Biochemical and Morphological Responses in Amazon Fish, Piaractus brachypomus Exposed To 2,4-D + Picloram Herbicide
Pesticides as herbicides present an elevated potential for contaminating the aquatic environment, being able to create negative effects for organisms that are not the target, like fish. We evaluated the morphology and function of the hepatic, branchial and renal tissue on Piaractus brachypomus exposed to the herbicide based on 2,4-D + Picloram. Fishes were exposed to a sublethal concentration of the herbicide Norton® (1.1 g L-1, corresponding to 420 mg L-1 of 2,4D and 100 mg L-1 of picloram) for 96 h. After exposure, the plasma, gills, liver, kidney and white muscle were sampled for biochemical and morphological analyses. Acute exposure induced an increase on plasmatic glucose and decrease of muscular glycogen, while the plasmatic protein was also decreased. Tissues GST-GSH enzymatic biotransformation pathway and lipid peroxidation (LPO) were unchanged after acute exposure. Histopathological indexes analysis showed alterations on gills, liver and kidney which was evidenced morphofunctional damage by histopathological organ indexes. The results demonstrate that the herbicide induce different organs responses characterized by morphological and metabolic alterations on P. brachypomus. The adjustments of energy demand and gill, liver and kidney morphological changes produced by Norton® may affect the capacity of vital functions and capacity for development and reproduction of the specie.
Introduction
Weed chemical control present on agriculture is performed mainly by herbicide application, which places Brazil as one of the main users in this market [1]. Formulation of herbicides that associates 2,4-dichlorophenoxyacetic acid (2,4-D) and 4-amino-3,5,6-trichloropicolinic acid (Picloram) are broadly recommended for post-emergent application, because presents an elevated chemical efficiency and do not allow regrowth of unwanted species [2]. Due to the widespread use of these compounds, both are frequently detected on reservoirs that provide potable water and provide an elevated potential for contamination of aquatic environments [3]. In general, harmful actions of mixture of 2,4-D and picloram reported on organisms include teratogenic, neurotoxic, cytotoxic and hepatotoxic effects [4, 5, 6, 7].
Liver is the organ most associated with detoxification and biotransformation process, in addition to performing other important functions such as protein, carbohydrate and lipid metabolism [8]. In rats, association of 2,4-D and Picloram created an inhibition of the liver energetic metabolism, demonstrated by modifications on intermediate metabolites [9]. In fish, the exposure to these mixtures can induce morphophysiological variations on hepatic tissue [5]. Therefore, plasmatic biomarkers contribute to the evaluation of the effects of pollutants on hepatic tissue, such as the acid phosphatase (ACP), alkaline phosphatase (ALP) and aminotransferase (alanine aminotransferase-ALT; aspartate aminotransferase-AST) activity. These biomarkers are useful because indicates hepatocyte membrane integrity which are responsible for transportation of phosphorylated intermediates and are associated with hepatic lesions which promote cellular breakdown [10, 11].
In addition to hepatic responses, other organs are considered targets for environmental changes, such as the gills, that work as a barrier between the external and internal environment [12]. Damage to these tissues can impair essential physiological functions for fish, such as gas exchange, osmoregulation and excretion, promoting severe consequences to individuals [13]. Therefore, there are reports that 2,4-D and Picloram combination was capable to induce morphological damage on gills and caused the decrease of ions regulation capacity on catfish (Ictalurus punctatus) [5]. In teleost, the kidneys are also one of the first organs to be affected by pollutants, due to the intense blood flow and for participating on excretion and hematopoietic activities [14]. For this reason, herbicide activities are frequently associated with dysfunctions on this tissue [15, 16].
The majority of xenobiotics can initiate oxidative stress, caused by reactive oxygen species (ROS) produced during metabolic transformation in organisms. The antioxidant defense system consists of a variety of enzymes and non- enzymatic antioxidants, such as the GST-GSH system, which operates on phase II of detoxification, conjugating metabolites produced by phase I of biotransformation. At normal conditions, after conjugation of toxic compounds the product is excreted from the cell, avoiding potential cellular damage [17]. However, when ROS is produced on levels that exceed the capacity for neutralization of the antioxidant defense system, occurs the oxidative damage, causing cellular damage such as lipid peroxidation (LPO) [18].
In general, the biochemical and physiological adjustments may result on alterations that compromise organ’s function in fish [15, 19]. In this context, this study investigated morphofunctional damage of the hepatic, branchial and renal tissue in Piaractus brachypomus exposed to the commercial herbicide based on 2,4-D + picloram, using integrated plasmatic, histopathological and biotransformation biomarkers.
Materials and Methods
Fish
Piaractus brachypomus juveniles, n = 20 (body mass = 38.67 ± 6.92 g and total length = 13.38 ± 2.26 cm) were obtained from a pisciculture on Xinguara (Pará, Brazil) and transported to the Laboratory of Morphophysiology and Biochemistry of Neotropical fishes, at Federal University of Northern Tocantins. The fish were acclimated for a minimal period of 30 days in tanks with capacity for 5000 L, with constant water supply. The animals were fed daily with commercial fish food containing 40% of protein and maintained in a natural photoperiod.
Analyses of the physical-chemical variables of water from the period of acclimation were evaluated daily: absence of nitrite; temperature of 25.5 ± 0.04 ºC, water hardness of 50 ± 0.00 CaCO3 (ppm); pH of 7.2 ± 0.01; dissolved oxygen of 6.1 ± 0.01 mg L-1 and ammonia of 0.005 ± 0.00 ppm.
Experimental Design
After the period of acclimation, the fish were randomly divided in two groups (n = 10) and reallocated in aquariums of 200 L with monitored physical-chemical parameters and with continuous artificial aeration. The commercial formulation Norton® SL (soluble concentrate) was used as source of the associated active principles 2,4-D and picloram. A single sub-lethal concentration of 1.11 g L-1 of the herbicide (corresponding to 420 mg L-1 of 2,4D and 100 mg L-1 of picloram) was used for the 96 hours exposure.
After the exposure, fish were anesthetized with benzocaine (0.1 g L-1) and a blood sample (1 mL) was collected through the caudal vein with heparinized syringes. After biometry, euthanasia was done by medullar section and samples of liver, gills, kidney and muscle were collected and divided in aliquots. Samples of plasma and tissue were kept at -80 ºC for biochemical analyses and the remaining aliquots of gills, liver and kidney were fixated with 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.3) for morphological analyses. This experiment was approved by the Animal Ethics Committee (CEUA) of Federal University of Tocantins (Proc. N. 23101049/2019-65).
Biochemical Analysis
Samples of liver, gills, kidney and muscle were homogenized in potassium phosphate buffer 0.2 M, pH 7.8 or Tris-HCl buffer 100 mM, EDTA 2 mM and MgCl2 5 mM, pH 7.7 and centrifuged at 10000 g, 4°C for 10 min, the supernatant provided the enzymes for hepatoxicity, enzymatic and non- enzymatic antioxidant biomarkers and metabolites.
Determination of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activity on plasma (U mL-1) were determined by the commercial kit Labtest® (ref. 1008 and 109, Brazil) according to the manufacturer’s protocol. The method was based on the extinction of NADH and absorbance of 340 nm is directly proportional to ALT and AST activity in the sample. Commercial kit Labtest® (ref. 79 and ref. 39, Brazil) was used to determine the activity of plasmatic (mmol min mL-1) alkaline phosphatase (ALP) and acid phosphatase (ACP), according to the manufacturer’s protocol. ALP was determined based on the quantity of p-nitrophenol produced through the hydrolysis of p-nitrophenyl phosphate, measured at 405 nm and the enzymatic action of ACP was determined using a thymolphthalein monophosphate substrate.
Metabolites extracts were prepared after tissue homogenization or plasma deproteinization with trichloroacetic acid 20%. According to the methodology of Gentzkow CJ, et al. [20], ammonia (U mL-1 plasma) was quantified with photometric quantification as acid extracts. Plasmatic glucose (mg dL-1) was determined photometrically as neutral extracts with a commercial kit Labtest® (ref. 133) according to manufacturer’s protocol. Hepatic and muscular glycogen quantification (μmol of glycosyl-glucose g tissue-1) was established by the concentration of total reducing sugar in the sample, according to Bidinotto PM, et al. [21] and total protein content (mg g tissue-1 or mg mL plasma-1) was determined using Bradford reagent and bovine serum albumin (1 mg mL-1) as the standard, according to Kruger NJ [22].
To analyze the biotransformation of GSH-GST, GST (U mg protein-1) was determined according to the method described by Habig WH, et al. [23], using 1-chloro-2,4-dinitrobenzene (CDNB) as substrate. Absorbance of 340 nm was observed for 3 minutes with 30 seconds intervals. GSH concentration (nmol GSH mg protein-1) was defined using the methodology by White CC, et al. [24], whereas it was measured by naphthalene-2,3-dialdehyde (NDA) 10 mM conjugation with the supernatant GSH, producing a fluorescent compound. The measuring was made at excitation/emission of 742/528 nm against a GSH standard (40 nM).
Oxidative damage of lipids was evaluated with LPO (nmol mg protein-1), through FOX method (oxidation of iron by xylenol orange) according to the description of Jiang Z, et al. [25]. A 30 µL of sample + 270 µL of reaction medium (HPLC grade methanol 90%, 100 µM xylenol orange, 25 mM H2SO4, 4 mM BHT-butylated hydroxytoluene, 250 µM of FeSO4NH4-ferrous ammonium sulfate). The microplates were incubated at room temperature for 30 min in the dark and read at 560 nm, and compared to a standard curve of cumene hydroperoxide- CHP solution (1mM).
Morphological Analysis
Gills, liver and kidney samples were fixated in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.3) and dehydrated in a series of increasing ethanol concentrations and embedded in Historesin (Leica). Tissues were sectioned in 3 µm and stained with toluidine blue and basic fuchsine. For the blind histopathological analyses, a random field analysis was used. Tissue sections (5 per animal) were examined (25 random fields per section) using a light microscope Leica ICC50HD (400 x magnification).
Histopathological analyses were determined according to Paulino MG, et al. [19]. Alteration that each pathology causes in the analyzed organ, it was attributed an importance factor (IF): (1) easily reversible lesion with minimal pathological importance; (2) reversible in most cases when the stressor agent is neutralized, moderate importance; (3) irreversible lesion with extreme importance. Furthermore, to classify the frequency of each pathology during analyses, it was attributed a score: 0 = absence of lesion, 1 = rare occurrence, 2 = mild occurrence, 3= moderate occurrence and 4 = severe occurrence. The individual alteration index (Ialt) was determined by multiplying the score by IF and the organ histopathological indexes (Iorg) was determined as the sum of each Ialt for each animal (Iorg = ΣIalt).
Statistical Analysis
Data are expressed as mean ± S.E.M (standard error of the mean). Normal distribution was verified with D’Agostino & Pearson (n > 8) to test homogeneity of the variations between groups. For each parameter, the differences between the control and contaminated group were verified with an unpaired Student t-test or its correspondent nonparametric test Mann-Whitney, according to normality criteria, homogeneity and homoscedasticity. All statistical analyses were performed with GraphPad Prisma, considering differences when p ≤ 0.05.
Results
No fish died during the 96-h exposure to herbicide; therefore, the concentration containing 420 mg L-1 of 2,4D and 100 mg L-1 of picloram were sublethal. During exposure, fish behavior and color were unchanged.
Enzyme Activity and Metabolism
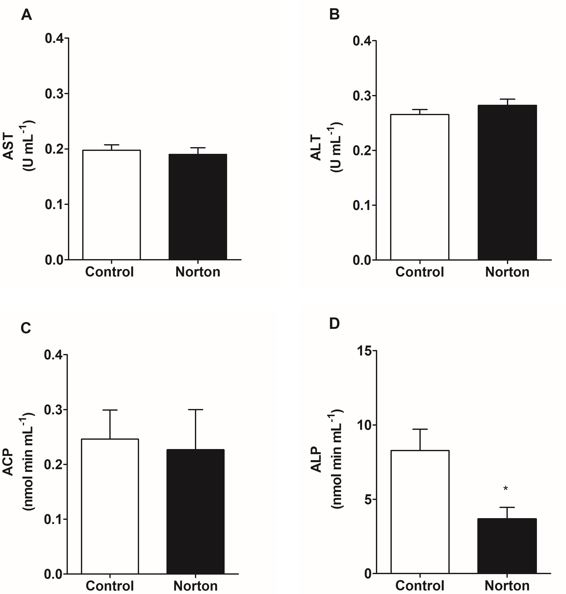
Biochemical plasma biomarkers responses to acute exposure to herbicide are shown in Figure 1. Activity of ALT, AST, ACP did not change and ALP decreased 56% regarding to the control group.
The metabolites concentration in the plasma, liver and muscle are represented on Table 1. After 96 h of exposure to the herbicide, meaningful metabolic alterations were observed in P. brachypomus. Levels of plasmatic glucose increased 16.9% on the exposed group, whereas muscular glycogen decreased 67.84%. Hepatic glycogen remained the same. Plasmatic protein decreased 20.4% in the exposed fish.
After acute exposure, the levels of GSH and GST activity were not significant compared to the control group. Likewise, increased of LPO did not change after exposure (Table 2).
| Metabolites | |||
|---|---|---|---|
| Variables | Tissue | Control | Norton (2,4D+Picloram) |
| Ammonia (mL-1 plasma) | Plasma | 0.45 ± 0.09 | 0.44 ± 0.07 |
| Glucose (mg dL-1) | Plasma | 147.90 ± 4.16 | 172.90 ± 7.56* |
| Glycogen (μmol of glycosyl-glucose g tissue-1) | Liver | 16.91 ± 8.09 | 29.47 ± 7.84 |
| Muscle | 175.90 ± 51.09 | 56.58 ± 18.75* | |
| Protein (mg g tissue-1 or mg mL plasma-1) | Plasma | 18.20 ± 1.18 | 14.50 ± 0.92* |
| Liver | 5.03 ± 0.32 | 4.711 ± 0.28 | |
| Muscle | 2.00 ± 0.13 | 2.04 ± 0.08 |
Table 1: Metabolites concentration on plasma, liver and muscle of Piaractus brachypomus exposed to Norton herbicide (2,4-D + Picl
- GSH-GST biotransformation system and lipid oxidative damage
- Variables
- Tissue
- Control
- Norton (2,4D+Picloram)
- Liver
- 528.0 ± 38.31
- 518.0 ± 38.83
- GST (U mg protein-1)
- Gills
- 13.10 ± 0.888
- 15.01 ± 1,.294
- Kidney
- 27.28 ± 2.756
- 29.44 ± 2.725
- Liver
- 4.860 ± 0.708
- 5.333 ± 0.952
- GSH (nmol GSH mg protein-1)
- Gills
- 31.04 ± 1.955
- 30.71 ± 2.233
- Kidney
- 14.66 ± 3.507
- 6.353 ± 1.358
- Liver
- 1.441 ± 0.315
- 1.242 ± 0.400
- LPO (nmol mg protein-1)
- Gills
- 0.949 ± 0.342
- 0.845 ± 0.156
- Kidney
- 1.806 ± 0.490
- 0.829 ± 0.199
Table 2: Activity of the antioxidant enzyme GST, glutathione levels (GSH) and lipid peroxidation of liver, gills and kidney of
Liver, Kidney, Gills Histopathology and Damage Indexes
All pathologies observed in the liver, gills and kidney of P. brachypomus are represented, respectively, in Figures 2 to 4. In liver, control group presented a characteristic and homogenous parenchyma, hepatocytes with polygonal contours, rounded nuclei and central nucleoli (Figure 2A). After exposure, it was observed damage such as alteration of hepatocyte architecture (Figure 2B), atrophy and nuclear alterations (Figure 2B,2D).
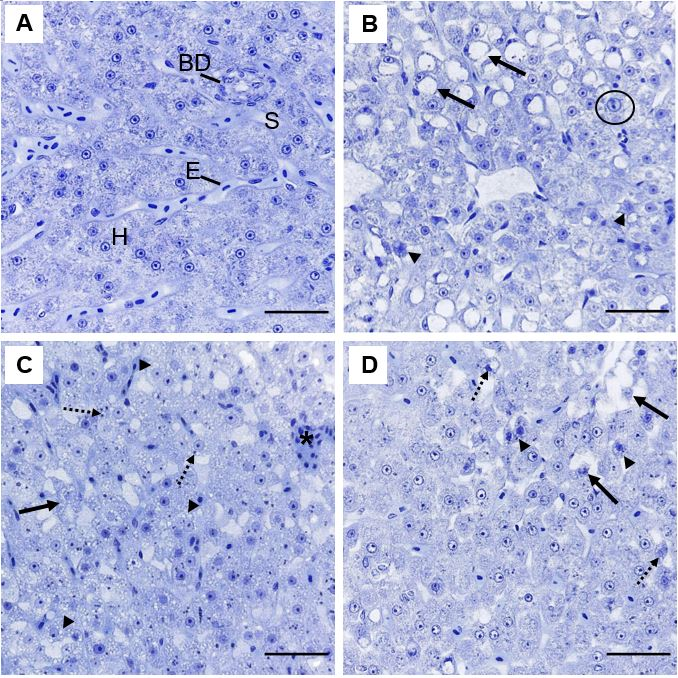
Figure 2: Representative histopathologies on liver of Piaractus brachypomus exposed to Norton herbicide (2,4-D + Picloram) for 96 h. (A) Control. (B-D) liver of fish exposed to the herbicide. Alteration of hepatic architecture with nuclear alteration (arrow), hepatocytes atrophy (arrowhead) and nuclear hypertrophy (circle) can be observed on (B). Cellular degeneration (arrow), cytoplasmatic inclusions (dashed arrow) and vacuolization (arrowhead) can be observed on (C). Nuclear alteration (dashed arrow), hepatocyte atrophy (arrowhead) and cellular degeneration (arrow) can be observed on (D). BD = Biliary duct; E = erythrocyte; H = hepatocyte and S = sinusoids. Scale bar = 20 µm.
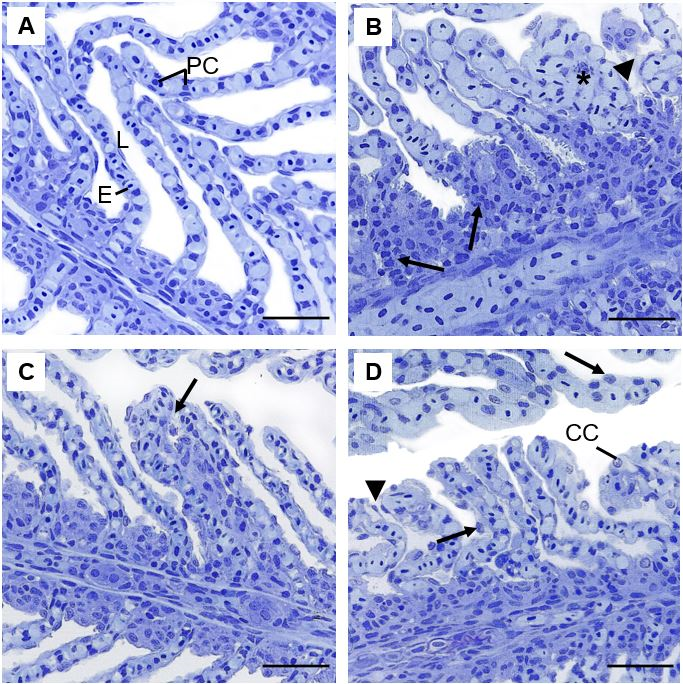
Figure 3: Representative histopathologies on gills of Piaractus brachypomus exposed to Norton herbicide (2,4-D + Picloram) for 96 h. (A) Control. (B-D) gills of fish exposed to the herbicide. Pillar cells constriction (arrow), lamellar congestion (*) and epithelium rupture (arrowhead) can be observed on (B); hyperplasia (arrow) can be observed on (C); epithelium displacement (arrowhead) and cellular hypertrophy (arrows) can be observed on (D). E = erythrocyte; L= lamellae; PC = Pillar cells; CC = Chloride cells. Scale bar = 20 µm.
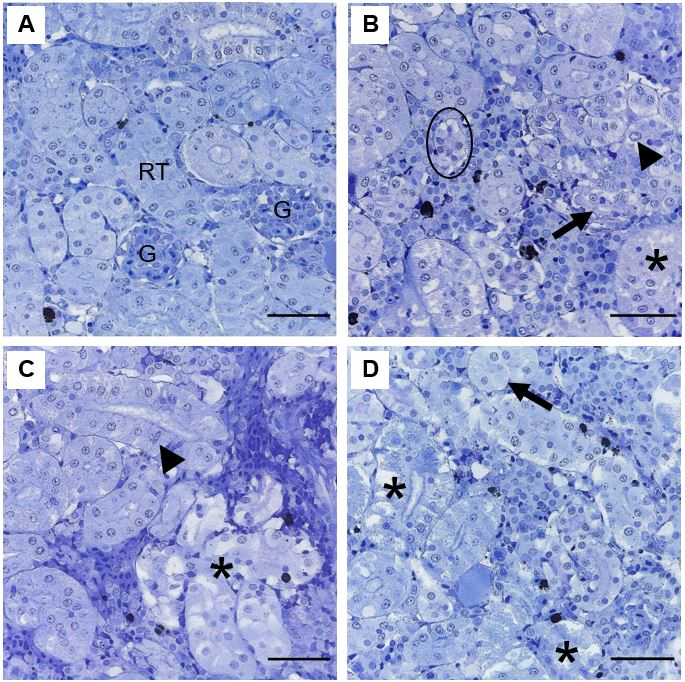
Figure 4: Representative histopathologies on kidney of Piaractus brachypomus exposed to Norton herbicide (2,4-D + Picloram) for 96 h. (A) Control. (B-D) Kidney of exposed fish. Nuclear alterations of tubular cell (arrowhead), tubule atrophy (circle), disarranged tubules (arrow); tubular degeneration (*) and tubule hypertrophy (dashed arrow) can be observed. G = glomerulus; RT = Renal tubules. Scale bar = 20 µm.
The general morphological characteristics of gills on P. brachypomus control group present lamellar pavement epithelium with no alterations, lamellae pillar cells, without obstruction of flow and circulatory disturbances (Figure 3A). After exposure there were morphological alterations in the gills, including constriction of pillar cells system and hypertrophy of lamellar epithelium (Figure 3B-D).
In the kidney, animals from the control group presented a well-defined space between glomerulus and Bowman´s capsule and tubules surrounded by interstitial, hematopoietic and lymphoid tissue (Figure 4A). Most observed modifications were renal architecture and cellular alteration (Figure 4B-4D).
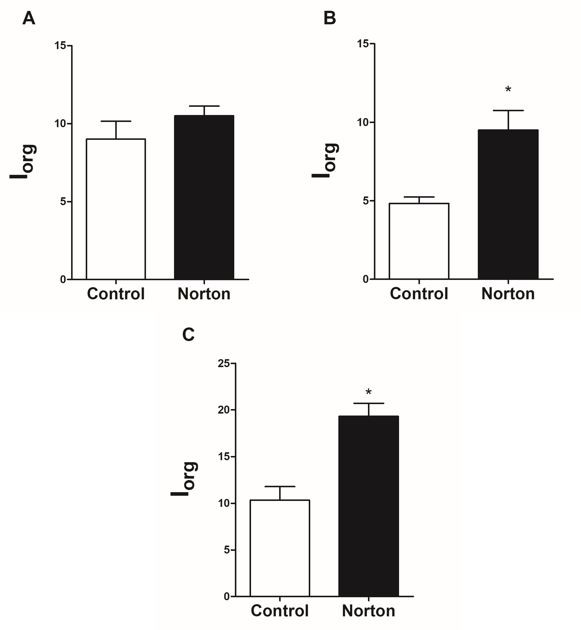
After acute exposure, hepatic, renal and gill exhibited changes. All pathologies observed and Ialt values have been describe in Table 3. Renal architecture parenchyma and cellular changes increased 150%, of which, cellular disorganization, degeneration and hypertrophy were most pronounced. In the liver, there was a significant increase in disorganization of the hepatic parenchyma (166%), nuclear changes (233%) and hepatocyte atrophy (506%). Gills Ialt shown increased of pillar cells constriction (240%) and hypertrophy of lamellar epithelium (77%).
After acute herbicide exposure, the Iorg of liver and gill of exposed groups increased significantly compared with control group (Figure 5), although some alteration was observed in kidney, the Iorg did not show renal tissue impairment.
| Individual Alteration Index (I ) alt | Exposure of 96 h | ||
|---|---|---|---|
| IF | Control | Norton (2,4D+Picloram) | |
| Renal Damage | |||
| Architecture and cellular alteration | 1 | 1.00 ± 0.25 | 2.50 ± 0.22* |
| Intracellular accumulation | 1 | 1.16 ± 0.21 | 1.33 ± 0.21 |
| Atrophy (tubules and glomerulus) | 2 | 1.00 ± 0.44 | 1.66 ± 0.33 |
| Nuclear alterations of renal epithelium | 2 | 2.66 ± 0.66 | 2.00 ± 0.00 |
| Alterations of interstitial tissue | 1 | 1.20 ± 0.20 | 1.20 ± 0.20 |
| Tubules hypertrophy | 1 | 1.33 ± 0.21 | 1.16 ± 0.21 |
| Glomerulus hypertrophy | 1 | 0 | 0.16 ± 0.16 |
| Hepatic Damage | |||
| Architecture and cellular alteration | 2 | 0.50 ± 0.22 | 1.33 ± 0.21* |
| Nuclear alterations | 1 | 1.00 ± 0.44 | 3.33 ± 0.84* |
| Cellular atrophy | 2 | 0.66 ± 0.42 | 4.00 ± 0.89* |
| Intracellular accumulation | 1 | 1.00 ± 0.25 | 0.50 ± 0.22 |
| Cytoplasmic degeneration | 1 | 0.66 ± 0.21 | 0.666± 0.21 |
| Hyperemia (sinusoids) | 1 | 0.33 ± 0.21 | 0.16 ± 0.16 |
| Hepatocyte hypertrophy | 1 | 0.33 ± 0.21 | 0.16 ± 0.16 |
| Gills Damage | |||
| Pillar cells (Constriction) | 1 | 0.83 ± 0.16 | 2.83 ± 0.40* |
| Hypertrophy of lamellar epithelium | 1 | 2.16 ± 0.54 | 3.83 ± 0.16* |
| Congestion/Hyperemia/Hemorrhage | 1 | 0.50 ± 0.22 | 0.66 ± 0.21 |
| Epithelium hyperplasia | 1 | 1.00 ± 0.44 | 2.33 ± 0.33 |
| Lamellar epithelium detachment and edema | 1 | 2.50 ± 0.42 | 3.50 ± 0.22 |
| Partial lamellar fusion | 1 | 0 | 0.50 ± 0.22 |
| Proliferation of chloride cells on lamellae | 1 | 1.00 ± 0.25 | 1.16 ± 0.40 |
| Epithelium rupture | 2 | 2.33 ± 0.33 | 4.16 ± 0.74 |
Table 3: Individual alteration index (lalt) of histopathologies of kidney, liver and gills of Piaractus brachypomus exposed to th
Discussion
The presence of 2,4-D + picloram-based herbicide in the water demonstrated toxicity for P. brachypomus. Our results showed that the herbicide at sublethal concentration may induced metabolic adjustments on the organisms which are adaptive strategies to maintain cellular function given the conditions.
Aminotransferases are fundamental for amino acids and proteins metabolism, and are released on blood stream after hepatic damage [26]. In fact, plasmatic analysis of transaminases concomitant with acid and alkaline phosphatases is necessary to evaluate hepatic damage on sublethal exposure [27]. In this study, it was not demonstrated alterations on the plasma activity of both aminotransferases, suggesting absence of cell membrane injuries and cell ruptures. According to Gallagher E, et al. [5], catfish (Ictalurus punctatus) exposed to the mixture of 2,4-D + Picloram demonstrated a higher metabolic rate and the normal levels of ALT, also indicating that injuries were not observed. These observations do not imply that the herbicide does not induce changes. After exposure to the herbicide, there was a decrease in ALP activity, which could be related to problems on transmembrane transportation through the hepatocyte membrane, whereas this enzyme has an important role on transportation of phosphorylated intermediates [10].
Carbohydrates are usually a primary source of energy on stressing conditions. Exposure to pesticides can cause reduction on both muscular and hepatic glycogen levels [28]. In this study, P. brachypomus presented a depletion of glycogen storage on muscle followed by increase of plasmatic glucose that was used as an energy source to physiological stress. Muscular glycogen degradation may be associated with increase of acetylcholine levels on muscle, caused by acetylcholinesterase inhibition Sastry K, et al. [29], because catecholamine’s secretion caused by this process promotes an increase on glycogenolysis to attend the muscular demand for ATP required by the increase on locomotor activity [30]. Therefore, previous works evidenced that the Norton herbicide based on 2,4-D + picloram was capable of creating locomotor disturbances and inhibit AChE activity on muscle of P. brachypomus after exposure to a sub-lethal concentration (unpublished data).
However, the need for glucose in fish can be answered with hydrolysis of hepatic glycogen and precursors such as lactate amino acids and pyruvate (gluconeogenesis) [31]. It is necessary to highlight that the degradation of muscular glycogen in this present study did not contribute directly to the increase of plasmatic glucose levels. According to Begg K, et al. [32], lactate (by-product of muscular glycogen) can act as a substrate on glucose synthesis and also contribute to preserve glycogen storage on liver. Similar responses were observed on Cyprinus carpio exposed to 2,4 diamine (derived from 2,4-D) that suggests a mechanism of adjustment to supply the energetic demand, caused by a hypoxic state created by the compound [33].
Use of biomarkers of biotransformation and cellular damage are part of the methods broadly used to evaluate the effects of pollutants over fish [34]. GST-GSH system acts on phase II of detoxification, conjugating metabolites produced by phase I of biotransformation. After conjugation of toxic compounds, the product created is capable of being excreted of the cell without causing potential cellular damage, such as lipid peroxidation [35]. In this present work, there were no alterations of the GST-GSH system on gills, liver and kidney. This suggests that the compound did not affect the detoxification capacity of the cellular antioxidant system, and with that, it was possible to contain the effects of oxidative stress. Similar results were observed by Gallagher E, et al. [5] on liver of I. punctatus exposed to a mixture of 2,4-D and picloram that did not present changes on GST activity.
In accordance with the results presented on the GST-GSH system in this study, unaltered levels of LPO on the analyzed organs of P. brachypomus indicate that the oxidants did not promote a significant decrease of membrane phospholipids that would cause severe damage on cells properties [36]. However, Salla BF, et al. [9] observed that the interactions of 2,4-D and picloram with the bilayer membrane of hepatocytes of Wistars rats can be highly destructive and can be related to cellular toxicity of these active principles. Nevertheless, it is important to highlight that the differences of biological responses towards herbicides occur by many factors, such as exposure route, formulation and species [37].
Morphological changes of fish after the exposure from the herbicide action over the tissues. Therefore, alteration of hepatic architecture can indicate modifications of the cytoskeleton, that creates disorganization on the organelle distribution and increase in intercellular space which may influence on the interaction between hepatocytes [38]. Hepatocyte atrophy on P. brachypomus after exposure to Norton herbicide comprehends an adaptation mechanism. According to Safahieh A, et al. [39], atrophy is a response to a decrease of specialized functions in cells. However, this pathology persistence can evolve to usually irreversible damage, such as apoptotic and oncotic necrosis [40].
In this present study, the majority of nuclear alterations were unspecific. Hypertrophy of nuclei suggests increase in cellular activity percentage whereas the cytoplasmic vacuolization may indicate a possible stage of initial degeneration [41]. These changes on hepatic tissue were reported on Poecilia vivípara, caused by 2,4-D on commercial formulation after sublethal exposure [42]. In this sense, hepatic lorg calculated for P. brachypomus suggests that the impairment of liver tissue caused by individual alterations compromise of the tissue and its function.
Alterations on gill structure can interfere in the proper operation of the organ, and generally, are caused by biochemical and physiological changes [43]. Some pathologies were significantly more frequent on exposed animals, such as hypertrophy of cells of the lamellar epithelium. These alterations are defense mechanisms that increase the distance between water and blood and reduce the interlamellar space to avoid absorption of toxic substances. However, it may occur a decrease of area of respiratory surface during this process, diminishing gas exchange and leading to osmoregulatory dysfunction [13]. These pathologies were observed on Clarias gariepinus exposed to 2,4-D amine Makinde GEO, et al. [44] and on Ctenopharyngodon idella exposed to Picloram [45]. Constriction of pillar cells system caused by exposure of P. brachypomus to the Norton herbicide resulted on circulatory disturbances. These alterations also interfere on gas exchange and can create lamellar epithelium detachment [46]. Considering the gill alterations, lorg indicated that the lesions presented on branchial tissue of P. brachypomus generated organ dysfunctions after 96 hours of exposure.
In fish, the kidney is an organ that is frequently associated with morphological alterations towards stressors [47]. In this study, the alterations of renal architecture and cellular structure observed on kidney of P. brachypomus are due to the degenerated and disorganized aspect of the tubular cells. These conditions may be related with the initial stage of the granular degenerative process that exhibits undefined cellular contour, increase on volume of the epithelial tubular cells and decrease of lumen caliber. These alterations may impair passage of the filtrate and compromise the process of reabsorption and tubular secretion Camargo MMP, et al. [48]; however, it is considered reversible [49, 50]. According to Gallagher E, et al. [5], combination of 2,4-D and picloram created damage of proximal tubules in the kidney of I. punctatus exposed to sublethal concentrations, indicating that the combination of active principles caused toxic effects on the tissue.
In this present study, lorg indicated that the total alterations presented did not compromise organ function. The kidneys have a fundamental role on xenobiotics excretion, as well as ionic and hydric balance, eliminating chemical compounds during the formation of urine, through the glomerular filtration, reabsorption or tubular secretion [47]. Decrease of plasmatic protein on P. brachypomus after the period of sublethal exposure may be related to the process of protein elimination on urine (proteinuria), which indicates that the observed morphological alterations (lalt) might have been caused harm even without compromising total kidney function (lorg). In embryos of Danio rerio exposed to the commercial herbicide Tordon (2,4-D + Picloram) was observed significate decrease of the total protein amount (body), it being an effect attributed to proteinuria [51].
Conclusion
The exposure to the Norton herbicide (2,4-D + Picloram) for 96 hours at sublethal concentration for P. brachypomus resulted on morphophysiological changes. Despite the complexity of the interaction between 2,4-D and Picloran of synergism effect cannot be excluded when commercial formulations are used on agricultural cultures. Our results showed absence of oxidative stress on tissues and severe injuries at acute exposure, however, histopathologies in the liver and gills suggest that most of them were induced by herbicide. Although Ialt showed specific and individual injuries, the Iorg suggest liver and gills dysfunction. Considering that concentration used was sublethal and environmentally relevant, 2,4-D and picloran associated may be absorbed by fish. P. brachypomus demonstrated ability to active metabolism rout to meet energy demands in stressful conditions, and, the integrated biomarkers were useful tools to evaluated to potential toxicity of herbicides.
References
-
Rigotto RM, Vasconcelos DP, Rocha MM (2014) Pesticide use in Brazil and problems for public health. Cad Saúde Pub 30(7): 1360-1362.
-
Caldeira DSA, Amaral VN, Casadei RA, Barros LV, Figueiredo ZN (2014) Controle de plantas daninhas em pastagem usando doses e misturas de herbicidas. Encicl Biosf 10: 1052-1060.
-
Donald DB, Cessna AJ, Sverko E, Glozier NE (2007) Pesticides in surface drinking-water supplies of the Northern Great plains. Environ Health Perspect 115(8): 1183-1190.
-
Blakley PM, Kim JS, Firneisz GD (1989) Effects of paternal subacute exposure to tordon 202c on fetal growth and development in CD-1 mice. Teratol 39: 237-241.
-
Gallagher E, Digiulio R (1991) Effects of 2, 4-dichlorophenoxyacetic acid and picloram on biotransformation, peroxisomal and serum enzyme activities in channel catfish (_Ictalurus punctatus_). Toxicol Lett 57(1): 65-72.
-
Oakes D, Pollak J (1999) Effects of a herbicide formulation, Tordon 75D®, and its individual components on the oxidative functions of mitochondria. Toxicol 136(1): 41- 52.
-
Leão MB, Gonçalves DF, Miranda GM, da Paixão GMX, Dall Corte CL (2019) Toxicological evaluation of the herbicide Palace® in Drosophila melanogaster. J Toxicol. Environ Health Part A 82(22): 1172-1185.
-
Tavares D, Paulino MG, Terezan AP, Fernandes JB, Giani A, et al. (2018) Biochemical and morphological biomarkers of the liver damage in the Neotropical fish, Piaractus mesopotamicus, injected with crude extract of cyanobacterium Radiocystis fernandoi. Environ Sci Pollut Res 25(16): 15349-15356.
-
Salla BF, Bracht L, Valderrama Parizotto A, Comar JF, Peralta RM, et al. (2019) Kinetics of the metabolic effects, distribution spaces and lipid-bilayer affinities of the organo-chlorinated herbicides 2,4-D and picloram in the liver. Toxicol Lette 313: 137-149.
-
Bernet D, Schmidt H, Wahli T, Burkhardt-Holm P (2001) Effluent from a sewage treatment works causes changes in serum chemistry of brown trout (_Salmo trutta L_.). Ecotoxicol Environ Saf 48(2): 140-147.
-
Ozer J, Ratner M, Shaw M, Bailey W, Schomaker S (2008) The current state of serum biomarkers of hepatotoxicity. Toxicol 245(3): 194-205.
-
Moron SE, Oba ET, De Andrade CA, Fernandes MN (2003) Chloride cell responses to ion challenge in two tropical freshwater fish, the erythrinids Hoplias malabaricus and Hoplerythrinus unitaeniatus. J of Experim Zoo 298(2): 93-104.
-
Sakuragui MM, Sanches JR, Fernandes MN (2003) Gill chloride cell proliferation and respiratory responses to hypoxia of the neotropical erythrinid fish Hoplias malabaricus. J Comp Physiol 173(4): 309-317.
-
Evans DH (1993) The Physiology of Fish. CRC Press, United States, pp: 592.
-
Husak VV, Mosiichuk NM, Maksymiv IV, Sluchyk IY, Storey JM, et al. (2014) Histopathological and biochemical changes in goldfish kidney due to exposure to the herbicide Sencor may be related to induction of oxidative stress. Aquat Toxicol 155: 181-189.
-
Badroo IA, Nandurkar HP, Khanday AH (2020) Toxicological impacts of herbicide paraquat dichloride on histological profile (gills, liver, and kidney) of freshwater fish Channa punctatus (Bloch). Environ Sci Pollut Res 27(31): 39054-39067.
-
Van der Oos R, Beyer J, Vermeulen PE (2003) Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ Toxicol Pharmacol 13(2): 57-149.
-
Sakuragui MM, Paulino MG, Souza NES, Tavares D, Terezan AP, et al. (2019) Crude extract of cyanobacterium Radiocystis fernandoi strain R28 induces anemia and oxidative stress in fish erythrocytes. Toxicon 169: 18-24.
-
Paulino MG, Tavares D, Terezan AP, Sakuragui MM, Pesenti E, et al. (2020) Biotransformations, antioxidant system responses and histopathological indexes in the liver of fish exposed to cyanobacterial extract. Environ. Toxic and Chem 39(5): 1041-1051.
-
Gentzkow CJ, Masen JM (1942) An accurate method for the determination of blood urea nitrogen by direct nesslerization. J Biol Chem 143(2): 531-544.
-
Bidinotto PM, Moraes G, Souza RHS (1997) Hepatic glycogen and glucose in eight tropical fresh water fish: a procedure for field determination of micro samples. Bol Tec CEPTA 10: 53-60.
-
Kruger NJ (1994) The Bradford method for protein quantification. Methods Mol Biol 32: 9-15.
-
Habig WH, Jakoby WB (1981) Assays for differentiation of glutathione Stransferases. Methods Enzymol 77: 398- 405.
-
White CC, Viernes H, Krejsa CM, Botta D, Kavanagh TJ (2003) Fluorescence‐ based microtiter plate assay for glutamate‐cysteine ligase activity. Anal Biochem 318(2): 175-180.
-
Jiang Z, Ahn DU, Ladner L, Sim JS (1992) Influence of feeding full-fat flax and sunflower seeds on internal and sensory qualities of eggs. Poult Sci 71(2): 378-382.
-
Ezenwaji NE, Bebe Y, Nwaigwe C (2013) Changes in liver and plasma enzymes of Clarias gariepinus exposed to sublethal concentrations of diesel. Afr J Biotechnol 12: 414-418.
-
Fathy M, Mohamed IA, Farghal AIA, Temerak SAH, Sayed AEDH (2019) Hemotoxic effects of some herbicides on juvenile of Nile tilapia Oreochromis niloticus. Environ Sci and Pollu Research 26(30): 30857-30865.
-
Hori TSF, Avilez IM, Inoue LK, Moraes G (2006) Metabolical changes induced by chronic phenol exposure in matrinxã Brycon cephalus (teleotei: characidae) juveniles. Comp Biochem Phisiol 143(1): 67-72.
-
Sastry K, Siddiqui A (1982) Chronic toxic effects of the carbamate pesticide sevin on carbohydrate metabolism in a freshwater snakehead fish, Channa punctatus. Toxicol Lett 14(2): 123-130.
-
Martinez-Porchas M, Martinez-Cordova LR, Ramos- Enriquez R (2009) Cortisol and glucose: reliable indicators of stress?. Pan Am J Aquat Sci 4: 158-178.
-
Li H, Xu W, Jin J, Yang Y, Zhu X, et al. (2018) Effects of starvation on glucose and lipid metabolism in gibel carp (Carassius auratus gibel var. CAS III). Aqua 496: 166-175.
-
Begg K, Pankhurst NW (2004) Endocrine and metabolic responses to stress in a laboratory population of the tropical damselfish Acanthochromis polyacanthus. J Fish Biol 64(1): 133-145.
-
Oruç EO, Üner N (1999) Effects of 2,4-Diamin on some parameters of protein and carbohydrate metabolisms in the serum, muscle and liver of Cyprinus carpio. Environ Pollut 105(2): 267-272.
-
Bonomo MM, de Castro Sachi IT, Paulino MG, Fernandes JB, Carlos RM, et al. (2021) Multi-biomarkers approach to access the impact of novel metal-insecticide based on flavonoid hesperidin on fish. Environ Poll 268(2): 115758.
-
Huang X, Li Y, Wang T, Liu H, Shi J, et al. (2020) Evaluation of the Oxidative Stress Status in Zebrafish (Danio rerio) Liver Induced by Three Typical Organic UV Filters (BP-4, PABA and PBSA). Internat J Environ Res and Pub Health 17(2): 651.
-
Srivastava B, Reddy PB (2017) Lipid peroxidation and DNA damage as biomarkers of pollution induced oxidative stress in fish. Life Sci Int J 4: 194-198.
-
Islam E, Khan MT, Irem S (2015) Biochemical mechanisms of signaling: perspectives in plants under arsenic stress. Ecotoxicol Environ Saf 114: 126-133.
-
Zhao Y, Li W, Chou L (1987) Cytoskeletal perturbation induced by herbicides, 2,4‐dichlorophenoxyacetic acid (2,4‐d) and 2,4,5‐trichlorophenoxyacetic acid (2,4,5‐t). J of Toxicol and Environ Health 20(1): 11-26.
-
Safahieh A, Hedayati A, Savari A, Movahedinia A (2011) Effect of sublethal dose of mercury toxicity on liver cells and tissue of yellowfin seabream. Toxicol Ind Health 28(7): 583-592.
-
Hinton DE, Segner H, Braunbeck T (2001) Toxic responses of the liver. In: Schlenk D, Bensen WH (Eds) Toxicity in Marine and Freshwater Teleosts, London: Taylor and Francis 1: 224-268.
-
Takashima F, Hibiya T (1995) An Atlas of Fish Hidtology. Normal and Pathological Features. Gustav Fisher Verlag, pp: 195.
-
Vigário AF, Sabóia-Morais SMT (2014) Effects of the 2,4- D herbicide on gills epithelia and liver of the fish Poecilia vivipara. Pesq Vet Bras 34: 523-528.
-
Fernandes MN, Mazon AF (2003) Environmental pollution and fish gill morphology. In: Val AL, Kapoor BG (Eds.), In Fish Adaptations. Sci Publi Inc 422: 203-231.
-
Makinde GEO, Olaifa FE, Bajo OT (2015) Acute toxicity and histopathological Changes in gill and liver of catfish (Clariasgaripinus) exposed to 2, 4- D Amine (Herbex D SI). J of Bio Agric and Heal 5(4): 145-149.
-
Botelho RG, Santos JB, Fernandes KM, Neves CA (2012) Effects of atrazine and picloram on grass carp: acute toxicity and histological assessment. Toxicol Environ Chem 94(1): 121-127.
-
Sachi ITC, Bonomo MM, Sakuragui MM, Modena PZ, Paulino MG, et al. (2021) Biochemical and morphological biomarker responses in the gills of a Neotropical fish exposed to a new flavonoid metal-insecticide. Ecotoxicol and Environ Safety 208: 111459.
-
Carmo TLL, Azevedo VC, Siqueira PR, Galvão TD, Santos FA, et al. (2018) Reactive oxygen species and other biochemical and morphological biomarkers in the gills and kidneys of the Neotropical freshwater fish, Prochilodus lineatus, exposed to titanium dioxide (TiO2) nanoparticles. Environ Sci and Poll Res 25(23): 22963- 22976.
-
Camargo MMP, Martinez CBR (2007) Histopathology of gills, kidney and liver of a Neotropical fish caged in an urban stream. Neotrop Ichthyol 5(3): 327-336.
-
Hinton DE, Lauren DJ (1990) Integrative histopathological approaches to detecting effects of environmental stressors on fishes. Am Fish Soc Symp 8: 51-66.
-
Silva AG, Martinez CBR (2007) Morphological changes in the kidney of a fish living in an urban stream. Environ Toxicol Pharmacol 23(2): 185-192.
-
Gomes DSF (2017) Effects of 2,4-dichlorophenoxyacetic acid and Tordon® on embryonic development and energy metabolism in zebrafish (danio rerio). Dissertation (Master in Bioactive Products and Biosciences). Uni Federal of Rio de Janeiro, pp: 81.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh