Lepas Lalandii sp. Nov: A Goose Barnacle (Crustacea) Species from DNA Barcoding
We analysed DNA based phylogenetic relationships of selected species of pedunculated barnacles, using species from the genus Chthamalus (order: Balanomorpha) as outgroup, to estimate the position of a new species of goose barnacle. Ninetythree individuals from 17 ingroups (including seven genera) and five outgroup species were studied. The pedunculated species included taxa from the order Scalpellomorpha a) superfamily: Lepadoidea (family Lepadidae; genera Conchoderma, Dosima, Lepas and family: Poecilasmatidae; genera Octolasmis and Poecilasma), and b) order Pollicipedomorpha (family: Pollicipedidae; genera Capitilum and Pollicipes). As the ingroup members represent a polyphyletic assemblage, our analysis was limited to estimating the status and the genetic relationship of the new species. We show that Lepas lalandii sp. nov. is monophyletic and a member of the Lepadidae, but separated from other Lepas. Full descriptions of Lepas lalandii sp. nov. will follow detailed morphological studies. This is conceivably the most extensive dataset for the individuals studied.
Introduction
DNA barcodes for animals, using the cytochrome oxidase 1 mitochondrial gene (CO1 or COI), is not only a particularly useful tool to accelerate species recognition, discrimination of biodiversity, facilitation of conservation efforts, stock management, ecosystem monitoring, but also useful for determining phylogeographic and speciation patterns. For example, it helps at identifying various life-forms (such as barnacle larvae and adults [1] carcass fragments, damaged specimens, prey in gut contents, etc. (for reviews of the above, see [2, 3]. Understanding the relationships of the diverse barnacles is increasingly important because they act as ecosystem engineers. They attach onto floating and other objects/substrata (e.g. rocks, boats, kelp, biofouling materials such as plastic bottles, various animals, feathers, etc.). Goose barnacles are a paraphyletic assemblage of species, characterised by the main body (capitulum) being raised above the substratum by a flexible stalk or peduncle. Most commonly “goose barnacle” is used to denote species of the monophyletic superfamily Lepadoidea, including those of the genus Lepas. Goose barnacles attach on up to 277 natural and anthropogenic substrata in marine environments [4, 5, 6, 7]. The free-floating or actively sailing substrata of goose barnacles contribute to their cosmopolitan distribution. As a result of their varied attachment adaptations, barnacles can have massive economic implications. For example, they cause drag when attaching to boats, increasing fuel consumption and it was calculated that fuel cost can increase to approximately $250 million US per annum [8, 9]. Pedunculated barnacles (“goose barnacles”) are particularly problematic because they extend far into the surrounding water compared to fouling acorn barnacles (Balanomorpha), but interestingly little antifouling research directly addresses these pedunculated forms. The stalks of some stalked barnacles (Pollicipes, Capitulum) are regarded as very tasteful (likened to crayfish). Large individuals (up to 5 cm long) of these species are gathered commercially and sold as luxury food items in countries such as China, Japan, Korea, Portugal, France, Spain, and Taiwan (https://en.wikipedia.org/wiki/ Capitulum_mitella). They colonise in large numbers and most valued when fresh, but can also can be preserved (canned/ tinned) and are expensive (up to €100 per plate and €200 per kilogram, e.g. see https://en.wikipedia.org/wiki/Goose_ barnacle). The high price reflects the difficulty of collection in the energy-rich upper rocky intertidal zone. As far as could be ascertained, aquaculture of stalked barnacles has unfortunately not been successful.
Phylogenetic relationships of stalked barnacles were until recently poorly understood and the taxonomy does not always reflect monophyletic units [10]. The above-mentioned robust molecularly based phylogeny (down to family) revealed that much uncertainty remains at lower levels. This is due to many species that were poorly described, with some genera and/or species being synonymised, and then separated in the past (see Discussion). The morphology of stalked barnacles varies extensively, reflecting their polyphyly, but it is uniform for members of the Lepadoidea, and especially for the sister families Lepadidae and Poecilasmatidae [10]. We use a DNA barcoding approach for species identification and to study phylogenetic relationships. This was done by comparing the DNA barcodes of 104 barnacle individuals (more sequences for individuals are available but does not change the outcome of these results) from 22 species. Although the analysed species are sampled broadly in thoracican barnacles, our attention is limited to the position of the new species and thus the superfamily Lepadoidea. Results outside the lepadoidea is not discussed but see [10;12] and [11] for a review that included 239 barnacle species with phylogenetic information. We use species of the Balanomorpha as outgroup, since they are an assured monophyletic group and far separated from the Lepadidae, which have our particular attention.
Material and Methods
Species studied abbreviations thereof and authorities are presented in Table 1.
| Species: | Authority: | Abbreviation: |
|---|---|---|
| Chthamalus challengeri | Hoek 1883 | {Cc} |
| Chthamalus dentatus | Krauss 1848 | {Cd} |
| Chthamalus malayensis | Pilsbry 1916 | {Chm} |
| Chthamalus moro | Pilsbry 1916 | {Cmo} |
| Chthamalus neglectus | Yan & Chan 2004 | {Cn} |
| Capitulum metella | Foster 1980 | {Cm} |
| Conchoderma auritum | (Linnaeus 1767) | {Ca} |
| Conchoderma virgatum | Spengler 1789 | {Cv} |
| Dosima fascicularis | (Ellis & Solander 1786) | {Df} |
| Lepas anatifera | Linnaeus 1758 | {Lana} |
| Lepas anserifera | Linnaeus 1767 | {Lans} |
| Lepas australis | Darwin 1851 | {Laus} |
| Lepas pectinata | Spengler 1793 | {Lp} |
| Lepas testudinata | Aurivillius 1894 | {Lt} |
| Octolasmis angulata | (Aurivillius 1894) | {Oa} |
| Octolasmis cor | (Aurivillius 1892) | {Oc} |
| Octolasmis warwickii | Gray 1825 | {Ow} |
| Poecilasma litum | Pilsbry 1907 | {Pl} |
| Poecilasma obliqua | Hoek 1907 | {Po} |
| Policepes elegans | (Lesson 1831) | {Pe} |
| Policepes policepes | (Gmelin 1789) | {Pp} |
Table 1: Species studied (with outgroup species in bold), authorities and abbreviations used.
Five species (14 individuals) of Chthamalus Ranzani, 1817 were selected as outgroups. {Cd} and {Ca} (sampled from a beached Humpback whale, Megaptera novaeangliae, Borowski 1781) were deposited on BOLD (www. barcodinglife.org) as described in [8]. DNA extraction and analyses followed [3, 8]. We only studied adult specimens, and individuals with GenBank numbers between MT563421 and MT563441 were collected for the present study (see LB numbers on Figure 1). {Lana} (Figure 2a), {Lans} (Figure 2b) and L. lalandii sp. nov. (abbreviated henceforth as {Ll};
Figure 2c), were collected at Lampies Bay (-32.715, 17.961) at the West Coast of South Africa. {Lana} and {Lans} were sampled from a cast ashore plastic bottle (Figure 2d). {Ll} was collected from the roots of cast ashore sea bamboo at the West coast of South Africa, a species of kelp (Ecklonia maxima (Osbeck) Papenfuss, Figure 2e). No distinct morphological characteristic was noted thus far, and no specific distributions of the new species exist. Therefore, both were not helpful in assigning a species name for the new taxon. The species name assigned “lalandii” translates to gift from God. It is the scientific name for many species, including West coast rock lobsters, also found locally (South Africa). The individuals were deposited at the Natural History Museum of Denmark. Additional sequences were mined from GenBank (see Figure 1 for GenBank accession numbers).
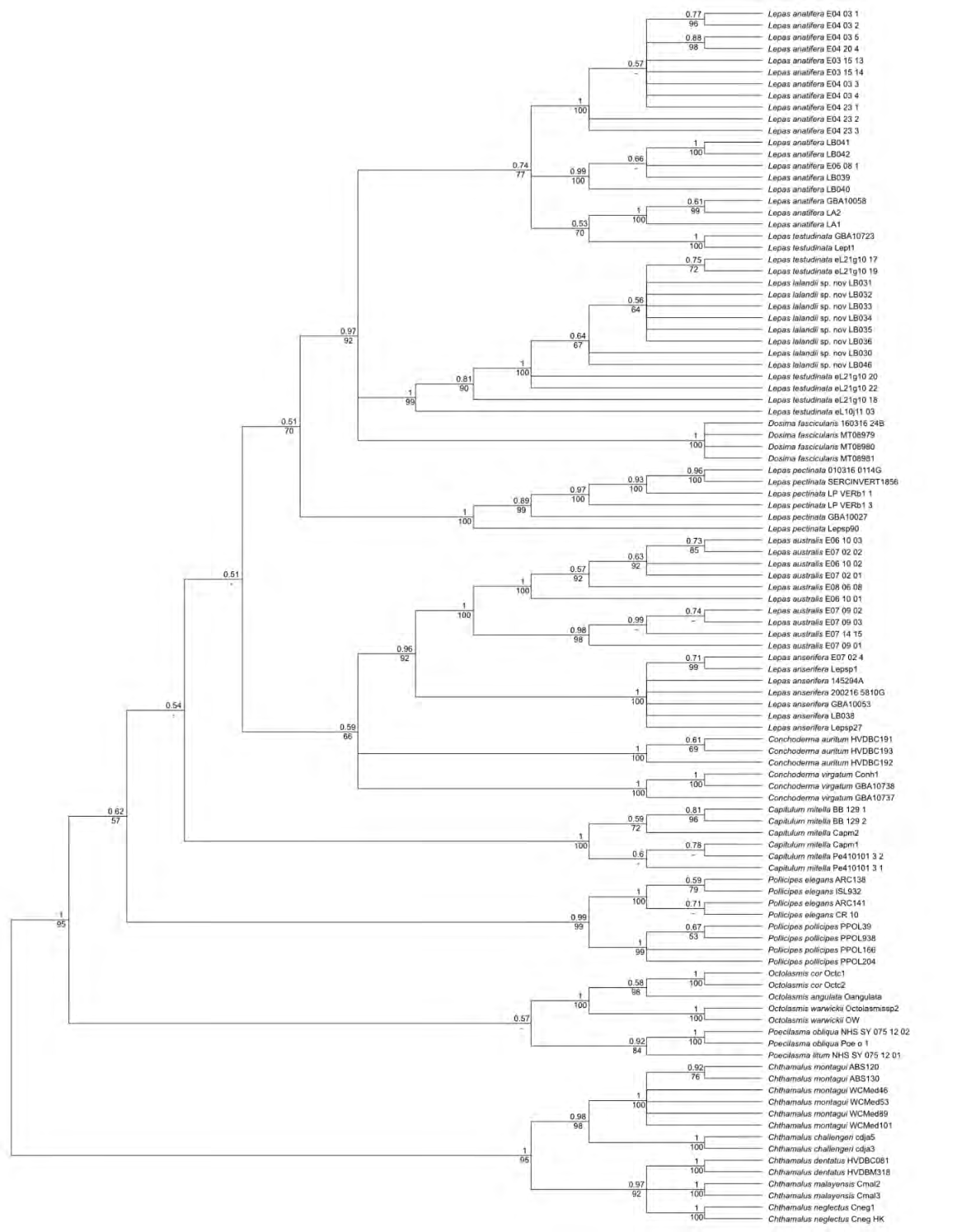

All sequences were edited in Geneious v.8.1.9 (https:// www.geneious.com), aligned using MAFFT v7.017 [13], and subsequently manually adjusted where needed. The maximum likelihood (ML) analysis was performed on the aligned sequences of stalk barnacles using IQTREE2 v 2.1.2 [14, 15]. A single consensus tree was generated from 1,000 ultrafast bootstrap (UFB) approximations. The Bayesian inference analysis (BI, [16, 17]) of the aligned sequences was done on the CIPRES online portal (http://www.phylo. org/sub_sections/portal, [18]) using MrBayes v.3.2.2 on XSEDE. Four chains were run for 10,000,000 Markov chain Monte Carlo (MCMC, [19]) generations. The sampling of trees occurred every 1,000 generations and the burnin was set to 1,000,000. A consensus tree was constructed from the remaining trees. The values were evaluated using the following scale: below 0.95, weakly supported; 0.95-1.0, strongly supported.
Results and Discussion
The rapid advancements in phylogenetic techniques have enabled the identification of cryptic species and discovery of new species, especially in the marine environment.
Particularly, for cosmopolitan species such as barnacles, the presence of cryptic diversity [8] has implications for understanding species boundaries. {Lana} and {Lans} were sampled from a cast ashore plastic bottle, and although biofouling is a major concern (especially in the marine environment), in this study like others (see Introduction), it was a habitat for the above-mentioned barnacle species. The minimum base pair (BP) lengths=418, maximum=639, median=590 and mean=597. The number of consistent sites=356, the number of parsimony informative sites=264 and Best-fit model according to BIC: TVM+I+G (found with ModelFinder).
These are some interesting synonyms from Wikipedia for species in the present studied: {Cv} was L. virgata Spengler, 1790; {Df} was L. fascicularis Ellis and Solander, 1786; {Ca} was L. aurita Linnaeus, 1767 and {Cm} was L. mitella Linnaeus, 1758. This not only confirms the affinity of the former three species with the Lepadidae, but also the confusion thereof with a species from the Poecilasmatidae ({Cm}; note: with Lepas as synonym). In addition, [5] reported four Lepas species ({Lana}, {Lans}, {Lp}, {Lt}) in a cluster with {Cm}), {Cv} and Conchoderma hunteri (Owen,
1830) sample GBA10738 (which is now a synonym for {Cv}). They also found three Octolasmis ({Oa}, {Oc}, {Ow}) spp. in a separate clade, with five Chthamalus spp. (including four in the present study: {Cc}, {Chm}, {Cmo}, {Cn}) sister to the group mentioned in the sentence above. [1] found same for these Lepas (plus {Laus}), {Ca} and {Cv} in one cluster. They ([1]) reported two Poecilasma spp. and four Octolasmis spp. in another cluster; with {Cm} and {Pp} in a separate cluster. Four Chthamalus spp. (including {Cc} and {Cmo} as in the present study) was sister to all the above [1]. The results from the above-mentioned two studies, therefore, found different groupings of these members of the families Lepadidae, Poecilasmatiday and Pollicipedidae, which is the motivation for including it in the present study.
COI Analyses
Chthamalus is an excellent choice as outgroup (as mentioned above in [1, 5]). Peréz-Losada, et al. [1] reported Poecilasmatidae sister to the Lepadidae as also found by us (Figures 1 & 4) and [5] for all three Octolasmis spp. Posterior probabilities (PP) values are 100% for the outgroup species, where n>1 (Figure 1). Specimen sampling is an important consideration to resolve genetic variation within and differentiation between species. Between 5-11 individuals per species is more commonly studied, but one (if an existing DNA library for the same species already exists) or two individuals per species have been used to facilitate reliable identifications [20]. Our average sample size of approximately six individuals per species for the in groups studied, can thus be considered as adequate. COI is extremely reliable for species identification. Nevertheless, HVDB experienced an initial identification problem with the LB038 sample: it was included in the field identification using the photographs and description in [21] as {Lana} because we collected it in-between the other samples with shiny white shell plates. Lepas species are almost impossible to differentiate morphologically (see the 44 identical character states published in [1]) and they are cosmopolitan, further complicating species discrimination. However, the COI analysis clearly showed that LB038 must be {Lans} and not {Lana} (with 100% accuracy from its nucleotide sequence BLAST results); see also Figure 3 for diagnostic character differences and Figure 1 for phylogenetic relationships). HVDB also identified {Ll} as {Lt} using [21] due to its brownish-yellow shell plate colour and smaller size (compared to the bright, shiny white shell plates and larger individuals collected at the same locality). However, its barcodes are unlike those of {Lt} (see Figure 3 for diagnostic sequences, Figures 1 & 3 for phylogenies). A more comprehensive study showed distinct differences between the latter two species and presented haplotype genealogies for comparison [11]. From the present results (Figure 1), it is evident that the species from South Africa in [11] is also the new species {Ll} as combined in Figure 4.
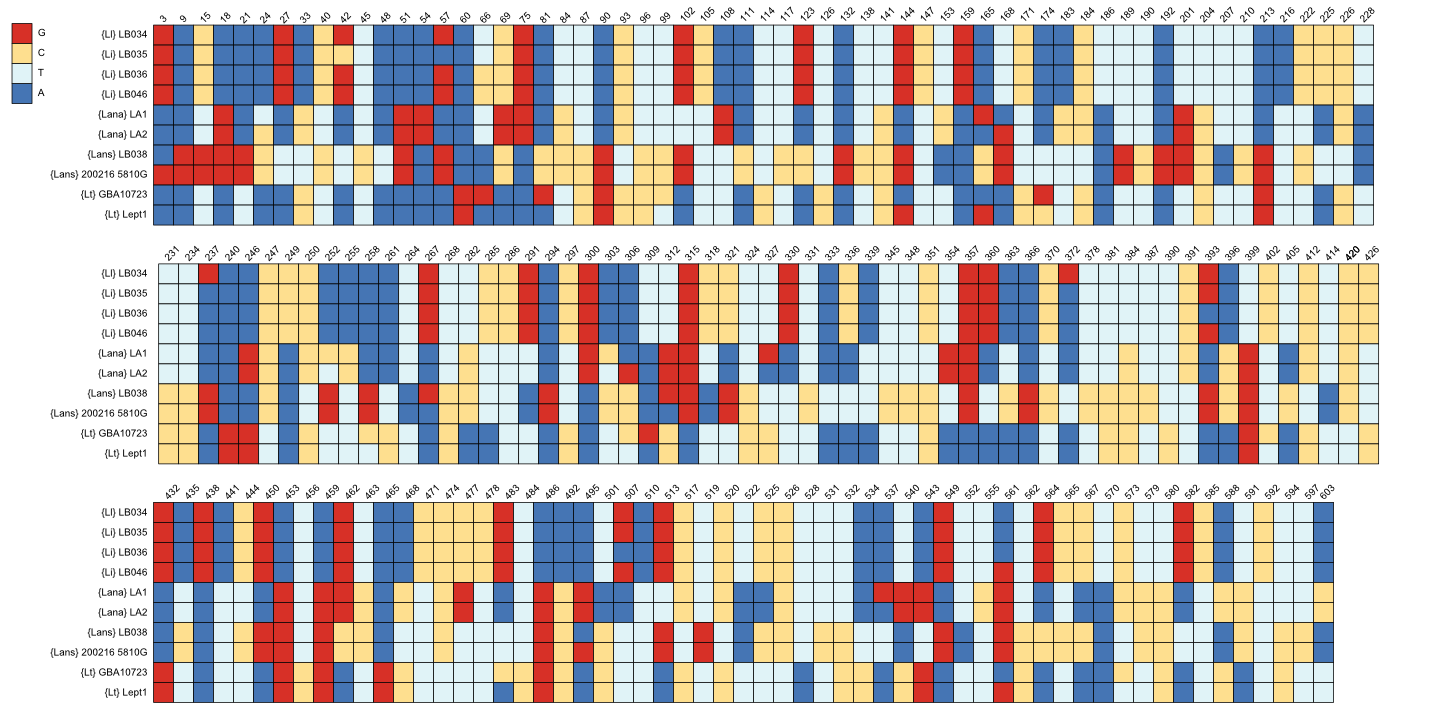
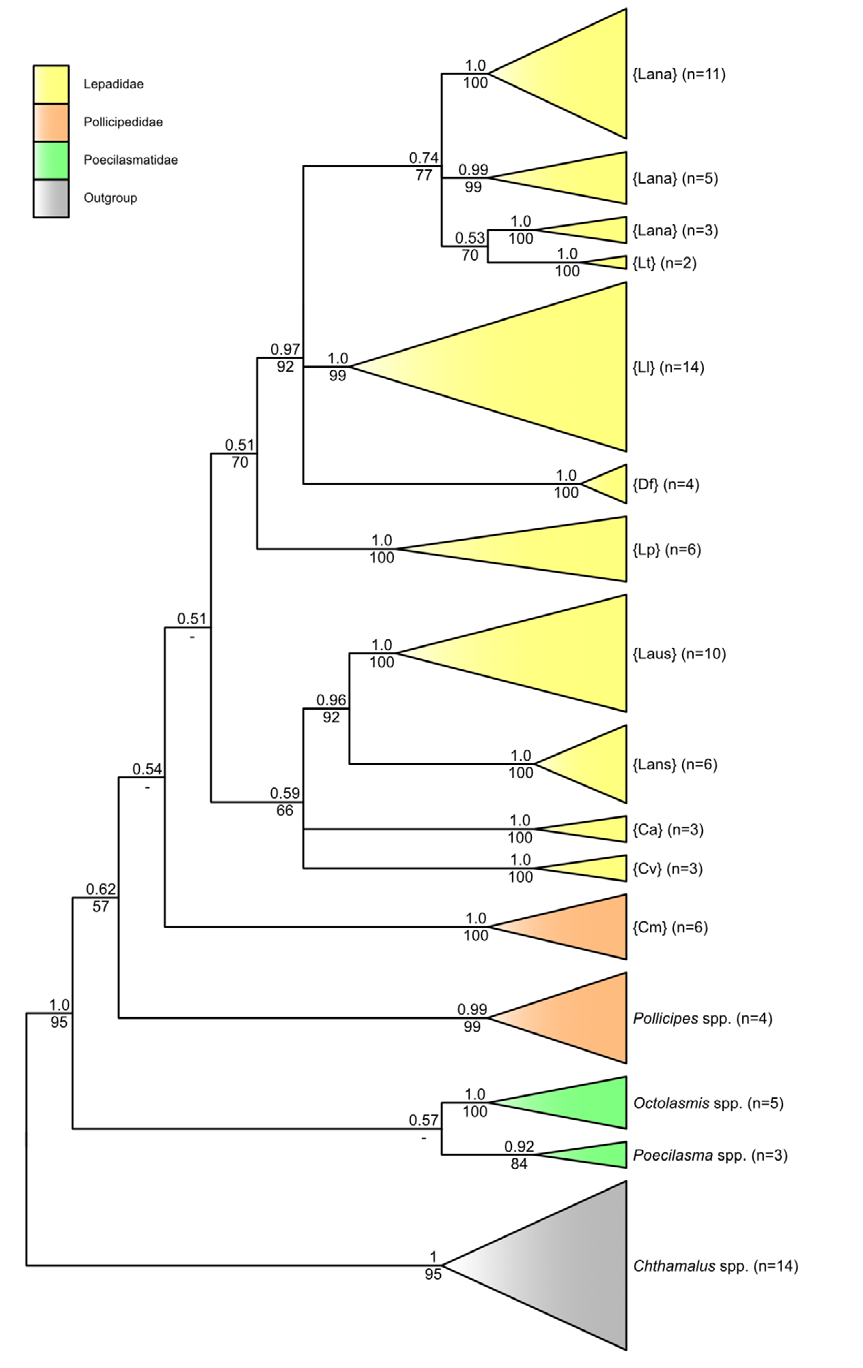
For fifteen of the sixteen species studied, where sample size (n) > 1, monophyly is recovered with high PP values (average ≥0.99; Figure 1). The only exception is the polyphyletic {Lana} cluster. The {Lana}/{Lt} grouping is not surprising, since one of the synonyms for {Lt} is Lepas anatifera var. testudinata Nilsson-Cantell, 1928 [22]. This indicates that more detailed research is required to resolve it and that more species could be involved, conceivably due to geographical divergence (e.g. note the one branch of {Lana} contains individuals with LA and GBA numbers, and clusters with {Lt}, whereas the other two clusters have different accession numbers; Figure 1). The individuals with accession numbers E03 and E04 are from Chile, the two LA samples are from Taiwan, E06 and GBA10058 are from the Gulf of Mexico, and ours are from South Africa. The two {Lt} individuals are also from Taiwan.
In a more recent study, [11] used a phylogenetic approach (from mitochondrial 16S and COI sequences, and the nuclear 18S gene) to elucidate patterns of inter-and intra-species divergence of five species of Lepas. They also reported similar results (i.e. two groups of {Lana}, and with a split between {Lt} individuals from South Africa and Australia). To conclude, COI is sufficient for species identification in goose barnacles, but not for differentiation (phylogenetic resolution) with poor backbone PP support. This was also reported in previous studies. Nevertheless, it is evident that the new species is distinct from all others studied. The marine fauna of South Africa needs more extensive studies (e.g. new species are discovered as in the present study and [23] who described a new jellyfish species from the southern-west coast of South Africa).
Acknowledgements
“We are grateful to the CCDB (University of Guelph, Canada) for some of the DNA sequencing. A part of the project was funded by the Government of Canada through Genome Canada and the Ontario Genomics Institute (2008-OGI- ICI-03). The research was supported by the ACDB (University of Johannesburg) and samples were deposited at the Natural History Museum of Denmark. We thank Ronny van Rooyen for assistance with sample collections”.
References
-
Perez Losada M, Harp M, Hoeg JT, Achituv Y, Jones D, et al. (2008) The tempo and mode of barnacle evolution. Molecular Phylogenetics and Evolution 46(1): 328-346.
-
Bezeng BS, Davies TJ, Daru BH, Kabongo RM, Maurin O, et al. (2017) Ten years of plant DNA barcoding at the African Centre for DNA Barcoding. Genome 60(7): 629- 639.
-
van der Bank FH, Greenfield R, Daru BH, Yessoufou K (2012) DNA barcoding reveals micro-evolutionary changes and river systemlevel phylogeographic resolution of African silver catfish, _Schilbe intermedius_ (Actinopterygii: Siluriformes: Schilbeidae) from seven populations across different African river systems. Acta Ichthyologica et Piscatoria 42(4): 307-320.
-
Ashton GV, Davidson IC, Geller J, Ruiz GM (2016) Disentangling the biogeography of ship biofouling: barnacles in the Northeast Pacific. Global Ecology and Biogeography 25(6): 739-750.
-
Chen HN, Hoeg JT, Chan BKK (2013) Morphometric and molecular identification of individual barnacle cyprids from wild plankton: an approach to detecting fouling and invasive barnacle species. Biofouling 29(2): 133-145.
-
Rech S, Pichs YJB, Garcıa Vazquez E (2018) Anthropogenic marine litter composition in coastal areas may be a predictor of potentially invasive rafting fauna. PLoS ONE 13(1): e0191859.
-
Whitehead TO, Biccard A, Griffiths CL (2011) South African pelagic goose barnacles (Cirripedia, Thoracica): substratum preferences and influence of plastic debris on abundance and distribution. Crustaceana 84(5-6): 635-649.
-
Bezeng SB, van der Bank HF (2019) DNA barcoding of southern African crustaceans reveals a mix of invasive species and potential cryptic diversity. Plos One, pp: 1-12.
-
Sigler K (2016) Barnacles. Business and Economics Journal 7:2.
-
Chan BKK, Dreyer N, Gale A, Glenner H, Saucedo CH, et al. (2021) The evolutionary diversity of barnacles, with an updated classification of fossil and living forms. Zoological Journal of the Linnean Society, pp: 1-58.
-
Schiffer PH, Herbig HG (2016) Endorsing Darwin: global biogeography of the epipelagic goose barnacles _Lepas_ spp. (Cirripedia, Lepadomorpha) proves cryptic speciation. Zoological Journal of the Linnean Society 177 (3): 507-525.
-
Ewers-Saucedo C, Owen CL, Pérez-Losada M, Hoeg JT, Glenner H, et al. (2019) Towards a barnacle tree of life: integrating diverse phylogenetic efforts into a comprehensive hypothesis of thecostracan evolution. PeerJ.
-
Katoh K, Misawa K, Kuma KI, Miyata T (2002) MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Research 30(14): 3059-3066.
-
Nguyen LT, Schmidt HA, Von Haeseler A, Minh BQ (2015) IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Molecular Biology and Evolution 32(1): 268-274.
-
Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams MD, et al. (2020) IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Molecular Biology and Evolution 37(5): 1530-1534.
-
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17(8): 754-755.
-
Ronquist F, Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19(12): 1572-1574.
-
Miller MA, Pfeiffer W, Schwartz T (2010) Creating the CIPRES science gateway for inference of large phylogenetic trees. Gateway Computing Environments Workshop, pp: 1-7.
-
Geyer CJ (1991) Markov Chain Monte Carlo Maximum Likelihood. Computing Science and Statistics (1): 156- 163.
-
Zhang AB, He LJ, Crozier RH, Muster C, Zhu CD (2010) Estimating sample sizes for DNA barcoding. Molecular Phylogenetics and Evolution 54(3): 1035-1039.
-
Branch GM, Griffiths CL, Branch ML, Beckley LE (2010) Two Oceans. A guide to the marine life of southern Africa. Struik Nature Cape Town, pp: 456.
-
Chan BKK, Prabowo RE, Lee KS (2009) Crustacean fauna of Taiwan: barnacles, volume 1-Cirripedia: Thoracica excluding the Porgomatinae and Acastinae. National Taiwan Ocean University, pp 297.
-
Ras V, Neethling S, Engelbrecht A, et al. (2020) There are three species of _Chrysaora_ (Scyphozoa: Discomedusae) in the Benguela upwelling ecosystem, not two. Zootaxa 4778 (3): 401-438.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh