Circadian Rhythm of the Phytoplankton Assemblage during a Spawning Season of Sardinops sagax caerulea in Bahía Magdalena, Baja California Sur, Mexico
The Magdalena-Almejas Lagoon System is considered an area of interest both from biological and fishing points of view. This study deals with the variation of phytoplankton assemblages and total and partial cell abundance in three circadian cycles during the winter of 1989. The results obtained showed that there are short timescale changes (circadian rhythms) in water temperature, species composition, total and partial abundance, resulting from the tidal phase. Diatoms and dinoflagellates were the most abundant taxonomic groups. In total, 99 species and infraspecific taxa from 60 genera were found: 79 diatoms, 14 dinoflagellates, 2 cyanobacteria, 2 silicoflagellates, an ebridean and an euglenophyte. Proliferation of some diatom species, such as Eucampia zodiacus, Chaetoceros compressus, C. curvisetus, Coscinodiscus radiatus, Detonula pumila and the dinoflagellate Prorocentrum koreanum, was responsible for the maximal phytoplankton abundances, the alternating dominance of size fractions and the low species diversity. The remaining groups (cyanobacteria, silicoflagellates, ebrideans and euglenophytes) were not well represented either in number of species or abundance. In general, the high availability of food (phytoplankton) for sardine larvae during their reproductive period was noted; therefore, food is not a limiting factor for their development.
Introduction
Lagoon environments are coastal ecotones, partially or permanently connected to the sea, where 25% of global primary production takes place [1]. The interest in their studies originates from the use of these ecotones as nursery grounds by a great variety of marine fish and some crustaceans of commercial importance. They are optimal areas for mariculture, as well as for minor pelagic fisheries, such as sardine, anchovy and related species [2]. The Magdalena-Almejas Lagoon System is one of the most important coastal lagoon complexes in Baja California Sur, NW Mexico [3, 4], as it is one of the few areas where commercial sardine catches of the Monterey sardine (South American pilchard, Sardinops sagax caerulea (Teleostei: Alosidae) fishery occur [5]. In Bahía Magdalena, the spawning period lasts from January to May, with a maximum in February-March [6]. The success of sardine and anchovy fisheries in upwelling regions was initially attributed to their ability to feed directly upon phytoplankton [7]. According to McFarlane, et al. [8], sardines are opportunistic feeders with dominant taxonomic groups in their diet reflecting abundance and availability of their prey of euphausiids (and their eggs), copepods and diatoms.
Circadian rhythms drive phytoplankton photosynthesis and zooplankton migration, while circalunar rhythms link zooplankton depth to moonlight. Circadian rhythms also govern the activities of phyto- and zooplankton, ensuring a balanced ecological order [9]. Among the main studies on spatio-temporal variations in phytoplankton in the Magdalena-Almejas Lagoon System are those of Nienhuis, et al. [10] which related the distribution and abundance of phytoplankton to eutrophic waters. Gárate- Lizárraga and Siqueiros-Beltrones [11] reported that phytoplankton abundance peaks in spring and autumn, with the microphytoplankton fraction (20-200 μm) being dominant. In contrast, Gárate-Lizárraga, et al. [12] found the nanophytoplanktonic fraction (2-20 μm) to be the most important, especially under oligotrophic conditions. Regarding diurnal variations, Acosta-Ruíz, et al. [13] reported that salinity, water temperature and chlorophyll-a concentration were strongly correlated with the tidal cycle. Other phytoplankton studies have diversified to include those related to algal blooms [4, 14, 15]. Cervantes-Duarte, et al. [16] determined the variation in phytoplankton primary productivity from spring 2013 to summer 2014, noting the cold months as the most productive due to the contribution of eutrophic waters by upwelling. Recently, Cervantes-Duarte, et al. [17] determined water quality and trophic status from 2015 to 2018 in Bahía Magdalena, finding annual and interannual changes in hydrobiological structure. In 2015, they recorded the minimum values of the Trophic State Index (TRIX), attributing them to the effects of warming events such as El Niño and The Blob.
The present study aimed to determine the circadian variation in the phytoplankton species composition and abundance during three 24-hour cycles (February 1989) at the entrance of Bahía Magdalena during the spawning season of Sardinops sagax caerulea.
Material and Methods
Study Area
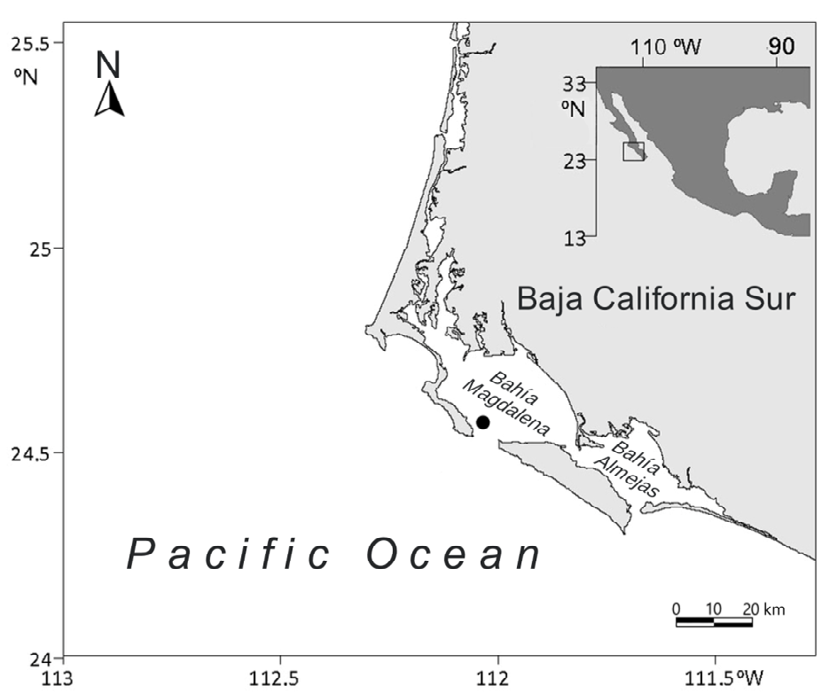
The Magdalena-Almejas Lagoon System is located on the western coast of the Baja California Sur Peninsula (Figure 1) between 24°15’ N and 25°20’ N, and 111°30’ W and 112°12’ W [18]. It has an average annual precipitation of 14 mm, with low or absent freshwater input and a high degree of evaporation [18], and it is thus considered an anti-estuarine system [13]. Nutrient exchange between the lagoon system and the ocean takes place through the main mouth of the bay, and it is defined by the pattern of tidal currents, so that maximum concentrations occur during the flow and decrease during the ebb [19].
Sampling
Three 24-hour samplings were conducted at a station near the mouth of the Bahía Magdalena, located at Colnett Point; site depth ca. 40 m (Figure 1). The period of sampling was chosen based on the highest primary productivity and a pronounced upwelling in the cold months, as previously mentioned. The first cycle was from February 9 to 10, 1989 (cycle 1), the second from February 16 to 17, 1989 (cycle 2) and the third from February 23 to 24, 1989 (cycle 3). Phytoplankton samples (250 ml) were taken every hour at the surface and at a depth of 5 m with a Van Dorn bottle and preserved in 3% buffered formalin, which permitted the adequate counting of micro- and nanoplanktonic microalgae. The frequency of sampling was chosen considering the highly dynamic study area. Furthermore, it is known that in cultures of marine phytoplankton under optimal conditions, the maximum rates can reach 4-6 divisions per day [20]. It was also concluded that diatoms, in general, tend to divide faster than dinoflagellates, and “a 24-hour lifetime can retain as an average or an order of magnitude” [20].
Hydrological Measurements, Microalgal Counting and Identification
Water temperature was also recorded at both levels using a bucket thermometer. Upwelling index values from the adjacent oceanic zone were provided by the Pacific Fisheries Environmental Group of NOAA, Monterey, CA, USA [21]. Samples for phytoplankton analysis were settled in 5-mL Utermöhl sedimentation chambers and observed under a phase-contrast inverted Carl Zeiss microscope. The total phytoplankton, nanoplankton and microplankton abundances (cells L-1) were estimated simultaneously with the species identifications of microplankton [22, 23].
Specialized taxonomic keys for each group were used to identify microphytoplankton species [24]. The nomenclature of phytoplankton species was updated using the database of Guiry & Guiry [25]. Tychoplanktonic (benthic) species (only diatoms) were also included (Table 1). AlgaeBase was consulted to verify currently accepted taxonomic names [25]. Abbreviations of authors of scientific names are used according to Brummit, et al. [26] unless they were not listed in the cited book. The Shannon-Wiener H’ diversity index was applied for data processing. Nanophytoplankton was counted; however, the cells were not identified.
| DIATOMS | 56 | bNitzschia spathulata Bréb. ex W.Sm. | |
|---|---|---|---|
| 1 | bActinoptychus senarius (Ehrenb.) Ehrenb. | 57 | bpOdontella aurita (Lyngb.) C.Agardh |
| 2 | bActinoptychus vulgaris Schumann | 58 | bParalia fenestrata Sawai et Nagumo |
| 3 | bAmphora marina W.Sm. | 59 | Planktoniella sol (G.C.Wall.) F.Schütt |
| 4 | bAmphora sp. | 60 | bPleurosigma sp. |
| 5 | bArdissonea robusta (Ralfs) De Not. | 61 | Proboscia alata (Brightw.) Sundström |
| 6 | Asterionellopsis glacialis (Castrac.) Round | 62 | bPsammodictyon panduriforme (W.Greg.) D.G.Mann |
| 7 | bAsteromphalus flabellatus (Bréb.) Grev. | 63 | Pseudo-nitzschia pseudodelicatissima (Hasle) Hasle |
| 8 | bAsteromphalus heptactis (Bréb.) Ralfs | 64 | Pseudo-nitzschia sp. |
| 9 | Bacteriastrum delicatulum Cleve | 65 | Pseudosolenia calcar-avis (Schultze) Sundström |
| 10 | bCampylodiscus neofastuosus Ruck et Nakov | 66 | Rhizosolenia bergoni H.Perag. |
| 11 | Cerataulina pelagica (Cleve) Hendey | 67 | Rhizosolenia hyalina Ostenf. |
| 12 | Chaetoceros affinis Lauder | 68 | Rhizosolenia imbricata Brightw. |
| 13 | Chaetoceros atlanticus Cleve | 69 | Skeletonema costatum (Grev.) Cleve |
| 14 | Chaetoceros brevis F.Schütt | 70 | Stephanopyxis palmeriana (Grev.) Grunow |
| 15 | Chaetoceros compressus Lauder | 71 | Sundstroemia setigera (Brightw.) Medlin |
| 16 | Chaetoceros costatus Pavill. | 72 | bSynedra sp. |
| 17 | Chaetoceros curvisetus Cleve | 73 | Thalassionema nitzschioides (Grunow) Mereschk. |
| 18 | Chaetoceros danicus Cleve | 74 | Thalassiosira eccentrica (Ehrenb.) Cleve |
| 19 | Chaetoceros debilis Cleve | 75 | Thalassiosira gravida Cleve |
| 20 | Chaetoceros didymus Ehrenb. | 76 | Thalassiosira sp. |
| 21 | Chaetoceros eibenii Grunow | 77 | Thalassiothrix heteromorpha G.Karst. |
| 22 | Chaetoceros lorenzianus Grunow | var. mediterranea (Pavill.) Hallegr. | |
| 23 | Chaetoceros messanensis Castrac. | 78 | bTrachyneis aspera (Ehrenb.) Cleve |
| 24 | Chaetoceros peruvianus Brightw. | 79 | bpTrieres mobiliensis (Bailey) Ashworth et E.C.Theriot |
| 25 | Chaetoceros sp. | ||
| 26 | Chaetoceros lauderi Ralfs ex Lauder | DINOFLAGELLATES | |
| 27 | Chaetoceros coarctatus Lauder | 80 | Amphidinium sphenoides Wulff |
| 28 | Coscinodiscus asteromphalus Ehrenb. | 81 | Dinophysis hastata F.Stein |
| 29 | Coscinodiscus curvatulus Grunow | 82 | Gyrodinium cf. herbaceum Kof. et Swezy |
| 30 | Coscinodiscus perforatus Ehrenb. | 83 | Oxytoxum variabile J.Schiller |
| 31 | Coscinodiscus radiatus Ehrenb. | 84 | Phalacroma oxytoxoides (Kof.) F.Gómez, P.López-García et D.Moreira |
| 32 | bpCylindrotheca closterium (Ehrenb.) Reimann et J.C.Lewin | 85 | Pronoctiluca pelagica Fabre-Domergue |
| 33 | Dactyliosolen fragilissimus (Bergon) Hasle | 86 | Prorocentrum koreanum M.-S.Han, S.Y.Cho et P.Wang |
| 34 | Dactyliosolen mediterraneus (H.Perag.) H.Perag. | 87 | Protoperidinium conicum (Gran) Balech |
| 35 | Dactyliosolen phuketensis (Sundström) Hasle | 88 | Protoperidinium pellucidum Bergh |
| 36 | Detonula pumila (Castrac.) Gran | 89 | Scrippsiella acuminata (Ehrenb.) Kretschmann, Elbr., Zinssmeister |
| 37 | Ditylum brigthwellii (T.West) Grunow | S.Soehner, Kirsch, Kusber et Gottschling | |
| 38 | Eucampia zodiacus Ehrenb. | 90 | Tripos balechii (Meave, Okolodkov et M.E.Zamudio) F.Gómez |
| 39 | bFragilariopsis doliolus (G.C.Wall.) Medlin et P.A.Sims | 91 | Tripos furca (Ehrenb.) F.Gómez |
| 40 | Gossleriella tropica F.Schütt | 92 | Tripos fusus (Ehrenb.) F.Gómez |
| 41 | bGrammatophora marina (Lyngb.) Kütz. | 93 | Tripos trichoceros (Ehrenb.) F.Gómez |
| 42 | Guinardia flaccida (Castrac.) H. Perag. | ||
| 43 | Guinardia striata (Stolterf.) Hasle | CYANOBACTERIA | |
| 44 | bGyrosigma balticum (Ehrenb.) Rabenh. | 94 | Trichodesmium erythraeum Ehrenb. ex Gomont |
| 45 | bGyrosigma fasciola (Ehrenb.) J.W.Griff. et Henfrey | 95 | Trichodesmium sp. |
| 46 | bHalamphora terroris (Ehrenb.) P. Wang | ||
| 47 | Hemiaulus sinensis Grev. | SILICOFLAGELLATES | |
| 48 | Leptocylindrus danicus Cleve | 96 | Dictyocha messanensis Haeckel |
| 49 | bLicmophora abbreviata C.Agardh | 97 | Octactis pulchra J.Schiller |
| 50 | bLicmophora sp. | ||
| 51 | Meuniera membranacea (Cleve) P.C.Silva | EBRIDEANS | |
| 52 | bNavicula cancellata Donkin | 98 | Ebria tripartita (Shumann) Lemmerm. |
| 53 | bNavicula distans (W.Sm.) Bréb. | ||
| 54 | bNavicula zostereti Grunow | EUGLENOPHYTES | |
| 55 | bNavicula sp. | 99 | Eutreptia sp. |
Table 1: A list of microphytoplanktonic species found during the second and the third circadian samplings in Bahía Magdalena in F
Results
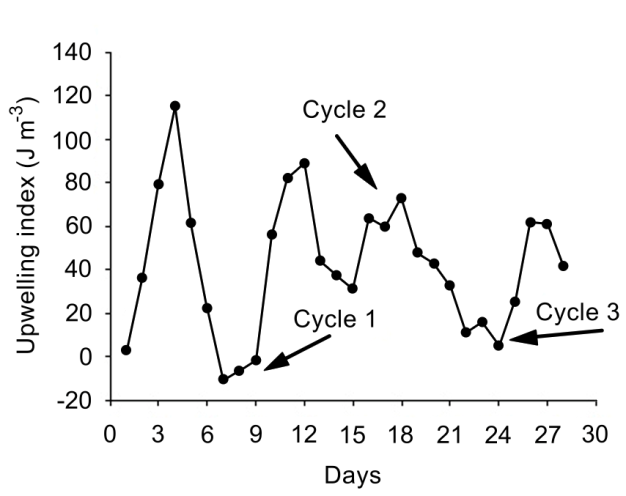
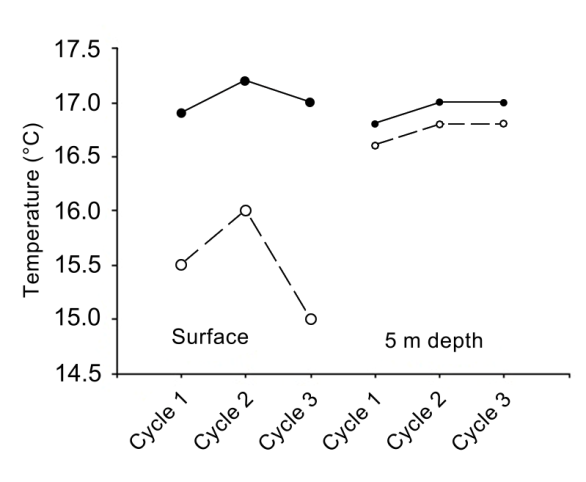
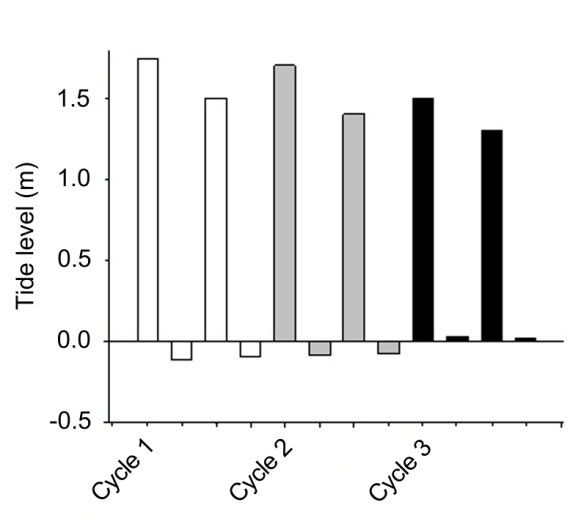
The upwelling index showed maximum values in early February (115.25 m3 s-1 and 88.75 m3 s-1, respectively), and it decreased slightly during the third week before increasing again toward the end of the month (Figure 2). For the purposes of this study, only the maximum and minimum values of the variables are presented because they can be considered as reliable indicators of the feeding conditions faced by juvenile pelagic first feeders and even adult specimens. The temperature in the three circadian cycles varied between 15.0 and 17.2 °C (Figure 3). The maximal temperatures occurred during the hours of maximum solar irradiation, while the minimal values were recorded during the first hours of the day (03:00 to 05:00 hours). The mean comparison analysis revealed that in all three cycles there were no significant differences (with 95% confidence) between the surface temperature and that at 5 m depth. A semi-diurnal tide was registered, with two high tides and two low tides (Figure 4). A slight decrease in tidal amplitude records was observed; the maximum was registered during the first cycle (1.86 m), while the minimum was observed during the third cycle (1.28 m) (Figure 4).
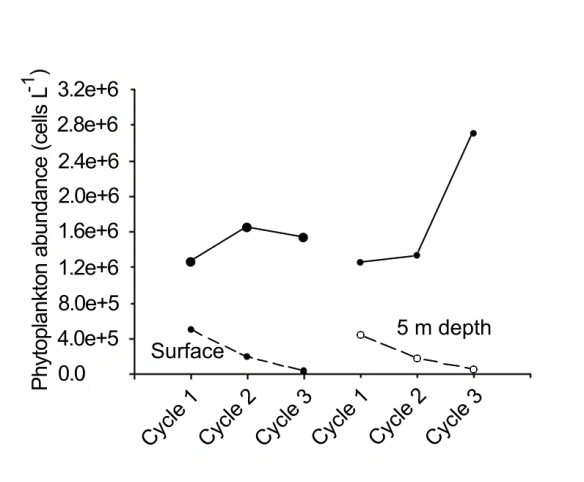
In particular, during the first cycle, the total phytoplankton abundance was relatively high and did not show significant differences between the two depth levels (Figure 5). The maximum recorded was at 16:00, both at the surface (1,263,400 cells L-1) and at 5 m depth (1,253,400 cells L-1). The lowest abundances at both depth levels were recorded at 00:00, ranging from 444,200 cells L-1 to 497,200 cells L-1. The results of the analysis of variance performed for the surface layer showed no significant differences (with 95% confidence) between the abundances found at high tide and low tide hours. In the second cycle, there were also no significant differences (with 95% confidence) in phytoplankton abundance between the levels. The total abundance at the surface showed a maximum of 1,650,200 cells L-1, while at 5 m depth it was 1,332,600 cells L-1; the lowest abundance recorded at the surface was 681,000 cells L-1, while for the deeper level it was 179,200 cells L-1. As with the first two cycles, in the third one there was no difference between the abundance values at both levels. The cell abundance at the surface showed a maximum of 1,531,400 cells L-1, and at the deeper level the maximum was 2,704,000 cells L-1. In contrast, the lowest surface phytoplankton abundances were 31,000 cells L-1 and 57,200 cells L-1 at 5 m depth.
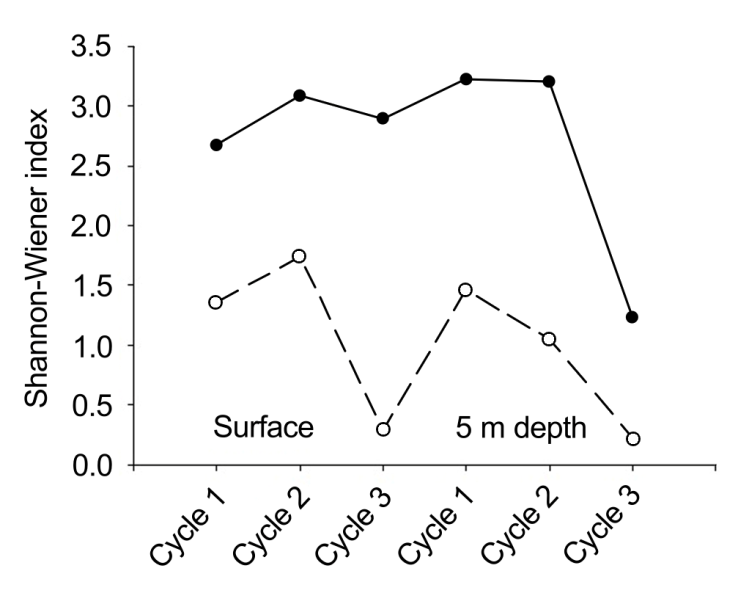
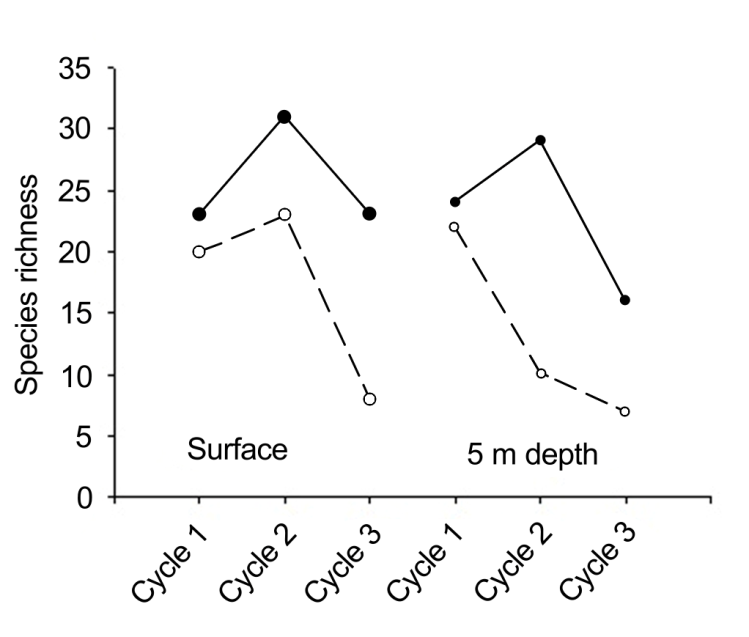
During the first cycle, the microphytoplankton consisted of 86 species and infraspecific taxa (62 diatoms, 18 dinoflagellates, 4 silicoflagellates, an euglenophyte and a cyanobacterium). The dominant species were the chain- forming centric diatoms Chaetoceros compressus Lauder (686,000-745,000 cells L-1), Eucampia zodiacus Ehrenb. (226,400-312,000 cells L-1), Chaetoceros sp. (191,600- 236,400 cells L-1), and the dinoflagellate Prorocentrum koreanum M.S.Han, S.Y.Cho et P.Wang (164,800-209,200 cells L-1). For the second cycle, there was a clear increase in the number of diatom species (70); however, the total number of species was the same as in the first cycle; 11 dinoflagellates, 4 silicoflagellates and an euglenophyte were observed. Diatoms were numerically most important; Chaetoceros compressus (548,600-583,600 cells L-1), C. curvisetus Cleve (243,200-274,400 cells L-1), Coscinodiscus radiatus Ehrenb. (403,200-423,400 cells L-1), and E. zodiacus (106,400- 128,800 cells L-1). During the third cycle, there was a clear decrease in the total number of species (55): 47 diatoms, 4 dinoflagellates, 2 silicoflagellates, an euglenophyte and a cyanobacterium. Eucampia zodiacus (1,893,600-2,288,000 cells L-1) was the most abundant, behaving similarly at both depth levels. Other less abundant species Detonula pumila (Castrac.) Gran (32,600-42,800 cells L-1) and Coscinodiscus radiatus (15,600-92,000 cells L-1). During the first cycle, diversity (H’) and species richness (S) values at the surface were lower compared to those recorded at 5 m depth (Figures 6 & 7); the minimum value was recorded at 16:00 for both surface (H’=1.36, S=20) and the 5 m depth (H’=1.46, S=22). The maximum diversity value for the surface was found at 20:00, (H’=2.67, S=23), while at 5 m it occurred at 10:52 (H’=3.22, S=24). An inverse relationship was observed between phytoplankton abundance and diversity and a direct relationship between diversity and tide maximum levels.
The low diversity observed could be influenced by the dominance of species, such as Chaetoceros sp., E. zodiacus, C. compressus and P. koreanum, which were present at both depth levels. For the second cycle, the diversity index showed higher values. The maximum (H’=3.08, S=31) was at 02:30 at the surface, while for the deeper level it was at 15:30 (H’=3.20, S=29). The minimum diversity value was detected at 05:30 for the surface (H’=1.74, S=23) and at 21:30 for the deeper level (H’=1.05, S=10). In relation to the third cycle, in which important proliferations of E. zodiacus (32,400- 274,400 cells L-1) were found, the diversity was the lowest. Minimum values were reported at 23:30 at both depth levels, for the surface (H’=0.29, S=8) and for the 5 m depth (H’=0.22, S=7); maximum surface diversity values corresponded to 14:30 (H’=2.89, S=23) and at 17:30 for the deeper level (H’=1.23, S=16). We observed that maximum diversity values corresponded to lower phytoplankton abundances. With respect to the tide phase, a pattern opposite to that of the first cycle was observed, i.e., the maximum diversity values coincided with low tide and vice versa. The decrease in the values of this biological indicator was partly due to the proliferation of E. zodiacus.
In terms of the species composition during the second and third cycles, a total of 99 taxa from 60 genera was recorded (Table 1). Diatoms and dinoflagellates were by far the most diverse and abundant groups in number of taxa determined, with 79 and 14 species and infraspecific taxa, respectively. Other taxonomic groups observed were cyanobacteria (2 species), silicoflagellates (2), ebrideans (1) and euglenophytes (1). Species richness variation during the three cycles is shown in Figure 7. Most of the diatom species were planktonic; however, tychoplanktonic (benthic) diatom species were also well represented (Table 1) in our phytoplankton samples, and 25 benthic and 3 benthic- planktonic (based on our observations) species were found.
The nanoplankton showed lower abundances compared to the microphytoplankton. Variations in nanoplankton abundance were as follows: during cycle 2 from 86,400 cells L-1 (08:30) at 5 m depth to 900,000 cells L-1 (15:30) at the surface and during cycle 3 from 9,200 cells L-1 (21:30) to 590,800 cells L-1 (06:30) at the surface. Its contribution to the total phytoplankton abundance during cycle 2 ranged from 3.06 to 65.58% at the surface and from 15.78 to 65.57% at 5 m depth (mean 32.03%); during cycle 3, similar values were 6.38-71.88% and 6.57-81.58% (mean 26.84%), respectively. The mean values of the nanoplankton abundance for both sampling depths together were 296,825±175,379 cells L-1
during cycle 2 and 199,476±157,406 cells L-1 during cycle 3.
Discussion
Temperature in coastal lagoons is variable, which is directly related to the circulation pattern, which, in turn, is affected by geomorphology, winds, river discharge and tides [2]. This agrees with Acosta-Ruiz, et al. [13], the temperature values showed a strong correlation with the tidal cycle, and the maxima of this variable occurred during the ebb hours and the minima during the tidal flow; this evidences the inflow of cold water from the Pacific Ocean and the outflow of warm water from the lagoon system. This pattern was present for the first and third cycles. A slight decrease in tidal amplitude observed in the present study (Figure 4) agrees with the published literature [10, 13, 18]. In a coastal marine area, temperature may act indirectly by causing a stratification/mixing diel cycle [27].
Differences in total cell abundances between the surface and 5 m (Figure 5) may be due to vertical migration as a competitive strategy for phytoplankton under conditions where and when light and nutrients are spatially coincident [28]. Daily variations in phytoplankton have been observed in several studies, both in abundance and biomass [29]. “Observations at sea are hampered by horizontal and vertical transports, by migrations, by grazing and sinking, and also by microdistribution and by sampling errors.” [20]. The cell abundance can be associated with upwelling enrichment events (first cycle; Figure 2), which, in turn, favors the exponential growth of the centric chain-forming diatom Eucampia zodiacus. Lara-Lara, et al. [30] and Cardona-Canizales, et al. [31] noted that nutrients are utilized by phytoplankton until the upwelling process has weakened, which is an influential factor in the proliferation of photoautotrophic populations once the upwelling phenomenon is relaxed. Another important feature is that the maximum concentrations of zooplankton biomass [21] regularly coincided with high phytoplankton abundances, a relationship that may respond to the vertical migration capacity of zooplankton seeking optimal conditions for their development, possibly causing grazing pressure on the autotrophic component of the plankton communities [32].
It is evident that at the end of the first cycle in early February there was an increase in the rate of upwelling, which decreased before reaching the second cycle (Figure 2). Furthermore, the maximum values of the upwelling index coincided in time with the highest tide level (Figure 4). However, the nutrient-rich waters could enter the bay, favoring the access of new species associated with these waters or even favoring the development of species in the bay and thus increasing the diversity values. The increase in species richness could result from the incorporation of species favored by the enrichment of the euphotic zone due to the possible intrusion of eutrophic waters from the upwelling detected between February 7 and 12.
Studies of phytoplankton cell size are important because the different size classes affect physiological rates and ecological functions, including metabolic rate (growth, photosynthesis, respiration) and light absorption [33]. The dominance of the microphytoplankton fraction observed in our study contrasts with the dominance of the nanophytoplankton fraction reported off Bahía Magdalena [34]. This finding is probably due to more eutrophic waters within the lagoon (the study area) compared to oligotrophic conditions in the open sea, off the coastline, as previously mentioned [12]. It is important to emphasize that the aforecited publication refers to the west coast of the State of Baja California Sur, which, although it may influence the lagoon system, is hydrologically different. The dominance of microphytoplankton suggests a shortening of the trophic chain because this group is consumed by macrozooplankton and is of great importance in the feeding of adult clupeids [35, 36, 37]. Sardine stomachs usually contain both phytoplankton and zooplankton prey [38], and the minimal diel variation in stomach content suggests that these fish filter-feed almost continuously [39]. Arthur [40] pointed out that the food material from intestines of sardine larvae was composed mainly of eggs, nauplii and juvenile stages of copepods, and dinoflagellates. Rasoanarivo, et al. [41] found phytoplankton cells between 5 μm (Chlorella spp.) and 130 μm long (Synedra acus Kütz.) and concluded that there are close relationships between the frequency of ingested phytoplankton cells and that in surrounding waters. The size spectra of phytoplankton cells observed in these larvae may indicate that they can feed on both the nanophytoplankton and the microphytoplankton size fractions. Nevertheless, Morote, et al. [42] noted that phytoplankton was eaten only by early-stage anchovy and sardine larvae.
Among the various dominant microphytoplankton species in our study was Eucampia zodiacus. This species has been reported to have a temperate affinity, with recurrent proliferations during the cold months; however, high abundances have also been reported during the warm months [12]. Similar proliferations have been observed in different areas, which is evidence of its high acclimatization to different environments that gives it a cosmopolitan distribution [43, 44]. It is important to note that proliferations of this species can cause ecosystem damage as it is a species that can have a nutritional competitive effect mainly for nitrogen and cause the decline of other primary producers.
It was mentioned that at the end of the first cycle in our study there was an increase in the rate of upwelling (Figure 2). Upwelling events have been reported to have a beneficial effect on phytoplankton days after the event [45] or immediately, depending on the physiology of phytoplankton species [46]. Furuya, et al. [47] point out that under conditions of upwelling or enrichment of the water column with nutrients, which were observed in this study, large-celled diatoms grow twice as fast as small forms, which favors a greater abundance of microphytoplankton. These high concentrations of phytoplankton could represent availability of the vegetative portion of the food, not only for the larvae, but also for the adults of the sardine, thus ensuring the success of the fishery of these organisms in Bahía Magdalena.
Contribution of benthic (tychoplanktonic) diatoms to the phytoplankton species composition observed in our study (Table 1) agrees with the results of Gárate-Lizárraga, et al. [11, 12] who found that in shallow-water systems the diversity rises because of mixing with benthic diatom associations. Changes in the species composition occur during each circadian cycle; they may be due to vertical migration or horizontal transport that has been reported in these types of planktonic and benthic organisms [48]. In this context, photoadaptation plays a primary role because we are considering circadian cycles, which implies contrasting irradiance conditions [32]. This study shows that microalgal blooms can be either punctual or can be carried and maintained throughout the circadian cycle due to the influence of the tide (Figure 4). These phytoplankton blooms can occur at different times of the year, showing that phytoplankton abundance peaks can be influenced by the nutrient-enriched influence of resuspension by upwelling events [11, 49]. We also found that high standing stocks (phytoplankton) observed in the present study allowed us to infer a high availability of food sources for these organisms; we can thus conclude that food was not a limiting factor for their development. In our study, these high phytoplankton abundances occurred just before the high concentrations of sardine larvae reported by Aceves-Medina, et al. [21], ensuring food and success of the different stages of sardine larvae. According to Hjort, et al. [50] food availability is considered one of the main factors affecting the growth rate of pelagic fish larvae.
Circadian feeding patterns have been previously reported for larvae of the sardine and other fish species [51, 52]. Munuera-Fernández, et al. [53] inferred a relationship between the prey availability in the environment and the amount of prey per larva within a gut, despite the circadian feeding variability, the low number of sampling stations and the reported loss of gut contents.
However, circannual rhythms could permit the synchronization between sardine spawning and phytoplankton blooms. Short timescale variations, mainly in temperature (Figure 3), affect the attributes of the phytoplankton, which are manifested as variability in the species composition of microphytoplankton and/ or differential dominance of size fractions. The most representative species in terms of persistence and abundance belonged to diatoms (Eucampia zodiacus, Chaetoceros compressus, C. curvisetus, Coscinodiscus radiatus and Detonula pumila) and dinoflagellates (Prorocentrum koreanum). The dynamics of microalgal proliferations play an important role in the structure of the phytoplankton assemblages as it is an event that influences the variability of temperature and nutrient concentration. On the other hand, in relation to the availability of food for the sardine in its first feeding stage, it was found that the records of abundance (Figure 5) and diversity of phytoplankton species (Figure 6) allow us to infer that there should be no nutritional limitation for their development.
In Mexico, there are no studies related to the analysis of diet of larvae of the different sardine species that could explain the role that phyto- and zooplankton could have on larval growth and successive recruitment levels. Considering the increase in size of larvae during their development, Munuera-Fernández, et al. [53] followed a larval size- dependent approach for the analysis of prey size in the diet of larval sardine. Our results could help us to better understand the pelagic trophic chain that involves Sardinops sagax caerulea and other clupeiforms and will provide more details for future fishery regulations.
Acknowledgments
We are grateful to Marcia M. Gowing (Seattle, WA, USA) for improving the English style, to Alexander Y. Okolodkov (Mexico City, Mexico) for his help with the illustrations, to Manuel E. Martínez-Cruz for technical help (ICIMAP-UV) and to Kenneth N. Mertens (IFREMER, Station de Biologie Marine, Concarneau, France) for his help with the literature. This study was funded by the Dirección de Estudios de Posgrado e Investigación (DEPI 880612) of the Instituto Politécnico Nacional (IPN), Mexico; project: “Validación de las estimaciones mensuales de la biomasa reproductora de S. sagax y O. libertate, en Bahía Magdalena, B.C.S.”. Additional funding was provided by the Secretaría de Investigación y Posgrado of the IPN (projects SIP-20230492 and SIP- 20240611). IGL and GVD are COFAA (Comisión de Operación y Fomento de Actividades Académicas) fellows.
References
-
Ángel MV (1984) La oceanografia biológica y el informe FORE. In: Conferencias Conmemorativas de Anton Bruun, 1982: las ciencias oceánicas para el año 2000. Comisión Oceanográfica Intergubernamental, Colección técnica. UNESCO, Paris, France, pp: 37-40.
-
Yáñez-Arancibia A (1986) Ecología de la zona costera. AGT Editor, México, DF, México, pp: 189.
-
Contreras-Espinosa F (1989) Las lagunas costeras mexicanas. Centro de Ecodesarrollo, Secretaría de Pesca, México, pp: 121.
-
Gárate-Lizárraga I, Band-Schmidt CJ, Verdugo-Díaz G, Muñetón-Gómez MS, Félix-Pico EF (2007) Dinoflagelados (Dinophyceae) del sistema lagunar Magdalena-Almejas. In: Funes-Rodríguez R, Gómez-Gutiérrez J, (Eds.), Estudios ecológicos en Bahía Magdalena. Centro Interdisciplinario de Ciencias Marinas – Instituto Politécnico Nacional. La Paz, BCS, México, pp: 145-174.
-
Félix-Uraga R (1992) Composición por edades de la sardina monterrey Sardinops caeruleus capturada en Bahía Magdalena, B.C.S., México, de 1981 a 1991. Investigaciones Marinas CICIMAR 7(1): 9-13.
-
Gluyas-Millán MG (1989) Época de reproducción, distribución peso-longitud de tallas y relación de la macarela del litoral de Baja California. Investigaciones Marinas, CICIMAR 4(1): 65-72.
-
Ryther JH (1969) Photosynthesis and fish production in the sea. Science 166(3901): 72-76.
-
McFarlane G, Schweig J, Detering J, Hodes V (2010) Diet analysis of Pacific sardine (Sardinops sagax) off the west coast of Vancouver Island, British Columbia from 1997 to 2008. CalCOFI Reports 51: 169-181.
-
Calbet A (2024) The rhythms of plankton. In: The wonders of marine plankton. Springer Cham, pp: 17-21.
-
Nienhuis H, Guerrero-Caballero R (1985) A quantitative analysis of the annual phytoplankton cycle of the Magdalena Lagoon Complex (Mexico). Journal of Plankton Research 7(4): 427-441.
-
Gárate-Lizárraga I, Siqueiros-Beltrones DA (1998) Time variations in phytoplankton assemblages in a subtropical lagoon system after the 1982/83 El Niño event (1984/86). Pacific Science 52: 79-97.
-
Gárate-Lizárraga I, Verdugo-Díaz G, Alfaro-Siqueiros DA (2001a) Variations in phytoplankton assemblages during 1988-1989 in a subtropical lagoon system on the west coast of México. Oceánides 16(1): 1-16.
-
Acosta-Ruiz M de J, Lara-Lara JR (1978) Resultados físicoquímicos en un estudio de variación diurna en el área central de Bahía Magdalena, B.C.S. Ciencias Marinas 5(1): 37-46.
-
Gárate-Lizárraga I, Hernández-Orozco ML, Band- Schmidt CJ, Serrano-Casillas G (2001b) Red tides along the coasts of the Baja California Sur, Mexico (1984 to 2001). Oceánides 16(2): 127-134.
-
Gárate-Lizárraga I, Verdugo-Díaz G, Okolodkov YB (2016) Florecimientos algales nocivos en la costa occidental de Baja California Sur. In: García-Mendoza E, Quijano- Scheggia SI, (Eds.), Florecimientos algales nocivos en México. CICESE, Ensenada, BCS, México, pp: 44-59.
-
Cervantes-Duarte R, Barón-Campis S, Hernández- Becerril DU, Gaxiola-Castro G, Lara-Lara JR (2018) Temporal variability of phytoplankton biomass and primary production off Magdalena Bay, Baja California Sur, Mexico. Ciencias Marinas 44(2): 91-105.
-
Cervantes-Duarte R, Jiménez-Quiroz MC, Funes- Rodríguez R, Hernández-Trujillo S, González-Armas R, Anaya-Godinez E (2021) Interannual variability in the trophic status and water quality of Bahía Magdalena, Mexico, during the 2015-2018 period: TRIX. Regional Studies in Marine Science 42: 101638.
-
Álvarez-Borrego S, Galindo-Bect LA, Chee-Barragán A (1975) Características hidroquímicas de Bahía Magdalena, B.C.S. Ciencias Marinas 2(2): 94-109.
-
Guerrero GR, Cervantes-Duarte R, Jimenez-Illescas AR (1988) Nutrient variation during a tidal cycle at the mouth of the coastal lagoon in the Northwest of México. Indian Journal of Marine Science 17: 235-237.
-
Sournia A (1974) Circadian periodicities in natural populations of marine phytoplankton. Advances in Marine Biology 12: 325-389.
-
Aceves-Medina G, Saldierna-Martínez R, Hernández- Rivas ME (1992) Variación diurna de la abundancia de larvas de peces en la boca de Bahía Magdalena, Baja California Sur, México. Revista de Investigación Científica 3(1): 61-69.
-
Dussart BH (1965) Les différentes catégories de plancton. Hydrobiologia 26: 72-74.
-
Malone TC (1980) Algal size. In: Morris I (Ed.), The physiological ecology of phytoplankton. Blackwell Scientific Publications, Oxford, England, UK, pp: 433- 453.
-
Hasle G (1978) Using the inverted microscope. In: Sournia A (Ed.), Phytoplankton manual. UNESCO, Paris, France, pp: 191-196.
-
Guiry MD, Guiry GM (2024) AlgaeBase. World-wide electronic publication, National University of Ireland, Galway (taxonomic information republished from AlgaeBase with permission of MD Guiry).
-
Brummit RK, Powell CE (1992) Authors of plant names. A list of authors of scientific names of plants with recommended standard forms of their names, including abbreviations. Royal Botanical Gardens, Kew, London, England, UK, pp: 732.
-
Sournia A (1968) Variations saisonnières et nycthémérales du phytoplankton marin et de la production primaire dans une baie tropicale, à Nosy- Bé (Madagascar). Internationale Revue der gesampten Hydrobiologie 53(1): 1-76.
-
Jephson T, Carlsson P (2009) Species and stratification dependent diel vertical migration behaviour of three dinoflagellate species in a laboratory study. Journal of Plankton Research 31(11): 1353-1362.
-
Valdés L, Álvarez-Osorio MT, Lavin A, Varela M, Carballo R (1991) Ciclo anual de parámetros hidrográficos, nutrientes y plancton en la plataforma continental de La Coruña (NO, España). Boletín del Instituto Español de Oceanografía 7(1): 91-138.
-
Lara-Lara JR, Álvarez-Borrego S, Small LF (1980) Variability and tidal exchange of ecological properties in a coastal lagoon. Estuarine and Coastal Marine Science 11(6): 613-637.
-
Cardona-Canizales C, Millán-Núñez R, Lara-Lara JR, Valdez-Holguín JE (1990) Variabilidad temporal de la clorofila a en una zona de surgencias frente a Baja California. Ciencias Marinas 16(2): 53-65.
-
Parsons TR, Maita Y, Lalli CM (1984) A manual the chemical y biological methods for seawater analysis. Pergamon Press, Oxford, Oxfordshire, England, UK, pp: 173.
-
Finkel ZV, Beardall J, Flynn KJ, Quigg A, Rees TAV, Raven JA (2010) Phytoplankton in a changing world: cell size and elemental stoichiometry. Journal of Plankton Resources 32(1): 119-137.
-
Martínez-López A (1993) Distribución espacial del fitoplancton asociada con frentes en la costa occidental de Baja California Sur. Investigaciones Marinas CICIMAR 8(2): 71-86.
-
Varela M, Larrañaga A, Costas E, Rodríguez B (1988) Contenido estomacal de la sardina (Sardina pilchardus Walbaum) durante la campaña Sarcus 871 en las plataformas Cantábrica y de Galicia en febrero de 1987. Boletín del Instituto Español de Oceanografía 5(1): 17-
-
Huntley M (1989) Larval feeding of northern anchovy, Engraulis mordax, on dinoflagellates. Scientia Marina 53(2-3): 239-245.
-
Romero-Ibarra N, Esquivel-Herrera A (1989) Análisis del contenido gástrico de adultos de Opisthonema libertate, y su comparación con el plancton circundante. Investigaciones Marinas CICIMAR 4(2): 267-279.
-
Davies H (1957) The South African pilchard (Sardinops ocellata). Preliminary report on feeding off the west coast, 1953-56. Investigational Report Division of Fisheries South Africa 30: 141-184.
-
van der Lingen CD (1998) Nitrogen excretion and absorption efficiencies of sardine Sardinops sagax fed phytoplankton and zooplankton diets. Marine Ecology Progress Series 175: 67-76.
-
Arthur DK (1976) Food and feeding of larvae of three fishes occurring in the California Current, Sardinops sagax, Engraulis mordax, and Trachurus symmetricus. Fishery Bulletin 74(3): 517-530.
-
Rasoanarivo R, Folack J, Champalbert G, Becker B (1991) Relations entre les communautées phytoplanctoniques et l’alimentation des larves de Sardina pilchardus Walb. dans le golfe de Fos (Méditerranée occidentale): influence de la lumiére sur l’activité alimentaire des larves. Journal of Experimental Marine Biology and Ecology 151(1): 83-92.
-
Morote E, Olivar MP, Villate F, Uriarte I (2010) A comparison of anchovy (Engraulis encrasicolus) and sardine (Sardina pilchardus) larvae feeding in the Northwest Mediterranean: influence of prey availability and ontogeny. ICES Journal of Marine Science 67(5): 897-908.
-
Nishikawa T, Hori Y, Tanida K, Imai I (2007) Population dynamics of the harmful diatom Eucampia zodiacus Ehrenberg causing bleachings of Porphyra thalli in aquaculture in Harima-Nada, the Seto Inland Sea, Japan. Harmful Algae 6(6): 763-773.
-
Nishikawa T, Hori Y, Harada K, Imai I (2013) Annual regularity of reduction and restoration of cell size in the harmful diatom Eucampia zodiacus, and its application to the occurrence prediction of nori bleaching. Plankton and Benthos Research 8(4): 166-170.
-
Otero J, Álvarez-Salgado XA, Bode A (2020) Phytoplankton diversity effect on ecosystem functioning in a coastal upwelling system. Frontiers in Marine Science 7: Article 592255.
-
Marañón E (2015) Cell size as a key determinant of phytoplankton metabolism and community structure. Annual Review of Marine Science 7(1): 241- 264.
-
Furuya K, Takahashi M, Nemoto T (1986) Summer phytoplankton community structure and growth in a regional upwelling area off Hachijo Island, Japan. Journal of Experimental Marine Biology and Ecology 96: 43-55.
-
Valiela I (1984) Marine ecological processes. Springer- Verlag, New York, NY, USA, pp: 546.
-
Gárate-Lizárraga I (1992) Variación espacio temporal de la abundancia fitoplanctónica y de la estructura de las asociaciones microfitoplanctónicas en el Sistema Lagunar Magdalena-Almejas durante 1984-1986. Tesis de Maestría, CICIMAR-IPN, La Paz, B.C.S., México, pp: 84.
-
Hjort J (1914) Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. Rapports et Procés Verbaux des Réunions, Conseil International pour l’Exploration de la Mer 20: 228.
-
Conway DVP (1980) The food of larval blue whiting, Micromesistius potassou (Risso), in the Rockall area. Journal of Fish Biology 16: 709-723.
-
Conway, DVP, Coombs SH, Fernández de Puelles ML, Tranter PRG (1994) Feeding of larval sardine, Sardina pilchardus (Walbaum), off the north coast of Spain. Boletín del Instituto Español de Oceanografía 10(2): 165-175.
-
Munuera-Fernández I, González-Quiros R (2006) Analysis of feeding of Sardina pilchardus (Walbaum, 1792) larval stages in the central Cantabrian Sea. Scientia Marina 70S1: 131-139.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh