. The salinity levels in the tanks were three tanks each at 29.5 ‰, 33.5 ‰, and three at 37.5‰, for each group plus one tank without animals as control.
Test Scyliorhinus canicula (2010)
During the nine weeks of experiments, three replicate fishes, ranging in size between 38 and 48 cm total length, housed at the Aquarium Mondo Marino of Massa Marittima (GR), were available in four different pools. These tanks contained a total volume of 82 litres (l), and were free of filter or skimmer. The tubs were filled with water decanted in two large non-toxic fibreglass cylinders in the lab, 2000 l each. Water was drawn periodically directly from the sea in the municipality Follonica, in the locality of Pratoranieri. The preparation consisted in filling the three tanks the night before the experiment with water drawn directly into the sea in locations indicated above and in bringing the salinity, with the addition of reverse osmosis water, to the previously established values. The tanks were connected to an oxygenator with a porous stone, for the night before, leading to a saturated oxygen concentration. The next morning oxygenators were disconnected, and the dissolved oxygen measured, as well as temperature and pH (at time zero, t0). The sharks were then placed one in each tank of different salinity. The values were 38 ‰, 30 ‰, 35 ‰, and tank N° 4 contained sea water salinity 35 ‰ which served as control. Every shark remained in the same tank for five hours, to be transferred to its original tank. This protocol was repeated for four days per week (Tuesday to Friday). Each specimen was tested for each salinity. From time zero (t0) the measurements were performed every hour five times, of dissolved oxygen and temperature both in the tanks that housed the animals and in that of the control tank; therefore six oximeter measurements for each tank (t0- t1- t2- t3- t4- t5). The pH was measured only at t0 and to t5. The observations were made during a nine-week period.
Results
Test 2006 Scyliorhinus stellaris
Data collected on the level of dissolved oxygen in the tank showed little oxygen consumption. During approximately the first 60-90’ the oxygen consumption was, on average, approximately 0.6-0.8mg/l per hour per fish; after this adaptation time, the consumption become constant to 0,2-0,3mg/l per hour in the three different groups for the remaining four hours (figures: 1-3).
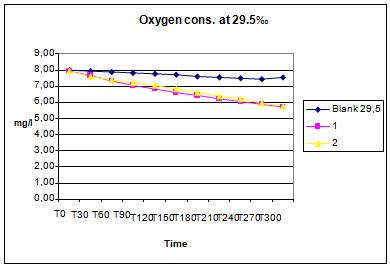
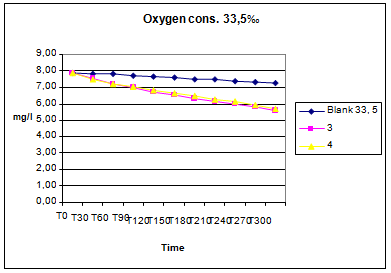
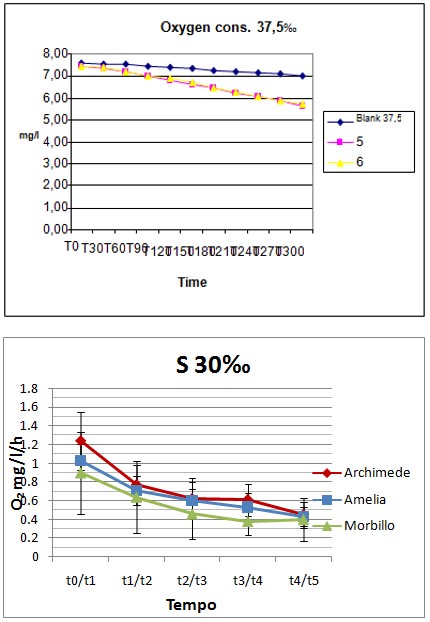
Test 2010 Scyliorhinus canicula
Data collected in this first series of observations indicate an increase in the oxygen consumption within the first two hours of experimentation for all the specimens in all conditions of salinity, and a stabilisation of consumption in the last three hours of the experiments. The average consumption in the first two hours was approximately 1 mg/l/h, and then stabilized near 0.4 mg/l/h in the remaining hours. The significance of differences was calculated using the Student's t-test. The temperature did not vary significantly during the five hours of experimentation but a slight decrease in pH and salinity was observed, regardless of the animal in the tub. This is presumed due to CO2 production by the sharks, and was not observed in the control tank.
Figure: 4
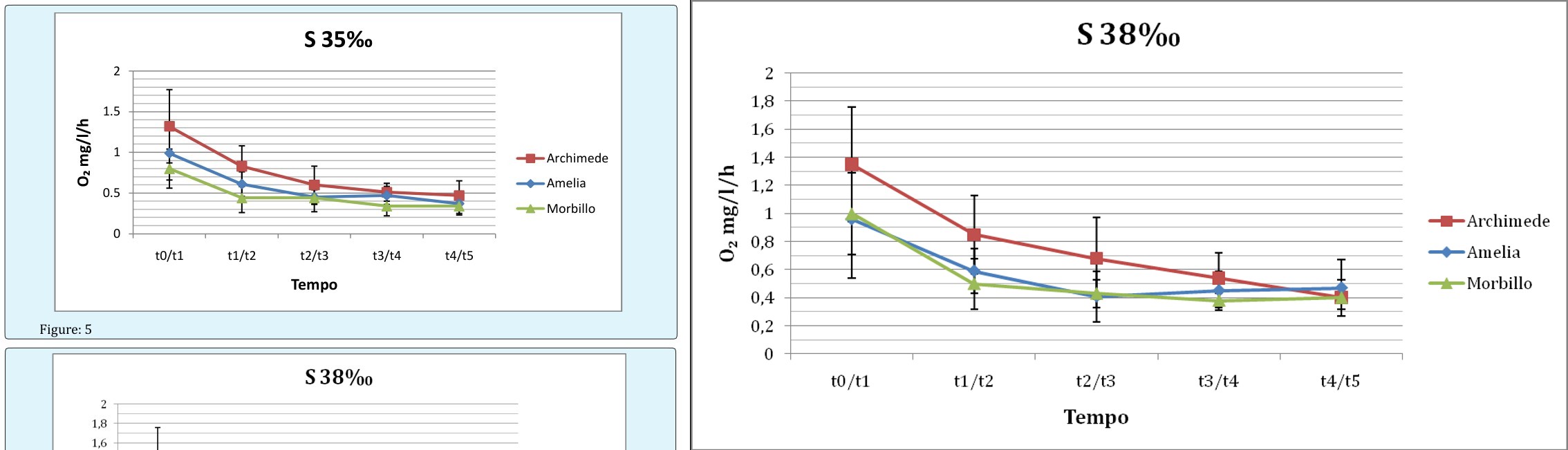
Figure: 6 Figure 4, 5, 6 Oxygen consumption, expressed in mg/l/h, as a function of time, for different animals exposed to the same salinity. Asterisks refer to the significant differences.
Conclusions
Nursehound and small spotted catsharks are sedentary sharks living on the bottom, adapted with a low metabolism like all the other Scyliorhinidae. During the embryonic development in egg cases they are able to the osmotic fluid changes but also about biochemical adaptations to stress, in order to evaluate particular evolutionary processes explaining this ability. The observations, strictly preliminary, need to be implemented with an increased number of specimens and widening the data set to younger as well as adult animals, supplemented with the collection of blood samples to examine the changes in values of ACTH and corticosteroids, which involve higher blood glucose concentrations and higher blood pressure which, in turn, are typically increased in General Adaptation Syndrome that is activated in stressful situations.
References
-
Borcea J (1904) Sur Un Las De Conformation Anormale De L'oviducte Droit Chez Line Petite Roussette Scyllium Canicula. Bulletin De La Societe Zoologique De France 29: 138-140.
-
Craik JCA (1978) Effects of Hypophysectomy on Vitellogenesis in the Elasmobranch Scyliorhinus Canicula L. Gen Comp Endocrinol 36 (1): 63-67.
-
Dobson S, Dodd JM (1977) The roles of temperature and photoperiod in the response of the testis of the dogfish, Scyliorhinus canicula L. to partial hypophysectomy (ventral lobectomy). Gen Comp Endocrinol 32 (1): 114-115.
-
Heisler N, Weitz H, Weitz AM (1976) Extracellular and intracellular pH with changes of temperature in the dogfish Scyliorhinus stellaris. Respir Physiol 26(2): 249-263.
-
Farner HP (1978) Embryonal development of the brain of the shark Scyliorhinus canicula (L.). I. Formation of the shape of the brain, the migration mode and phase and the structure of the diencephalon]. J Hirnforsch 19 (4): 313-332.
-
Jenkins N, Dodd JM (1982) Effects of ovariectomy of the dogfish Scyliorhinus canicula L. on circulating levels of androgen and oestradiol and on pituitary gonadotrophin content. J Fish Biol 21(3): 297-303.
-
Lissmann HW (1946) The neurological basis of the locomotory rhythm in the spinal dogfish (Scyllium canicula, Acanthias vulgaris); reflex behaviour. J Exp Biol 23(2): 143-161.
-
Piiper J, Schumann D (1967) Efficiency of Oxygen Exchange in the Gills of the Dogfish, Scyliorhinus Stellaris. Respir Physiol 2(2): 135-148.
-
Piiper J, Baumgarten-Schumann D (1968) Transport of O2 and CO2 by water and blood in gas exchange of the dogfish (Scyliorhinus stellaris). Respir Physiol 5(3): 326-337.
-
Rasch LJ, Martin KJ, Cooper RL, Metscher B, Underwood C, et al. (2016) An Ancient Dental Gene Set Governs Development And Continuous Regeneration Of Teeth In Sharks. Dev Bio 415 (2): 347-370 .
-
Sims DW (1996) The effect of body size on the standard metabolic rate of the lesser spotted dogfish. J Fish Biol 48(3): 542-544.
-
Sims DW, Southall EJ, Wearmouth VJ, Hutchinson N, Budd, et al. (2005) Refuging Behaviour In The Nursehound (_Scyliorhinus Stellaris)_: Evidence From Acoustic Telemetry. J Mar Biol Ass UK 85 (5): 1137- 1140.
-
Sumpter JP, Jenkins N, Dodd JM (1978) Hormonal Control of Steroidogenesis in an Elasmobranch Fish (Scyliorhinus Canicula L.): Studies Using a Specific Anti-Gonadotrophic Antibody. J Endocrinol 79(3 Suppl): 28-29p.
-
Taylor EW, Butler PJ (1971) Some Observations on the Relationship Between Heart Beat and Respiratory Movements in the Dogfish (Scyliorhinus Canicula L.). Comp Biochem Physiol A Physiol 39(2): 297-305.
-
Toulmond A, Dejours P, Truchot JP (1982) Cutaneous O2 and CO2 exchanges in the dogfish, Scyliorhinus canicula. Respiration Physiology 48(2): 169-181.
-
Van De Kamer JC, Zandbergen, MA (1981) The Hypothalamic-Hypophyseal System And Its Evolutionary Aspects In Scyliorhinus Caniculus. Cell Tissue Res 214(3): 575-582.
-
Kormanik GA (1993) Ionic and osmotic environment of developing elasmobranch embryos. Environmental Biology of Fishes 38(1): 233-240.
-
Roberts RJ (1990) Fish Pathology. Baillière Tindal Book. Cassell Ltd. Edition, pp. 289.
-
Hughes GM (1960) The Mechanism of Gill Ventilation in the Dogfish and Skate. J Exp Biol 37: 11-27.
-
Hughes GM, Ballintijn CM (1965) The Muscular Basis Of The Respiratory Pumps In The Dogfish (_Scyliorhinus Canicula_). J Exp Biol 43: 363-383.
-
Hans S. (1956) The Stress of Life. Mcgraw-Hill, New York.
-
Hughes GM, Umezawa S (1968) Oxygen Consumption And Gill Water Flow In The Dogfish (_Scyliorhinus_ _Canicula_). J Exp Biol 49(3): 557-564.
-
Skomal G, Bernal D (2010) Physiological Responses to Stress in Sharks. In: J C Carrier, JA Musick, Heithaus MR (Eds), Sharks and Their Relatives Ii. Crc Press, pp. 457-488.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh