Feeding Strategy of the Snail Kite, (Rosthramus Sociabilis) in the Escobalito Reservoir, Toluviejo (Sucre, Colombia)
The feeding strategy of Rostrhamus sociabilis (Accipitridae: Accipitriformes) is studied in The Escobalito reservoir, Toluviejo municipality, Department of Sucre, Colombia. This kite feeds almost exclusively on aquatic molluscs. The type and daily quantity of the species it uses is hereby analyzed, as is the frequency of the catches throughout the day and the characteristics of that activity. This is the first report of the use of Marissa cornuarietis, in addition to Pomacea sp., by the snail kite. The number of individuals of this species that it consumes is greater than that of Pomacea sp., which could be due to their greater availability and smaller size. Capture frequency is higher in the early hours of the morning and the last hours of the afternoon, with short time intervals between each. These kites ingest more frequently shells of intermediate size of both species.
Introduction
The snail kite (gavilán caracolero, Rosthramus sociabilis, Vieillot, 1817) belongs to the order Accipitriformes, which includes three sub species distributed in North, South America and the Caribbean [1, 2], with three recognized subspecies: R.s. plumbeus, in Florida and the Caribbean, R.s. major, from Mexico and Guatemala and R.s. sociabilis present in much of South America. It is a species that feeds mainly on aquatic mollusks, although, according to several authors [3, 4], when the availability of snails is low they feed on turtles and freshwater crabs.
Feeding Strategy of the Snail Kite, (Rosthramus Sociabilis) in the Escobalito Reservoir, Toluviejo (Sucre, Colombia) The study of species with a narrow nutritional range is important because their existence is compromised in the absence of this resource and knowledge of their ecology could be used to establish conservation measures [5]. The feeding strategy of R. sociabilis seems to vary in relation to climatic factors, habitat structure and food availability [6]. This paper provides information about the feeding of this species in The Escobalito reservoir of Toluviejo (Sucre, Colombia) and the results are compared with those obtained by other researchers in Colombia and other countries.
Int J Zoo Animal Biol
Materials and Methods
The Escobalito is a water reservoir built approximately 40 years ago by the company Argos S. A., taking advantage of the course of The Escobalito stream, to use that resource in their industrial activities. It has a drainage basin of 34.14 ha and reaches a depth towards the center of approximately 3 m. It is located at 9° 28’47.33”N, 75° 27´49.00” W, in the Toluviejo municipality, Department of Sucre. On its banks there is ample lush vegetation and native woody trees, which were also planted from the beginning on. Outstanding among these are the ceiba bonga (Ceiba pentandra), guatacán rosado or roble costeño, (Tabebuia rosea), orejero (Enterolobium cyclocarpum), campano (Samanea saman), caracolí (Anacardium excelsum), as well as some fruit trees like mango (Mangifera indica), almendro (Terminalia catappa), and guayaba (Psidium guajava), among others. There are also species of vertebrates such as the pond slider (Trachemys scripta), Dahl's toad- headed turtle (Mesoclemmys dahli), spectacled caiman (Caiman crocodilus), numerous species of waterfowl and mammals such as the capybara (Hydrochoerus hydrochaeris). In the months in which the investigation was conducted, the reservoir presented a considerable area populated by aquatic vegetation composed of different species, in which mollusks, crustaceans and other invertebrates live.
Two previous visits to the work area were made, with the objective of determining the sites frequented by the kites, as well as its movements for the capture of food and the most used hangers. The places where shells accumulated continuously on the periphery of an elevated site were considered as a hanger, for example trees, fallen branches, stones, among others [7, 8].
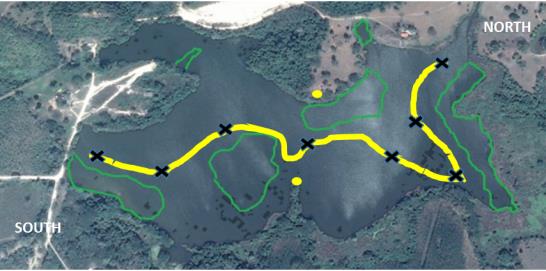
Between January and March 2019, 150 hours of direct observation were conducted using the Lehner focal sampling method, at the time between 06:00 and 18:00 hours. This method is based on the constant observation of a subject for a certain time [9]. For this, two ground stations were established on opposite banks of the lagoon, attended by two researchers and two others made a rowing boat tour covering the path illustrated in Figure 1 with 20-minute stops at sites marked with X. In the same Figure the areas occupied by the large masses of aquatic vegetation are shown.
For the detection and monitoring of the activity of the kites, Gosky Titan astronomical binoculars, 20x80, two Canon EOS Redel XS professional cameras and two lenses, one 70-300 mm and another 1330 mm lens were used (Figure 2). The exact time of each attempt to catch prey was recorded for adult individuals (male and female) and juvenile kites and if it was successful or not. It was considered failed when it was observed with certainty that it had dropped its prey or when, upon returning to the hanger, the bird did not show the feeding behavior because it was not carrying the prey in its claws or beak [8, 10]. The complete prey capture cycle was followed from 06:00 until 18:00 only for three females, the rest of the records were made on different times and days. In addition, the sites that the kites use as a hanger for feeding were noted.

Five trees used as hangers were selected to collect the shells of the mollusks consumed by the kite, accumulated in the soil, under these hangers. These trees were three guayacanes rosados or robles costeños (rosy trumpet tree, Tabebuia rosea), a orejero (monkey-ear tree, Enterolobium cyclocarpum) and a campano (monkey pod tree, Samanea saman). They were all several meters away from the water, so that the shells of the mollusks were not dragged by the current and had a time of permanence in that place. Using this strategy, it was possible to ensure that the shells detected had been consumed by the snail kite, since these did not present any drillings, unlike those consumed by other species, like the Carrao (limpkin, Aramus guarauna [Linnaeus, 1766] [11], common in this lagoon. In addition, these species differ in the feeding site in the reservoir, since the kite does it on the perch and the Carrao on the aquatic vegetation or very close to the shore [7].
The shells found under the above-mentioned trees were sorted by species, according to Vásquez & Sánches, et al. [12] and Linares, et al. [2] (Figure 3) and placed in cloth bags for later measurement using a digital vernier (0.01 mm) and verification of the species.

Results and Discussion
During sampling, 1 male, 9 females and three juveniles were observed making attempts to capture prey in the lagoon (Figure 4A-C). All individuals observed during feeding activity used the strategy of being located in an elevated site (perch) from where they flew and made short and circular flights over the lagoon to launch on their prey [6].

In Figure 5 the sequence of this activity can be appreciated: perching on the hanger, flying over the lagoon, attempt to capture prey by sudden descent with legs pointing down and open claws, taking flight with the snail in its claws, passing snail to the beak and returning to the hanger, where it extracts and consumes the animal, discarding its shell.

Figure 5: Capture and consumption of dams by the Caracolero hawk in the Escobalito lagoon, of Toluviejo, Sucre. A. Hawk perched on the hanger; B. Hawk flies over the lagoon; C. Hawk is launched on the dam; D. Hawk takes flight with the snail in his claw; E. Hawk passes the snail to the beak; F. Hawk returns to the hanger where he extracts the mollusk, discards the shell and consumes it (G and H).
Several authors have argued that the snail kite forages mainly in shallow wetlands, with low-profile emerging vegetation, that is, plants less than one meter above the water surface [13, 14]. The Escobalito reservoir has a greater depth and in this work it was observed that the main hangers used by R. sociabilis for feeding activity were logs, branches, reeds on the shore and other objects that were on the shore of the lagoon or protruded from the surface of the water (81.7%), and tree branches of different species near the shore (18.3%), as Ceiba pentandra (ceiba bonga, kapok tree), Enterolobium cyclocarpum (orejero, monkey-ear tree), Albizia niopoides (guacamayo) Albizia saman (campano, monkey-pod tree) (Figure 6). In all cases, the kite returns to the same hanger for the entire duration of its feeding activity.
the main food of the snail kite [6, 7, 11, 15]. Snyder y Kale, et al. [16] found both species under trees along the Magdalena River and also found more M. cornuarietis (699) than Pomacea sp. (28). These authors explained that the difference in favor of M. cornuarietis was due to the fact that the perch trees were in the water line and the shells had been recently submerged. Since Pomacea has less calcium in its shell, the current carried it easily, while the heavier shels of M. cornuarietis remained on the site. They also explained that perhaps the kites could confuse both species of mollusks from the air, because they were among the aquatic vegetation. In contrast, in the present work the hangers were all far from the water line and the shells collected do not correspond to mollusks ingested by other birds (as they lacked perforations), it can be concluded that in the Escobalito reservoir, at the time the research was conducted, the kites ingest more M. cornuarietis. It is possible that this is due to the greater availability of this mollusc species [7, 8], but this was not corroborated.

Table 1 shows the number of mollusc shells found under the five selected trees. 3,967 shells of two species of aquatic molluscs, Marisa cornuarietis (93%) and Pomacea sp. (7%) (Figure 7A & B), which constitutes a highly significant difference (X2=233.4 p<0.001; 4 gl) (Figure 8). This result is very different from what was stated by several authors who only point to Pomacea as
| Hanger | M. cornuarietis | Pomacea sp. | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| roble | 640 | 148 | 788 | ||||||||
| roble | 584 | 128 | 712 | ||||||||
| roble | 953 | 215 | 802 | ||||||||
| campano | 824 | 194 | 796 | ||||||||
| orejero | 682 | 187 | 869 | ||||||||
| Total | 3683 | 872 | 3967 |
Table 1: Number of shells of aquatic mollusks found under five perch trees in the Escobalito lagoon, of Toluviejo, Sucre.
None of the shells of Pomacea sp. had a hole in the outer surface and only 0.4% of the shells of M. cornuarietis had such a perforation (Table 2). This can be explained in two ways, the snail kite does not make a hole in the shells, since it has a beak adapted to reach the mollusk by cutting the connecting muscle [17, 18, 19], or the few shells found with holes had been preyed on by another species.
| Hanger | M. cornuarietis | Shell with hole | ||||||
|---|---|---|---|---|---|---|---|---|
| roble | 640 | 14 | ||||||
| roble | 584 | 2 | ||||||
| roble | 953 | 0 | ||||||
| campano | 824 | 0 | ||||||
| orejero | 682 | 0 | ||||||
| Total | 3683 | 16 |
Table 2: Number of M. cornuarietis shells collected under different tree species, which had holes in the greater volute.


The depigmentation shown by many of the shells of both species of mollusks is evidence that these hangers have been used for a long time by the snail hawk; many of them even appeared half-buried.
Seven size categories were established for the shells of both mollusc species (Figure 9). Table 3 shows the statistics for the length (mm) of the shells of M. cornuarietis and Pomacea sp. collected.

| Size classes | M. cornuarietis | Pomacea. sp. | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Longer less (mm) | Longitud higher (mm) | ||||||||||||||||
| n | X | S | n | X | |||||||||||||
| 1 | 427 | 12.4 | 0.62 | 69 | 23 | ||||||||||||
| 2 | 432 | 17.2 | 0.5 | 90 | 33.3 | ||||||||||||
| 3 | 735 | 19.7 | 0.44 | 115 | 39.9 | ||||||||||||
| 4 | 740 | 25.8 | 0.39 | 217 | 45.6 | ||||||||||||
| 5 | 484 | 33.1 | 0.42 | 209 | 52.9 | ||||||||||||
| 6 | 445 | 36.2 | 0.45 | 95 | 57.6 | ||||||||||||
| 7 | 420 | 45.6 | 0.51 | 77 | 59.3 |
Table 3: Statisticians of the greater length (mm) of the shells of M. cornuarietis and Pomacea. sp., collected under the trees us
For both species, the shells with highest frequency belong to the intermediate size categories. That is, categories 3 and 4 for M. cornuarietis and 4 and 5 for Pomacea sp., a result similar to that reported by Estela and Naranjo, et al. [7] and Wilcox and Fletcher, et al. [20] for Pomacea sp. Snyder and Kale, et al. [16] reported that 8% of Pomacea sp. and 38% of Marisa cornuarietis found in the piles of discarded shells were alive. They assumed that this was due to the awkward manipulation of large shells, as these were significantly larger than those actually consumed. The latter does not coincide with the present findings, since only one live mollusk of M. cornuarietis was collected, despite there being numerous freshly discarded shells under all the trees.
416 attempts to capture prey were recorded for all the kites observed performing this activity (Figure 10), of which 87% were successful. The greatest number of attempts was observed in the morning hours and at the end of the afternoon, as also pointed out by Beissinger, et al. [3] and Estela and Naranjo, et al. [7] for Pomacea sp. similar information for M. cornuarietis does not appear in the literature.
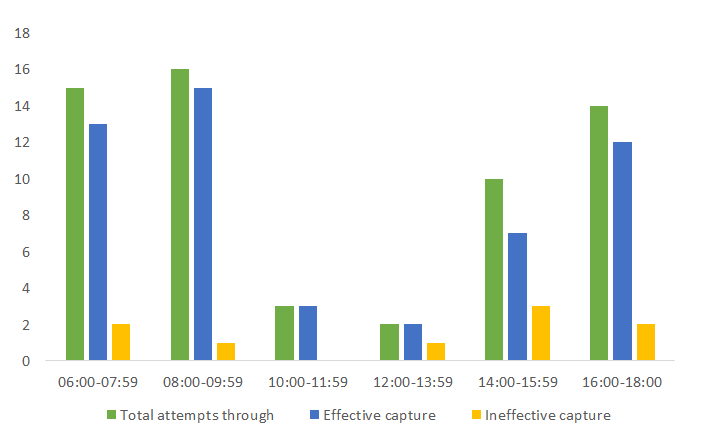
In the case of the three females observed during the diurnal feeding cycle (06:00 - 18:00) (Figure 11) it was also noted that the greatest number of capture attempts, regardless of the species of mollusk captured, occurred in the early hours of the morning (female 1: 27; female 2: 26; female 3: 25), as in Beissinger [3], and the interval between each attempt was shortest in the first hours and increased towards mid-morning. Between 10:00 and 16:00 the number of attempts decreased markedly (female 1: 3; female 2: 4; female 3: 4) and the time intervals between them were very long. In the afternoon, a behavior similar to that of the first hours was observed, since the number of capture attempts increased again (female 1: 14; female 2: 15; female 3: 16) and the time elapsed between each attempt shortened once more until the end of the afternoon.
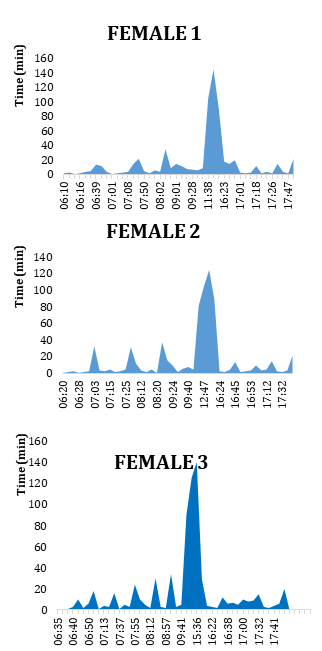
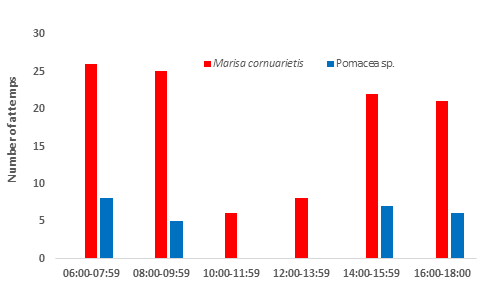
These three females had 44, 45 and 45 attempts respectively with a high capture effectiveness of 91% (40 mollusks consumed), 82% (37 mollusks consumed) and 91% (41 mollusks consumed), which meant an average effectiveness of 88 %. Thus they consume an average of 39 mollusks daily, captured in 44.6 attempts over 12 hours, especially in the early hours and late afternoon. Bergmann, et al. [6] had already reported a high catch effectiveness for R. sociabilis (76%) while Denis and Rodríguez-Ochoa, et al. [16] found that the Cuban snail kite consumes on average 28 snails of Pomacea sp. daily, suggesting that this consumption is due to the high energy needs of the species. The greater number of attempts and mollusks consumed by the three females of R. sociabilis in this work may be due to the need for a greater number of prey, especially because the individuals of the alternative prey species Marisa cornuarietis are smaller (less wet weight) than those of Pomacea sp. The analysis of the attempts to capture both mollusc species by these three females corroborates the latter statement (Figure 12), since the kites make more attempts to capture M. cornuarietis than Pomacea sp., maintaining in both cases the same pattern explained above, that is, more attempts in the morning and at the end of the afternoon.
Conclusion
Although Rosthramus sociabilis is a relatively stenophagous species, it varies its feeding strategies in relation to the availability of food that exists in its habitat. It is able to ingest a larger number of small prey, such as Marisa cornuarietis, to meet its energy needs, if larger ones, Pomacea sp. in this case, are not as abundant. As part of this strategy, it makes a greater number of capture attempts throughout the day, taking short rest periods in the morning and afternoon and much longer ones in the hottest hours.
References
-
Haas S, Kimball R, Martin J, Kitchens M (2009) Genetic divergence among Snail Kite subspecies: implications for the conservation of the endangered Florida Snail Kite Rostrhamus sociabilis. Ibis 151: 181-185.
-
Linares EL, Vera ML (2012) Catálogo de los moluscos continentales de Colombia. Biblioteca José Jerónimo Triana Numero 23. Universidad Nacional de Colombia. Facultad de Ciencias, Instituto de Ciencias Naturales, Bogotá D. C. Colombia 29(3): 360.
-
Beissinger S (1983) Hunting behavior, prey selection, and energetics of snail kite in Guyana: Consumer choice by a specialist. The Auk 100: 84-92.
-
Veiga JO, Raggio JM, Güller RM (2009) Cangrejos y tortugas en la dieta del caracolero (Rostrhamus sociabilis) en Argentina. Nuestras Aves 54: 47-48.
-
Cáceres Guillén M (2013) Comportamiento de forrajeo y uso del hábitat del gavilán caracolero (Rostrhamus sociabilis) en el Parque Nacional Palo Verde, Costa Rica. Tesis presentada como requisito para optar al grado de Magister Scientiae en Conservación y Manejo de Vida Silvestre. Sistema de Estudios de Posgrado de la Universidad Nacional. Heredia. Costa Rica.
-
Bergmann FB, Amaral HLC, Pinto DP, Chivittz CC, Tozetti AM (2013) Foraging activity of the snail kite, Rostrhamus sociabilis (Ave: Accipitridae) in wetlands of southern Brazil. Braz J Biolo 73(2): 245-252.
-
Estela F, Naranjo F (2005) Segregación en el tamaño de caracoles depredados por el Gavilán Caracolero (Rostrhamus sociabilis) y el Carrao (Aramus guarana) en el suroccidente de Colombia. Ornitología Colombiana 3: 36-41.
-
Chinchilla AyI, Barias (2018) Hábitos alimentarios del “Gavilán Caracolero” Rostrhamus sociabilis major (Nelson & Goldman 1933) en dos humedales del complejo Güija, Santa Ana, El Salvador. Trabajo de Grado. Facultad de Ciencias Naturales y Matemática Escuela de Biología, Universidad de El Salvador pp: 148.
-
Lehner P (1998) Handbook of ethological methods. Second edition 541.
-
McMullan MT, Donegan T, Quevedo A (2014) Field Guide to the birds of Colombia. 2nd (Edn.), Fundación Proaves de Colombia, Bogotá.
-
Díaz JM, Blanco JF (1996) Notas comparativas sobre el forrajeo ejercido por el Águila Caracolera (Rostrhamus sociabilis) y la Viudita Alegre (Aramus guarauna) sobre moluscos de agua dulce, en la laguna de Sonso, Valle del Cauca. Cespedesia 21(68): 171- 174.
-
Vázquez A, Sánche J (2015) Clave ilustrada y comentada para la identificación de moluscos gastrópodos fluviales de Cuba. Revista Cubana de Medicina Tropical 67(2): 231-243.
-
Bourne G (1985) The role of probability in Snail Kite foraging. Journal of Animal Ecology 54(3): 697-709.
-
Sykes P (1987) Snail kite nesting ecology in Florida. Florida Field Naturalist 15(3): 57-84.
-
Denis D, Rodríguez Ochoa A (2017) Modelación matemática del consumo de presas y el flujo de energía asociado a la reproducción de Rostrhamus sociabilis (Aves, Falconiformes). Animal Biodiversity and Conservation, 40(2): 221-245.
-
Snyder NFR, Kale H II (1983) Mollusk predation by snail kites in Colombia. The Auk 100(1): 93-97.
-
Méndez P, Curti M, de Montuto K, Benedetti A (2006) Las aves rapaces. Guía didáctica de educación ambiental. Fondo Peregrino. Panamá pp: 1-112.
-
Rueda C, Wiebe A (2014) Snail Kite (Rostrhamus sociabilis), Neotropical Birds. Online (T.S. Schulenber, Eds) Ithaca: Cornell Lab of Ornithology; retrieved from Neotroical Birds.
-
Chapman B, Bolen E (2015) Ecology of North America. 2nd (Edn.). pp: 352.
-
Wilcox RC, Fletcher RJ (2016) Experimental Test of Preferences for an Invasive Prey by an Endangered Predator: Implications for Conservation. PLoS One 11(11): e0165427.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind