Description of Pallisentis (Pallisentis) Paranandai n. sp. (Acanthocephala: Quadrigyridae) from the Intestine of the Great SNakehead Channa marulius (Hamilton) (Channidae) in the Ganga River, India
Pallisentis (Pallisentis) paranandai n. sp. is a freshwater fish parasite in the intestine of Channa marulius (Hamilton) (Actinopterygii: Anabantiformes: Channidae) from the Ganga River and its tributaries, the Indian Subcontinent. Our specimens were somewhat similar to those of Pallisentis nandai Sarkar, 1953 originally described from the liver of the leaffish, Nandus nandus (Hamilton) (Actinopterygii: Perciformes: Nandidae) from the Ganga River delta at Calcutta. Our description of P. paranandai n. sp. includes morphological, chemical, and molecular information distinguishing it from Pallisentis nandai. Both species, P. paranandai n. sp. and P. nandai, have similar anatomical organization but specimens of P. paranandi n. sp. lack proboscis bumps and undulating membrane lining of the proboscis receptacle found in P. nandai, have larger organs, sensory pores and discs on the proboscis, neck, and trunk not found in P. nandai. They also have different hook and spine chemistry (using EDXA) showing much higher levels of Calcium and Phosphorous and considerably lower levels of Sulfur compared to P. nandai. A comparison with the EDXA pattern of another species of Pallisentis, P. indica Mital and Lal, 1976 showed a distinctly different pattern. The sequencing and analysis of 18S region of ribosomal DNA sequences evaluate the relationship between the new species and other species of Pallisentis in the family Quadrigyridae. The phylogenetic tree of maximum likelihood (ML) and Bayesian Inference (BI) were implemented allowing us to validate P. paranandai n. sp. and to suggest that Pallisentis is a monophyletic group.
Introduction
Pallisentis (Pallisentis) Sarkar HL, et al. [1] was described from the liver of the Gangetic leaffish Nandus nandus (Nandidae) in the Ganga delta from a Calcutta fish market [1], and reported since from the livers of the same host throughout the length of the Ganga River and its tributaries from its northern sources in China to its lower delta in eastern India and Bangladesh [2, 3, 4, 5, 6] as well as from the livers of 5 other un-named species of fish [7] and the tank goby, Glossogobius giuris (Gobiidae) [4, 8]. We have recently redescribed P. nandai from N. nandus in Bijnor near the northern reaches of the Ganga correcting inaccuracies and adding new information including SEM, Energy Dispersive X-ray analysis (EDXA), micropores, and DNA analysis thus expanding the body of knowledge about P. nandai in particular and the genus Pallisentis Van Cleave, 1928 in general [8]. We have recently studied new specimens of a similar species of Pallisentis from the intestine of the great snakehead Channa marulius (Hamilton) (Channidae) in the Ganga River, India, that could readily be confused with P. nandai because the general organization of anatomical structures are rather similar. We provide a complete description of the new species and designate differentiating anatomical, chemical (EDXA) and molecular distinctions. In India, the validity of many species of acanthocephalans is often questionable because descriptions were often based on morphological observations lacking detail and illustrations, type specimens were not deposited in recognized museums, and previously published data was frequently ignored by Tadros G, et al. [9, 10, 11, 12, 13]. Molecular data is very scarce for Indian acanthocephalans. We supplement our study with the molecular profile of P. paranandai n. sp. Only about 25 sequences are available for the genus Pallisentis in GenBank database to date. In our present study, we provide the phylogenetic relationship of P. paranandai n. sp. based on the ribosomal 18S region that determines the phylogenetic relationship between the new species and other congeners. Molecular data is very scarce for Indian acanthocephalans as only 03 sequences for 28S gene, 07 for ITS region and 27 sequences for 18S gene are available that belongs to various species. We supplement our study with the molecular profile of P. paranandai n. sp. In our present study, we choose the ribosomal 18S gene to provide the phylogenetic relationship of P. paranandai n. sp. as most of the sequences available on GenBank database belongs to 18S that determines a better relationship between the new species and other congeners.
Material and Methods
Collections
We collected about 50 worms from the intestines of 18 of 32 examined fish between October and November, 2020 in the Ganga River at Bairaj, Bijnor (29º01’N, 77º45’E) in the state of Uttar Pradesh (U.P.), India. The fish were obtained from local fishermen in a small fish market in Bairaj of the 40 extended specimens that were used, 25 were processed for microscopical studies, 4 were used for SEM, and 4 for Gallium hook cuts and EDXA (Energy Dispersive x-ray analysis), 2 were used for molecular studies, and 5 remain in the first author’s collection. Freshly collected specimens were extended in water until proboscides everted then fixed in 70% ethanol for transport to our Arizona, USA laboratory for processing and further studies. The remaining ten specimens were too contorted and were discarded.
Methods for Microscopical Studies
Worms were punctured with a fine needle and subsequently stained in Mayer’s acid carmine, destained in 4% hydrochloric acid in 70% ethanol, dehydrated in ascending concentrations of ethanol (24hr each), and cleared in 100% xylene then in 50% Canada balsam and 50% xylene (24hr each). Whole worms were then mounted in Canada balsam. Measurements are in micrometers, unless otherwise noted; the range is followed by the mean values between parentheses. Width measurements represent maximum width. Trunk length does not include proboscis, neck, or bursa.
Line drawing were created by using a Ken-A-Vision micro-projector (Ward’s Biological Supply Co., Rochester, New York) which uses cool quartz iodine 150W illumination with 10X, 20X, and 43X objective lenses. Images of stained whole mounted specimens were projected vertically on 300 series Bristol draft paper (Starthmore, Westfield, Massachusetts), then traced and inked with India ink. Projected images were identical to the actual specimens being projected. Microscope images were created using 10X and 40X objective lenses of a BH2 light Olympus microscope (Olympus Optical Co., Osachi-shibamiya, Okaya, Nagano, Japan) attached to an AmScope 1000 video camera (United Scope LLC, dba AmScope, Irvine, California), linked to an ASUS laptop equipped with HDMI high-definition multimedia interface system (Taiwan-USA, Fremont, California). Images from the microscope are transferred from the laptop to a USB and stored for subsequent processing on a computer. Forty- two images were made to create used figures.
Specimens were deposited in the University of Nebraska’s State Museum’s Harold W. Manter Laboratory (HWML) collection, Lincoln, Nebraska, USA.
SEM (Scanning Electron Microscopy)
Specimens that had been fixed and stored in 70% ethanol were processed for SEM following standard methods [14]. These included critical point drying (CPD) in sample baskets and mounting on SEM sample mounts (stubs) using conductive double-sided carbon tape. Samples were coated with gold and palladium for 3 minutes using a Polaron #3500 sputter coater ((Quorum (Q150 TES) www. quorumtech.com) establishing an approximate thickness of 20 nm. Samples were placed and observed in an FEI Helios Dual Beam Nanolab 600 (FEI, Hillsboro, Oregon) Scanning Electron Microscope with digital images obtained in the Nanolab software system (FEI, Hillsboro, Oregon). Samples were received under low vacuum conditions using 10 KV, spot size 2, 0.7 Torr using a GSE detector.
Energy Dispersive X-Ray Analysis (EDXA)
Standard methods were used for preparation similar to the SEM procedure. Specimens were examined and positioned with the above SEM instrument, which was equipped with a Phoenix energy-dispersive x-ray analyzer (FEI, Hillsboro, Oregon). X-ray spot analysis and live scan analysis were performed at 16 Kv with a spot size of 5 and results were recorded on charts and stored with digital imaging software attached to a computer. The TEAM (Texture and Elemental Analytical Microscopy) software system (FEI, Hillsboro, Oregon) was used. Data was stored in a USB for future analysis. The data included weight percent and atom percent of the detected elements following correction factors.
Ion Sectioning of Hooks
A dual-beam SEM with a gallium (Ga) ion source (GIS) is used for the LIMS (Liquid Ion Metal Source) part of the process. The hooks of the acanthocephalans were centered on the SEM stage and cross sectioned using a probe current between 0.2nA and 2.1nA according to the rate at which the area is cut. The time of cutting is based on the nature and sensitivity of the tissue. Following the initial cut, the sample also underwent a milling process to obtain a smooth surface. The cut was then analyzed with X-ray at the tip, middle, and base of hooks for chemical ions with an electron beam (Tungsten) to obtain an X-ray spectrum. Results were stored with the attached imaging software. The intensity of the GIS was variable according to the nature of the material being cut.
Molecular Methods
Two specimens fixed in ethanol were individually used for DNA extraction with the use of DNeasy® Blood & Tissue Kit (QIAGEN, Hilden, Germany) following manufacturer’s instructions. The 18S rDNA region was amplified by using the primers, WormA (5′-GCGAATGGCTCATTAAATCAG-3′) + 1270R (5′-CCGTCAATTCCTTTAAGT-3′) [14, 15] and 18SU467F (5′-ATCCAAGGAAGGCAGCAGGC-3′) + 18SL1310R (5′-CTCCACCAACTAAGAACGGC-3′) [16]. For each molecular marker, the following PCR profile was performed, denaturation at 95°C for 3 min, followed by 40 cycles of 94°C for 40 s, annealing at 55°C for 45 s, extension at 72°C for 1min and followed by a final extension at 72°C for 10 min, then stored at 4 ºC. PCR reactions (25μl) consisted of 3.5μl DNA, 1.2μl of each forward and reverse primer, 2.5μl of 10X buffer with MgCl2 (Biotools, Madrid, Spain),1μl of Taq polymerase (1 U, Biotools), 3.2μl of deoxyribonucleoside triphosphates, and 12.4μl of water. PCR products were electrophoresed and purified with the PurelinkTM Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen, Löhne, Germany). PCR products were sequenced for both directions with the same primers mentioned above using Big Dye Terminator vr. 3.1 cycle sequencing kit in ABI 3130 Genetic Analyzer (Applied Biosystems, Boston, MA, USA).
The sequences of 18S obtained in the present study were aligned with sequences of other specimens of Pallisentis species that were available in the GenBank database using the software Thompson JD, et al. [17]. Nucleotide substitution model was selected for 18S gene marker using the jModeltest program [18]. The selected model was GTR + G + I. The 18S datasets was analyzed using maximum likelihood (ML) and Bayesian inference (BI). For ML analyses, we used the program MEGA version 7.0 [19] to obtain tree with 1000 bootstrap replicates in the rapid bootstrap algorithm. Genetic distances (uncorrected p) were estimated with MEGA version 7.0. Phylogenetic tree in BI analyses were generated with Topali 2.5 [20], the Metropolis-coupled Markov chain Monte Carlo (MCMCMC) simulations were run in four independent runs for 10 million generations and sampled every 1000 generations, and burn-in was set to 25%. Sequences of the outgroup Macracanthorhynchus ingens (AF001844) and Moniliformis cryptosaudi (MH401043) of Archiacanthocephala were used for rooting the trees in the 18S gene analyses.
Results
The current distribution of P. paranandai in the the great snakehead C. marulius appears to be in the Ganga River and its tributaries in India but may extend elsewhere within the snakehead’s range of distribution from Pakistan to South
China [21]. The habitats of the host includes lakes, swamps with submerged vegetation, canals, and rivers with rocky or sandy substrate. The fish apparantly acquires infection through its diet of crustaceans, but also feeds on aquatic insects, frogs, snakes, earthworms, tadpoles and fish [22, 23, 24].
We provide a complete description of P. paranandai n. sp. and include morphometric data, SEM and microscope images, molecular analysis, and Energy Dispersive x-ray analysis (EDXA) of hooks and spines of our specimens. We also report the presence of para-receptacle structure (PRS) for the second time in a member of the genus Pallisentis. All previous records of PRS were from eoacanthocephalans with singled-walled proboscis receptacle in the genera Neoechinorhynchus Stiles et Hassall, 1905 and Acanthogyrus (Acanthosentis). Records of PRS in unrelated acanthocephalans with single-walled receptacle are also known. Additional details of proboscis hook roots, trunk spines, and micropores are described and detailed criteria for distinguishing our specimens from those oif P. nandai are included. The following morphological description is based on the microscopical examination of 25 specimens (11 males, 14 females) and 4 others used in the SEM studies. These specimens were collected from the intestines of 18 great snakeheads, C. marulius, between October and November, 2020 in the Ganga River at Bairaj, Bijnor (29º01’N, 77º45’E) in the state of Uttar Pradesh (U.P.)
Morphological Description
Pallisentis paranandai n. sp.
General with characters of family Quadrigyridae, genus Pallisentis, and subgenus Pallisentis as diagnosed by Amin, et al. [25]. Shared structures and spine counts larger and more numerous in females than in males (Table 1). Trunk spinose, curved ventrad posteriorly, small, slender, cylindrical with anterior swelling more prominent in females than in males (Figures 1, 2 & 7). Body wall even dorsoventrally with electron dense micropores with different diameter and distribution in different trunk regions extending to the epicuticle of spines (Figure 19), and up to 0-4 giant hypodermal nuclei dorsally and ventrally (Figures 1,5-7). Sensory pores and plates on proboscis, neck, and anterior trunk (Figures 8, 9, 13, 20 & 21). Transverse lacunar canals connect 2 major lateral longitudinal canals at regular intervals creating appearance of segmentation (Figures 1, 2 & 7). Trunk with triangular spines in 2 zones separated by spine-free zone (Figures 1, 3, 7, 8 & 14). Spines differentially vacuolated and ducted with cortical layer continuous with epicuticle of body wall (Figures 19 & 34). Collar spines (Figure 16) crowded and closely set beginning slightly posterior to anterior end of trunk and extending posteriorly anterior to posterior end of receptacle in both sexes (Figures 1, 3, 8 & 14). Collar spines in two perfect anterior circles with anterior-most circle having fewer spines and posterior irregular rows of spines (Figures 14 & 15). Anterior collar spines strengthened with 1 internal support rod each of equal length to dermal spines (Figure 29) Posterior trunk spines (Figures 17-18) with single, longer and deeply embedded support rod each (Figure 30). Trunk spines in complete circles aligned with transverse lacunar canals at regular intervals (Figure 17) widening posteriorly to level of male reproductive system (from mid-posterior testis to mid- cement gland) (Figures 1&2) and to level near anterior end to female reproductive system (Figure 7). All spines larger in females than males with collar spines about equal antero- posteriorly and trunk spines smaller and more widely spaced posteriorly than anteriorly (Table 1). Proboscis truncated, wider anteriorly and triangulating posteriorly into neck; with 10 rows of 4 hooks each with no protruding bumps between larger hooks (Figures 3, 9 & 10). Large apical organ marked externally (Figure 10) reaching posterior proboscis, with 2 or 3 large giant nuclei (Figures 3 & 27). Hooks most robust but unequal anteriorly in 2 adjacent circles, gradually smaller, and more slender posteriorly (Figures 3, 4, 9-13), with partially solid core and moderate cortical layer (Figures 31-33). All hooks with deeply marked ventrolateral serrations externally (Figures 11, Arrows, 12,13). Roots simple posteriorly directed and slightly curved, shorter than blades (Figure 4). Roots of apical hook most robust; those of other hook slender. Proboscis receptacle single walled, about 3-4 times as long as proboscis, with dissimilar dorsoventral walls, and with prominent triangulate cephalic ganglion slightly anterior to its base. Dorsal receptacle wall pinched close to posterior end at point of attachment of shorter retractor muscle and of insertion of anterior limb of para-receptacle structure (PRS). PRS attaches to body wall just anterior to collar spines and inserts in posterior end of receptacle posteriorly; nucleated basally with accessory cyst (Figure 26). Other retractor muscle longer extending along whole internal ventral wall of receptacle to its posterior end (Figure 3). Two sets of protractor muscles emerging from 2 posterior sites of receptacle, where long and short retractor muscles attach internally to receptacle wall, fan out to attach to body wall posteriorly. Lemnisci unequal, long, extending well posterior to receptacle (Figure 1). Gonopore terminal in males and ventro-terminal in females.
Males (based on 11 mature adults with sperm) see Table 1 for measurements and counts of anatomical structures and spines. Reproductive system in posterior half of trunk Testes contiguous elliptical elongate, with anterior testis slightly longer than posterior testis Sperm ducts drain each testis dorsally, turning ventral as they unite just anterior to cement gland then joining into large thin-walled common seminal vesicle (Figures 1 &2). Cement gland about as long as both testes with many crescent shaped giant nuclei and ducted connection to pear-shaped cement reservoir. Cement reservoir with 2 long and narrow ducts surrounding seminal vesicle to join base of penis posteriorly. Bursa round, muscular, with no sensory structures (Figure 25).
Females (based on 14 adults gravid with ovarian balls and eggs) see Table 1 for measurements and counts of anatomical structures and spines. Gonopore ventro-terminal with thick lipped slit vulva (Figures 22 & 23). Vagina slightly bent, with well-developed sphincter and 2 pairs of paravaginal ligaments extending anteriorly past uterine bell. Uterus somewhat short, thick walled, lobulated with few sphincter-like bulbs. Uterine bell thin walled, funnel-shaped, unattached to body wall, with few gland cells at base (Figures 5&28). Eggs elliptic with smooth surface and moderate polar prolongation of fertilization membrane (Figures 6&24).
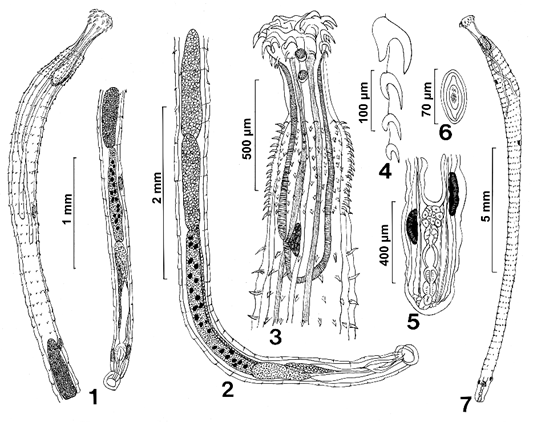
Figures 1-7: Line drawings of specimens of Pallisentis paranandai n. sp. from Channa marulius in the Ganga River, India. (1) A paratype male showing the extent of distribution of trunk spines to the level of the posterior testis, the multiple nucleated cement gland, and the relatively large subcutaneous giant nuclei. (2) A higher magnification of the male reproductive system. (3) The anterior part of a female specimen showing two giant nuclei in the apical organ, tight distribution of collar spines, the long and short retractor muscles, the unequal size of the dorsal and ventral proboscis receptacle wall; and the position and shape of the cephalic ganglion. (4) One row of proboscis hooks and roots. Note that only the anterior hook root is massive. (5) Detail of the female reproductive system. Note the lobulated uterus, the long two pairs of paravaginal filaments, and the slender uterine bell. (6) A ripe egg with some polar prolongation of fertilization membrane. (7) A whole female. Note the extent of distribution of trunk spines to a short distance before the posterior end.
| Males (n=11) | Females (n=14) | |
|---|---|---|
| Trunk L x W (mm) | 5.87-12.50 (8.37) × 0.30-0.57 (0.40)* | 8.75-20.50 (12.70) × 0.32-0.55 (0.45) |
| Proboscis L x W | 166-218 (190) × 208-270 (240) | 187-239 (205) × 270-343 (302) |
| Proboscis hooks | ||
| 1st L x W at base | 68-102 (83) × 20-31 (25) | 87-112 (97) × 28-40 (33) |
| 2nd L x W at base | 67-82 (72) × 15-21 (17) | 72-87 (79) × 15-23 (20) |
| 3rd L x W at base | 50-57 (53) × 10-11 (10) | 53-65 (59) × 10-13 (12) |
| 4th L x W at base | 35-45 (39) × 7-10 (8) | 38-50 (42) × 7-11 (9) |
| Neck L x W | 260-291 (270) × 145-187 (166) | 228-395 (280) × 187-239 (210) |
| Sensory pores & discs | On proboscis, neck, trunk | On proboscis, neck, trunk |
| Giant nuclei | ||
| Apical organ | 2 | 2 or 3 |
| Lemnisci | 2+2, unpronounced | 2 + 2, unpronounced |
| Subcutaneous | 1-4 (1.8) dorsal, 1-4 (2) ventral | 0-4 (1.5) dorsal, 1-2 (1.2) ventral |
| Cement gland | 15-25 (20) | |
| Receptacle L x W | 666-950 (794) × 134-250 (214) | 832-1,070 (946) × 135-291 (244) |
| Short lemniscus L x W | 1,750-2,750 (2,310) × 45-83 (69) | 1,920-2,700 (2,300) × 52-70 (67) |
| Long lemniscus L x W | 2,550-3,330 (2,900) × 57-94 (76) | 2,820-3,850 (3,110) × 52-94 (71) |
| Collar spine rings x no./ ring | 10-15 (13) × 18-20 (19) | 12-17 (15) × 18-22 (19) |
| Collar spine length | 30-40 (33) | 35-47 (42) |
| Trunk spine rings x no./ ring | 24-37 (32) × 12-15 (14) ant. to 4-6 (5) post. | 57-72 (65) × 12-16 (14) ant. to 4-6 (5) post. |
| Trunk spine length | 30-47 (41) ant; 35-47 (39) mid.; 27-40 (33) post. | 40-50 (45) ant; 38-60 (47) mid.; 33-52 (44) post. |
| Anterior aspinose area** | 114-198 (139) | 124-156 (144) |
| Posterior aspinose area** | 2,370-5,000 (3,410) (41% of trunk L) | 790-2,500 (1,580) (12% of trunk L) |
| Anterior testis L x W | 575-1,200 (874) × 146-325 (219) | |
| Posterior testis L x W | 520-1,100 (815) × 146-300 (223) | |
| Cement gland L x W | 884-2,500 (1,542) × 104-300 (190) | |
| Cement Reservoir L x W | 374-675 (514) × 104-250 (183) | |
| Saefftigen’s pouch L x W | 520-1,040 (683) × 104-175 (116) | |
| Common sperm duct L x W | 333-884 (559) × 52-125 (85) | |
| Bursa L x W | 230-260 (250) × 200-291 (250) | |
| ♀ reproductive system L | 614-1,080 (856) (7% of trunk length) | |
| Ripe eggs L x W | 62-82 (73) × 28-37 (32) | |
Table 1: Morphometric characteristic of Pallisentis paranandai from the intestine of Channa marulius in India.
* Range (mean) in micrometers unless otherwise stated. ** Anterior aspinose area: between collar and trunk spines. Posterior aspinose area: posterior to trunk spines. Table 1: Morphometric characteristic of Pallisentis paranandai from the intestine of Channa marulius in India.

Figures 8-13: SEM of specimens of Pallisentis paranandai n. sp. from Channa marulius in the Ganga River, India. (8) Anterior part of a male specimen showing the proboscis, neck, close circles of collar spines starting at anterior trunk, and the more widely spaced trunk spines. Sensory pores (white arrows) and sensory disc (black arrow) are marked. (9) A profile of the proboscis and neck. Note the massive and unequal size of the apical hooks, the sensory pores (white arrows) and sensory disc (black arrow). (10) A near apical view of a proboscis showing the 10 hook rows and the terminal surface of the apical organ (arrow). Note the unequal size of the large apical hooks in 2 adjacent circles. (11-13) Lateral view of apical, middle and posterior hooks, respectively, showing the elevated serrations at their base (arrows in Figure 11) and sensory disc (black arrow) and sensory pore (white arrow, Figure 13).

Figures 14-19: SEM of specimens of Pallisentis paranandai n. sp. from Channa marulius in the Ganga River, India. (14) Anterior male trunk showing the organization of collar and anterior trunk spines. (15) The pattern of collar spines showing 3 anterior circles followed posteriorly by irregular rows of spines. Anterior-most circle with reduced number of spines. (16) A lateral perspective of magnified collar spines. (17) A portion of trunk showing regular circles of trunk spines. (18) A magnified trunk spine. (19) A magnified apical view of a ducted trunk spine (arrow) showing the continuity of micropores on its cortical layer.

Figures 20-25: SEM of specimens of Pallisentis paranandai n. sp. from Channa marulius in the Ganga River, India. (20) A sensory pore in the area of trunk spines. (21) A high magnification of the sensory pore shown in Figure 20. (22) The rounded posterior part of a female trunk showing the position of the gonopore. (23) A high magnification of the slit-like female gonopore shown in Figure 22. (24) A ripe egg, slightly dehydrated. (25) A highly muscular male bursa featuring no specialized sensory structures.
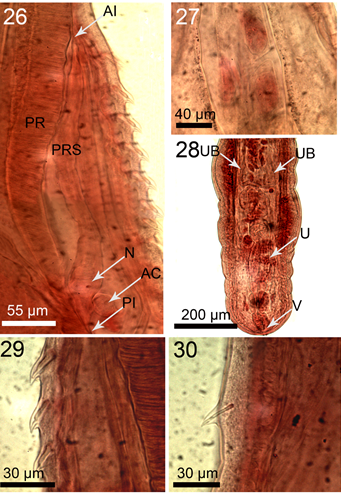
Figures 26-30: Microscope images of specimens of Pallisentis paranandai n. sp. from Channa marulius in the Ganga River, India. (26) The para-receptacle (PRS) structure adjacent to the wall of the proboscis receptacle (PR) in a female specimen showing its anterior insertion (AI) into the body wall, posterior insertion (PI) into the receptacle wall, basal enlarged nucleated are (N), and accessory cyst (AC). (27) Internal anatomy of the apical organ showing 3 giant nuclei. (28) Female reproductive system showing the slightly tilted vagina (V), lobulated uterus (U), and the slender, funnel-shaped, unattached uterine bell (UB). The paravaginal ligaments are not in focus. (29) Lateral view of a collar spine with single rod support. (30) Lateral view of a trunk spine also with single but thinner rod support.
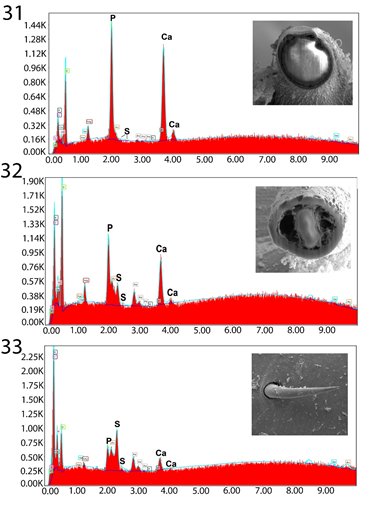
Figures 31-33: Energy Dispersive X-Ray spectrum of Gallium cut anterior mid-hook x-section (31), posterior mid hook x-section (32), and posterior hook edge (33) showing high levels of Calcium and Phosphorous anteriorly and higher level of Sulfur posteriorly. More specific figures in Table II. The x-ray data is the elemental analysis of the hook. Inserts: Gallium cut anterior hook x-section (31), Gallium cut posterior hook x-section (32), and SEM posterior hook surface/edge (33).
Taxonomic Summary
- Type Host: Great snakehead Channa marulius (Hamilton) (Actinopterygii: Anabantiformes: Channidae)
- Type Locality: The Ganga River at Bairaj, Bijnor (29º01’N, 77º45’E), state of Uttar Pradesh (U.P.), India.
- Site of Infection: Intestine. Materials deposited: Harold W. Manter Laboratory (HWML) collection no. 216403 (holotype male), 216404 (allotype female), 216405 (paratypes).
- Etymology: The name of the new species describes its close similarities to Pallisentis nandai Sarkar, 1953.
- Representative DNA Sequence: The 18S rDNA sequence of P. paranandai n. sp. was deposited in GenBank under the accession numbers MW723432 and MW723433. ZooBank LSID#7017AC2A-E80D-4700-B9B5-2D0CE10FEC6A
Remarks
The description of P. paranandai n. sp. is put into perspective in comparison with that of the closely related P. nandai whose original description from Nandus nandus by Sarkar, et al. [1] was the only source of taxonomic information until Amin, et al. [8] provided a comprehensive redescription from the same host species and in the same locality from where our P. paranandai were obtained. Accounts by Soota and Bhattacharya, et al. [2], Bhattacharya, et al. [3], and Naidu, et al. [4] basically repeated Sarkar’s, et al. [1] description. Our description of P. paranandai is compared with our revised description of P. nandai [8] which also included SEM and microscope images, EDXA (Energy Dispersive X-Ray Analysis) and molecular analysis (Table 2).
Despite close morphological similarities between our P. paranadai material (Figures 1-4&7) from the intestine of C. marulius and P. nandai [8] (Figures 1-4) from the liver of N. nandus, distinct differences summarized in (Table 2) readily separate the two species. These distinctions belong in two categories, (1) morphometric differences and (2) discrete non-morphometric characters. (1) Specimens of P. paranandai have markedly larger size of trunk, collar and trunk spines, lemnisci, male and female reproductive systems but smaller size of proboscis and anterior hooks compared to specimens of P. nandai. (2) Specimens of P. paranandai lack the proboscis bumps, the undulating cell layer lining of the receptacle, and the double or triple support rods in collar spines characteristic of P. nandai but have a network of sensory pores and plates on the proboscis, neck, and the trunk (especially in the area of trunk spines) not found in P. nandai. In addition, the hooks and spines of P. paranandai n. sp. contained different levels of Sulfur, Calcium, and Phosphorous than those of P. nandai (Tables 3 & 4). The following results of molecular analysis will further elucidate additional distinctions between the two species.
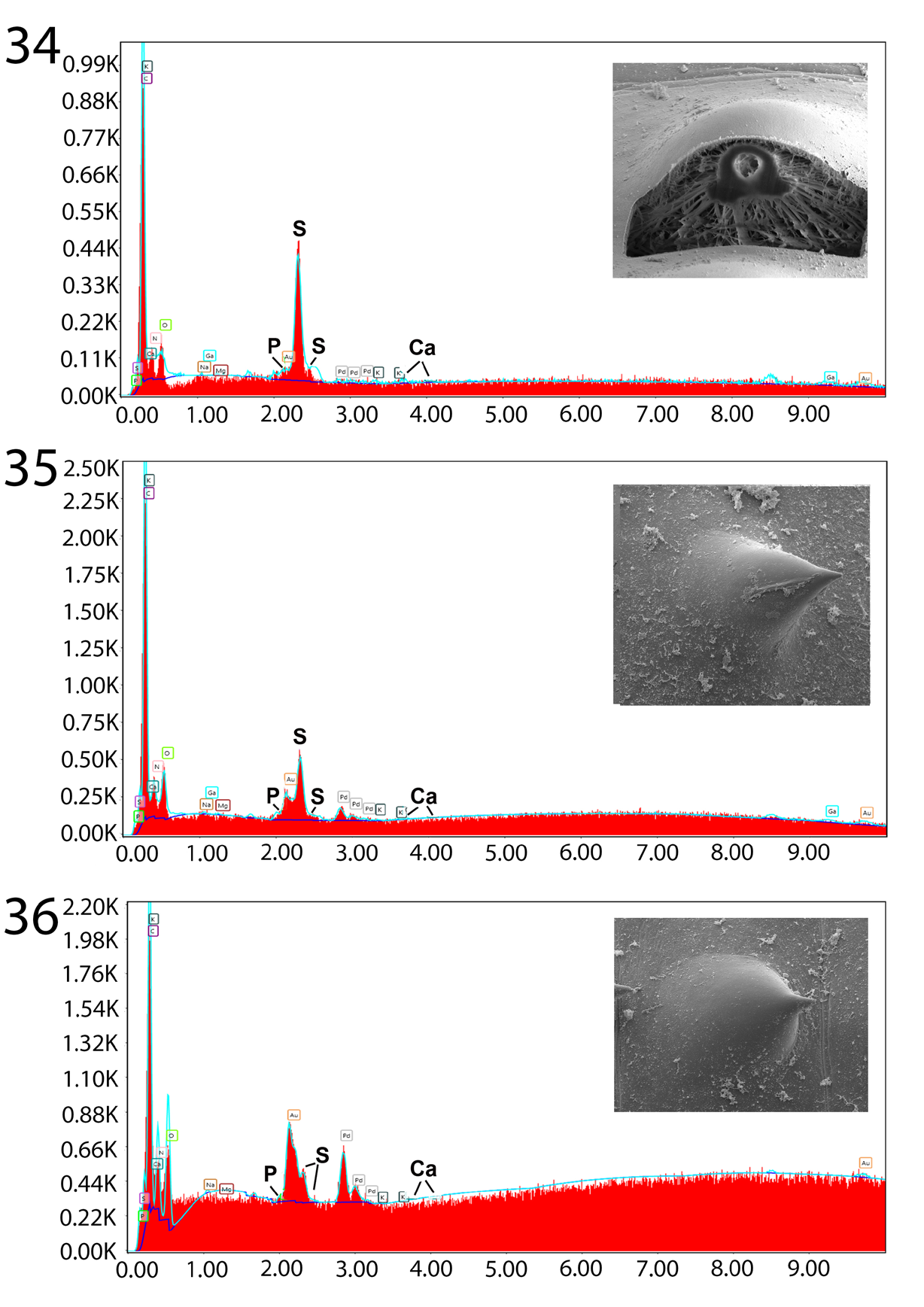
Figures 34-36: Energy Dispersive X-Ray spectrum of Gallium cut anterior collar spine x-section (34), anterior intact trunk spine (35), and posterior intact trunk spine (36) showing moderate levels of sulfur in only collar spines and insignificant levels of Calcium and Phosphorous in all spines. See Table III for more specific figures. Inserts: Gallium cut anterior collar spine cross- section (34), SEM of anterior intact trunk spine (35), and SEM of posterior intact trunk spine (36).
| Species | Pallisentis (P.) nandai | Pallisentis (P.) paranandai |
|---|---|---|
| Host | Nandus nandus | Channa marulius |
| Site of infection | Liver | Intestine |
| Reference | Amin OM, et al. [8] | This paper |
| Sample size | (n=11 males, 12 females) | (n=11 males, 14 females) |
| Trunk L (mm) | ||
| males | 5.25-9.74 (6.90)* | 5.87-12.50 (8.37) |
| females | 7.12-13.12 (10.48) | 8.75-20.50 (12.70) |
| Sensory pores & discs | Not detectable | On proboscis, neck, trunk (Figures 8, 9, 13, 20 & 21) |
| Proboscis L x W | ||
| males | 218-281 (242) × 218-322 (264) | 166-218 (190) × 208-270 (240) |
| females | 198-260 (233) × 198-343 (289) | 187-239 (205) × 270-343 (302) |
| Proboscis bumps** | Present (Figures 8 & 11) | Absent (Figure 9) |
| Anterior hook L | 75-112 (96) | 68-102 (83) |
| Sulfur in mid hook cut | 27.08% | 0% (ant.), 3.25% (post.) (Table 2) |
| Phosphorus in mid hook cut | 2.30% | 21.47% (ant.), 10.35% (post.) (Table 2) |
| Calcium in mid hook cut | 44.91% | 16.29% (post. hook) (Table 2) |
| Sulfur in hook edge cut | 20.82% | 2.42% (ant.), 8.35% (post.) (Table 2) |
| Calcium in hook edge cut | 23.25% | 4.95% (ant.), 5.88% (post.) (Table 2) |
| Collar spine L | ||
| (ant., males) | 25-32 (27) | 30-40 (30) |
| (ant., females) | 27-37 (32) | 35-47 (42) |
| Support rods | 2 or 3 heavy rods per spine (Figure 29) | 1 slender rod per spine |
| Trunk spine L | ||
| (ant., males) | 30-40 (35) | 30-47 (41) |
| (ant., females) | 27-37 (33) | 40-50 (45) |
| Sulfur in cut collar spines | 23.17% (Table 4) | 16.97% (ant.), 8.55% (post.) (Table 3) |
| Sulfur in cut trunk spines | 12.77% (Table 4) | 2.23% (ant.), 1.93% (post.) (Table 3) |
| Giant nuclei | ||
| Subcutaneous | 3 gigantic in one female (Figure 32) | Normal 0-4 dorsal, 0-4 ventral (Figures 1, 4 & 7) |
| Lemniscal | Not detectable | 2 + 2, unpronounced |
| Apical organ | 2 occasional, barely visible | usually 2, prominent (Figure 3) |
| Cement gland | 18-29 [24] (Figure 2) | 15-25 [20] (Figures 1 & 2) |
| Receptacle lining | Undulating cell layer (Figure 3) | Undulating cell layer absent (Figure 3) |
| Short lemniscus L (♂) | 920-2,120 (1,810) | 1,750-2,750 (2,310) |
| Short lemniscus L (♀) | 2,080-2,440 (2,260) | 1,920-2,700 (2,300) |
| Long lemniscus L (♂) | 2,080-2,500 (2,340) | 2,550-3,330 (2,900) |
| Long lemniscus L (♀) | 2,500-2,910 (2,770) | 2,820-3,850 (3,110) |
| Ant. Testis L | 489-842 (622) (Figure 2) | 575-1,200 (874) (Figures 1 & 2) |
| Post. Testis L | 437-832 (606) (Figure 2) | 520-1,100 (815) (Figures 1 & 2) |
| Cement gland L xW | 990-1,510 (1,180) × 130-170 (160) | 884-2,500 (1,542) × 104-300 (190) |
| Female reprod. syst. L | 439-801 (653) | 614-1,080 (856) |
| Uterus | Tube-like (Figures 5 & 35) | Lobulated in sphincter-like segments (Figure 5) |
| Eggs | No eggs | 62-82 (73) × 28-37 (32) (Figure 6) |
Table 2: Distinguishing characteristics between Pallisentis nandai and Pallisentis paranandai in the Ganga River at Bairaj, India
* Range (mean) in micrometers unless if otherwise stated. ** Discrete non-morphometric characters are bolded. Table 2: Distinguishing characteristics between Pallisentis nandai and Pallisentis paranandai in the Ganga River at Bairaj, India.
| Element* | Anterior hook | Posterior hook | ||||
|---|---|---|---|---|---|---|
| Middle** (x-section) | Edge (x- Section) | Intact Hook | Middle (x-section)** | Edge (x- Section) | Intact Hook | |
| Sodium (Na) | 0.06 | 0.1 | 1.43 | 0.14 | 0.01 | 0.59 |
| Magnesium (Mg) | 1.83 | 0.02 | 1.2 | 1.95 | 0.63 | 0.28 |
| Phosphorus (P) | 21.47** | 3.37 | 6.46 | 10.35 | 4.82 | 4.62 |
| Sulphur (S) | 0 | 2.42 | 3.38 | 3.25 | 8.35 | 4.07 |
| Calcium (Ca) | 40.31 | 4.95 | 8.12 | 16.29 | 5.88 | 5.75 |
Table 3: ** Chemical composition of proboscis hooks of male _Pallisentis paranandai_ from the intestine of _Channa marulius_ in I
*Palladium (Pd) and Gold (Au) were used to count the specimens. Gallium was used for the cross cut of the hooks. Other elements (C, O, N) common in organic matter are omitted. Reported in weight (WT)%. See graphs (Figs. 31-33) of these 3 EDXA patterns. *Bolded figures show marked contrast between anterior vs. posterior hooks. Table 3: Chemical composition of proboscis hooks of male Pallisentis paranandai from the intestine of Channa marulius in India.
| Element* | Collar spines | Trunk spines | ||
|---|---|---|---|---|
| Anterior (x-section) | Posterior (x-section) | Anterior (intact) | Posterior (intact) | |
| Sodium (Na) | 0.29 | 0.26 | 0.5 | 0.43 |
| Magnesium (Mg) | 0.08 | 0.08 | 0.01 | 0 |
| Phosphorus (P) | 0.65 | 0.67 | 0.01 | 0.3 |
| Sulphur (S) | 16.94** | 8.55 | 2.23 | 1.93 |
| Calcium (Ca) | 0.95 | 0.13 | 0.06 | 0.12 |
Table 4: ** Chemical composition of collar and trunk spines of male _Pallisentis paranandai_ from the intestine of _Channa maruli
*Palladium (Pd) and Gold (Au) were used to count the specimens. Gallium was used for the cross cut of the spines. Other elements (C, O, N) common in organic matter are omitted. Reported in weight (WT)%. Bolded figures show marked contrast between anterior and posterior collar spines compared to trunk spines. Table 4:** Chemical composition of collar and trunk spines of male Pallisentis paranandai from the intestine of Channa marulius in India.
Our specimens of P. paranandai from C. marulius also bore similarities to Pallisentis (Brevitritospinus) indica Mital, et al. [10], Amin OM, et al. [26] redescribed from the spotted snakehead, Channa punctatus Bloch & Schneider in the Kali Nadi River, Aligarh. The subgenus Brevitritospinus Amin, Heckmann, Ha, Luc, and Doanh [25] was erected to describe species of Pallisentis with “Proboscis hooks in circle 3 about half as long as hooks in circle 2 (and) cement glands usually small with few giant nuclei” compared the subgenus Pallisentis (the subgenus of P. paranandai n. sp.) being characterized with “Proboscis hooks gradually declining in size posteriorly (and) cement glands usually long with many giant nuclei” [25]. Only specimens of Pallisentis indica have a body plan, ducted spines, para-receptacle structure, and eggs similar to P. paranandai but is distinguished from it by having a smaller trunk, Y-shaped collar and trunk spines with multiple rod supports more distant from posterior end of females, no apparent giant hypodermal nuclei, very small and almost equal hooks in circles 3 & 4, almost equal large hooks in circles 1 & 2, all hooks with slender roots, proboscis with protruding bumps in space between hooks in second circle and between those hooks and hooks of third circle, male reproductive structures markedly smaller with only 9-18 giant nuclei in the cement gland, and a tubular uterus [26].
The Para-Receptacle Structure (PRS)
The PRS is here reported in a species of Pallisentis for the third time where its insertion point into the receptacle is associated with the posterior attachment site of the shorter retractor muscles to the receptacle. Like species of Neoechinorhynchus and Acanthosentis, P. paranandai also has a weak single-wall proboscis receptacle. We have examined specimens of 4 other species of Pallisentis in OMA’s personal collection. Only specimens P. indica had a PRS similar to that described in P. nandai. Specimens of the other 3 species examined that did not have PRS are Pallisentis (Pallisentis)
celatus (Van Cleave, 1928) Baylis, 1933, Pallisentis (Brevitritospinus) vietnamensis Amin, Heckmann, Ha, Luc, Donah [25] and Pallisentis (Pallisentis) rexus Wongkham and Whitfield [27].
Energy Dispersive X-ray Analysis (EDXA)
The unique metal composition of hooks (EDXA) demonstrated a considerably high but variable level of Calcium (40.31% and 16.29% in anterior and posterior hooks, respectively) and Phosphorous (21.47% and 10.35%) but neglible level Sulfur (Table 3, Figures 31-33). Anterior and posterior collar spines had high and moderate levels of Sulfur (16.95% and 8.55%) but negligable levels of of Calcium and Phosphorous while all chemicals were very scarce in all trunk spines (Table 4, Figures 34-36). A comparison with the EDXA pattern of other species of Pallisentis, P. indica and P. nandai were considerably different.
Micropores
The electron dense micropores present throughout epidermal surface of the trunk of P. paranandai are described_._ They have been found in various regions of the trunk in different diameters and distributions and were shown to extend into the epicuticle of trunk spines (Figure 19).
Molecular Results
Partial sequences of 18S molecular marker were generated of the new species, Pallisentis paranandai n.
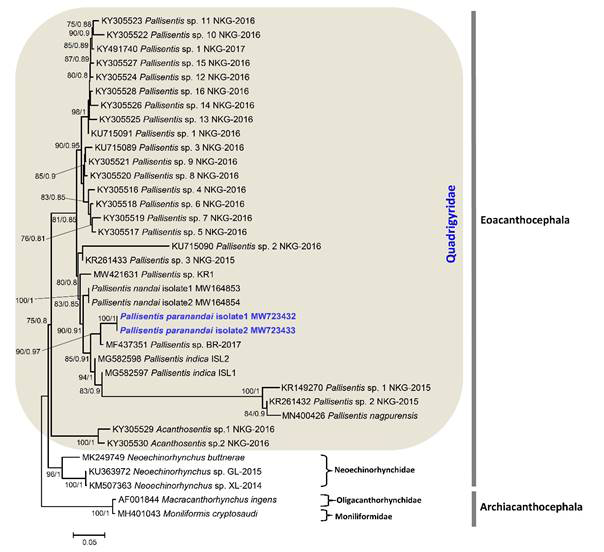
sp. The sequences used for the phylogenetic analysis are listed in (Table 5). The ML and BI trees obtained are similar in topology and showed that the new species is sister to Pallisentis sp. BR-2017 (MF437351) as unpublished data submitted on GenBank by Zimik and Roy (2017) from India belongs to the host Channa striatus with high bootstrap values (Figure 37). The phylogenetic analysis indicated that Pallisentis paranandai n. sp. displays close relationships to other Indian species: P. indica (MG582597 and MG582598) from C. punctata; P. nandai (MW164853 and MW164854) from Nandus nandus [8] (Figure 37). Additionally, the other species clustered in the clade of P. paranandai are: P. anandai (as KR149270 Pallisentis sp. 1 NKG-2015). Gautam NK, et al. [13] from C. punctata, Pallisentis sp. 2 NKG-2015 (KR261432) as unpublished sequence with no host details available on GenBank database and P. nagpurensis (MN400426) that also clustered together with above mentioned species from C. striata as unpublished data by Rana, et al. Although the present results of phylogenetic analysis based on the 18S gene of rDNA for which the maximum sequences i.e., 25 to date is available on GenBank database shows monophyly for the Pallisentis group based on the current studied species. The genetic divergence estimated between P. paranandai n. sp. with Pallisentis sp. BR-2017 (MF437351) is 0.30%, with P. indica ranging from 0.19 to 0.27%, with P. nandai 0.50% and from 1.15 to 1.19% with Pallisentis sp. 2 NKG-2015 (KR261432), Pallisentis sp. 1 NKG-2015 (KR149270) and P. nagpurensis (MN400426), respectively that also shows in the genetic data matrix based on p-distances.
| Species | Host | Location | GenBank accession no. 18S gene | References |
|---|---|---|---|---|
| Class: Eoacanthocephala | ||||
| Family: Quadrigyridae | ||||
| Genus: Pallisentis Van Cleave, 1928 | ||||
| Pallisentis sp. 1 NKG-2015 | HNA | India | KR149270 | Gautam NK, et al. |
| Pallisentis sp. 2 NKG-2015 | HNA | India | KR261432 | Gautam NK, et al. |
| Pallisentis sp. 3 NKG-2015 | HNA | India | KR261433 | Gautam NK, et al. |
| Pallisentis sp. 1 NKG-2016 | Channa striata | India | KU715091 | Gautam NK, et al. |
| Pallisentis sp. 2 NKG-2016 | Channa punctata | India | KU715090 | Gautam NK, et al. |
| Pallisentis sp. 3 NKG-2016 | Channa punctata | India | KU715089 | Gautam NK, et al. |
| Pallisentis sp. 4 NKG-2016 | HNA | India | KY305516 | Gautam NK, et al. |
| Pallisentis sp. 5 NKG-2016 | HNA | India | KY305517 | Gautam NK, et al. |
| Pallisentis sp. 6 NKG-2016 | HNA | India | KY305518 | Gautam NK, et al. |
| Pallisentis sp. 7 NKG-2016 | HNA | India | KY305519 | Gautam NK, et al. |
| Pallisentis sp. 8 NKG-2016 | HNA | India | KY305520 | Gautam NK, et al. |
| Pallisentis sp. 9 NKG-2016 | HNA | India | KY305521 | Gautam NK, et al. |
| Pallisentis sp. 10 NKG-2016 | HNA | India | KY305522 | Gautam NK, et al. |
| Pallisentis sp. 11 NKG-2016 | HNA | India | KY305523 | Gautam NK, et al. |
| Pallisentis sp. 12 NKG-2016 | Channa striata | India | KY305524 | Gautam NK, et al. [28] |
| Pallisentis sp. 13 NKG-2016 | HNA | India | KY305525 | Gautam NK, et al. |
| Pallisentis sp. 14 NKG-2016 | HNA | India | KY305526 | Gautam NK, et al. |
| Pallisentis sp. 15 NKG-2016 | Channa striata | India | KY305527 | Gautam NK, et al. [28] |
| Pallisentis sp. 16 NKG-2016 | HNA | India | KY305528 | Gautam NK, et al. |
| Pallisentis sp. 1 NKG-2017 | HNA | India | KY491740 | Gautam NK, et al. [13] |
| Pallisentis sp. BR-2017 | Channa striata | India | MF437351 | Zimik, et al. |
| Pallisentis nandai Isolate 1 | Nandus nandus | India | MW164853 | Amin OM, et al. [8] |
| Pallisentis nandai Isolate 2 | Nandus nandus | India | MW164854 | Amin OM, et al. [8] |
| Pallisentis sp. KR1 | HNA | India | MW421631 | Rana, et al. * |
| Pallisentis paranandai Isolate 1 | Channa marulius | India | MW723432 | Present study |
| Pallisentis paranandai Isolate 2 | Channa marulius | India | MW723433 | Present study |
| Pallisentis indica ISL1 | Channa punctata | India | MG582597 | Chaudhary A, et al. [29] |
| Pallisentis indica ISL2 | Channa striata | India | MG582598 | Chaudhary A, et al. [29] |
| Pallisentis nagpurensis | Channa punctata | India | MN400426 | Rana, et al. |
| Genus: Acanthosentis Verma and Datta 1929 | ||||
| Acanthosentis sp.1 NKG-2016 | Mystus seenghala | India | KY305529 | Gautam NK, et al. [28] |
| Acanthosentis sp.2 NKG-2016 | Mystus seenghala | India | KY305530 | Gautam NK, et al. [28] |
| Family: Neoechinorhynchidae | ||||
| Genus: Neoechinorhynchus Hamann, 1892 | ||||
| Neoechinorhynchus buttnerae | Gammarus pulex | France | MK249749 | Souza, et al. |
| Neoechinorhynchus sp. XL-2014 | HNA | China | KM507363 | Liu, et al. |
| Neoechinorhynchus sp. GL-2015 | Capoeta aculeate | Iran | KU363972 | Adel, et al. |
| Outgroups | ||||
| Class: Archiacanthocephala | ||||
| Family: Moniliformidae | ||||
| Genus: Moniliformis Travassos, 1915 | ||||
| Moniliformis cryptosaudi | Hemiechinus auritus | Iraq | MH401043 | Amin OM, et al. [30] |
| Family: Oligacanthorhynchidae | ||||
| Genus: Macracanthorhynchus Travassos, 1917 | ||||
| Macracanthorhynchus ingens | Procyon lotor | USA | AF001844 | Near JT, et al. [31] |
Table 5: ** Species of acanthocephalans included in the phylogenetic analysis of 18S gene with information on the host, locality
HNA= host name not available. Species sequenced during the present study is in bold. * Unpublished sequences available on Genbank database. Table 5: Species of acanthocephalans included in the phylogenetic analysis of 18S gene with information on the host, locality and GenBank accession number.
Discussion
The validity of many Indian species of acanthocephalans has often been subject to questions because descriptions were often based on morphological observations lacking detail and illustrations, and type specimens were not deposited in recognized museums [9, 10, 11, 12, 13]. Molecular data is very scarce for Indian acanthocephalans. Our study of P. paranandai includes the classical morphological treatment along with SEM, line drawings, and microscope images of structures not readily available otherwise, as well a study of the chemical analysis of hooks and spines considered to be of taxonomic value in species recognition in very much the same way molecular analysis is useful. We supplement our study of P. paranandai n. sp. with molecular data and compare with other related species, especially P. nandai, and P. indicus. Only about thirty sequences are available for the genus Pallisentis in GenBank database. To date, only 27 sequences are available for 18S rRNA, 07 for ITS1-5.8S-ITS2 cluster, while only 03 sequences are available for 28S rRNA gene which demonstrates the scarcity of molecular data for the species of Pallisentis in comparison to the species diversity available [8]. In our present study, we provide the phylogenetic relationship of P. paranandai n. sp. based on the 18S ribosomal RNA gene (18S rDNA) within the genus Pallisentis (Quadrigyridae) [28, 29, 30, 31].
The Para-Receptacle Structure (PRS)
The PRS inserts anteriorly in the body wall near the neck and posteriorly at the posterior end of the receptacle. The presence of the PRS in Eoacanthocephalans with weak single proboscis receptacle wall was first demonstrated in Neoechinorhynchus (N.) qatarensis Amin, Saoud, Alkuwari by Amin OM, et al. [32]. It had since been reported in other Eoacanthocephalan species of Neoechinorhynchus Stiles and Hassall, 1905 and in Acanthogyrus (Acanthosentis) Verma & Datta, 1929 by Amin OM, et al. [33] and Amin OM, et al. [30, 34, 35, 36, 37]. Occasional records of PRS in unrelated acanthocephalans also with single-walled receptacle include those in Tenuisentis niloticus (Meyer, 1932) as redescribed by Amin OM, et al. [38] from Heterotis niloticus (Cuvier) in Burkina Faso and in Intraproboscis sanghae n. sp. (Gigantorhynchidae) from the black-bellied pangolin Phataginus tetradactyla Lin. in the Central Africal Republic [39]. In the description of the PRS, Amin OM, et al. [32, 40] proposed that it may regulate the hydrostatic pressure in the receptacle to facilitate the retraction and eversion of the proboscis. The PRS is here reported for the first time in P. paranandai n. sp. It has been described in 2 other members of the genus Pallisentis, P. nandai and P. indica.
Micropores
The micropores of P. paranandai, like those reported from other species of the Acanthocephala, are associated with internal crypts and vary in diameter and distribution in different trunk regions corresponding with differential absorption of nutrients. We have reported micropores in a large number of acanthocephalan species [41] and in a few more since, and demonstrated the tunneling from the cuticular surface into the internal crypts by TEM. Amin OM, et al. [42] gave a summary of the structural-functional relationship of the micropores in various acanthocephalan species including Rhadinorhynchus ornatus Van Cleave, 1918, Polymorphus minutus (Goeze, 1782) Lühe, 1911, Moniliformis moniliformis (Bremser, 1811) Travassos (1915), Macracanthorhynchus hirudinaceus (Pallas, 1781) Travassos (1916, 1917), and Sclerocollum rubrimaris Schmidt & Paperna, 1978. Wright and Lumsden and Byram and Fisher reported that the peripheral canals of the micropores are continuous with canalicular crypts [43, 44]. These crypts appear to “constitute a huge increase in external surface area . . . implicated in nutrient up take.” Whitfield estimated a 44-fold increase at a surface density of 15 invaginations per 1µm² of Moniliformis moniliformis (Bremser, 1811) Travassos, 1915 tegumental surface [45]. The micropores and the peripheral canal connections to the canaliculi of the inner layer of the tegument of Corynosoma strumosum (Rudolphi, 1802) Lühe, 1904 from the Caspian seal Pusa caspica (Gmelin) in the Caspian Sea were demonstrated by transmission electron micrographs in Amin OM, et al. [46].
Energy Dispersive X-Ray Analysis (EDXA)
Results of the X-ray scans of the gallium cut hooks (dual beam SEM) of P. paranandai show differential composition and distribution of metals in different hooks with higher levels of Calcium and Phosphorous in anterior compared to posterior hooks, but neglible level Sulfur except at the edge of posterior hooks (Table 3). Anterior collar spines had higher levels of Sulfur than posteriorcollar spines but negligable levels of of Calcium and Phosphorous while all chemicals were very scarce in all trunk spines (Table 4). In P. Nandai, calcium and sulfur were considerably higher at the basal arch of hooks where tension and strength are paramount compared to the hook tip and edge where the level of sulfur was considerably higher (Tables 5, 6 and Figures 39, 40 in Amin OM, et al. [8]). In P. İndica, the EDXA pattern for the hook tip was different that of the hook tip of P. nandai with higher levels of Calcium (20.86% vs. 5.33 %) and Phosphorous (12.04% vs. 0.75%) but the level of Sulfur was lower (4.08% vs. 11.76%) (Table 7).
The chemical elements present in the hooks are typical for acanthocephalans [47, 48, 49, 50]. Note the moderate outer layer (Figure 12 in Amin OM, et al. [8]) of the hook which relates to the Sulfur (S) content (Tables 5 & 6) in the hook of P. nandai which is different than in other acanthocephalans [26, 50, 51, 52]. The high Sulfur content shows up in the outer edge of X-ray scans of hooks (Table 5 & 6, [53]). The hook center in mid cuts has a completely different chemical profile than the cortical layer (Table 6 [8]). X-ray scans (EDXA) provide insight into the hardened components, e.g., calcium and phosphorus, of acanthocephalan hooks. The EDXA appears to be species specific, as in finger prints. EDXA is shown to have significant diagnostic value in acanthocephalan systematics, e.g., Moniliformis cryptosaudi Amin, Heckmann, Sharifdini, Albayati, 2019 was erected based primarily on its EDXA pattern [37].
Molecular Analysis
The present study shows our continued efforts to discover the genetic diversity of this diverse group of species of Pallisentis with more than 30 species [54]. After Amin, et al. [54], additional species were also published by various workers [8, 12, 13, 28, 55, 56]. Regardless the number of species added to the genus Pallisentis, most were described from Channa spp. which suggests their wide distribution in Channidae in India. Thus, the molecular techniques are important for acanthocephalan identification and phylogenetic relationships [35, 36, 37, 57, 58, 59, 60, 61] especially, where the hosts belong to same genus. In India, species of Pallisentis are poorly explored as compared to other genera in regard to molecular biology. ML and BI phylogenetic analysis of 18S gene dataset show that the new species represents an independent clade with high bootstrap values. The new species nests within a clade comprising P. indica (MG582597 and MG582598); P. nandai (MW164853 and MW164854); P. anandai (as KR149270 Pallisentis sp. 1 NKG-2015); Pallisentis sp. 2 NKG-2015 (KR261432); P. nagpurensis (MN400426). We also found that species of Pallisentis show monophyletic assemblage on the basis of present data in the 18S molecular marker as similar to other previous studies [8, 28] although this requires additional investigation with the addition of more molecular data generated from species within this diverse genus.
Conclusion
In conclusion, to the best of our knowledge, the 18S gene is potentially useful molecular marker for investigating Pallisentis phylogeny. The current phylogenetic analyses have also shown the importance of molecular approaches along with morphological data to supplement the study of acanthocephalans and the diversity of species of Pallisentis from freshwater fishes in India.
Acknowledgement
We thank Madison Laurence, Bean Museum (BYU), Provo, Utah for expert help in the preparation and organization of plates and figures and Michael Standing, Electron Optics Laboratory (BYU), for his technical help and expertise in the preparation and production of the SEM images and the EDXA data. We would like to acknowledge the laboratory facilities provided by Department of Zoology, Chaudhary Charan Singh University, Meerut, India. This project was supported by Institutional Grants from the Biology Department, Brigham Young University (BYU), Provo, Utah and from the Parasitology Center, Inc. (PCI), Scottsdale, Arizona. The authors declare no conflicts of interest. The authors declare that they have observed all applicable ethical standards.
References
-
Sarkar HL (1953) On a new Acanthocephala, _Pallisentis_ _nandai_, from the fish _Nandus nandus_ (Hamilton), with notes on other species of the genus. Proceedings of the Zoological Society of Bengal 6(2): 139-147.
-
Soota TD, Bhattacharya SB (1982) On the validity of the species of _Pallisentis_ Van Cleave, 1928 (Acanthocephala: Pallisentidae) from the Indian Subcontinent. Records of the Zoological Survey of India 80: 157-167.
-
Bhattacharya SB (2007) Handbook on Indian Acanthocephala. Zoological Survey of India, Kolkata, pp:
-
Naidu KV (2012) Fauna of India and adjacent countries- Acanthocephala. Zoological Survey of India, Kolkata, pp: 638.
-
Ahmed ATA (1981) Helminth infection in freshwater fishes of Bangladesh. Fish Pathol 15(3-4): 229-236.
-
Parveen S, Sultana S (2014) Infestation of helminth parasites in Gangetic leaffish _Nandus nandus_ (Hamilton, 1822). Bangladesh J Zool 42(2): 183-190.
-
Alam MN, Alam MdJ (2014) A comparative study of endoparasite infestation of _Oreochromis niloticus_ (Linnaeus, 1758) in polluted and non-polluted water bodies of Bangladesh. Int J fauna Biol 1(4): 04-09.
-
Amin OM, Heckmann RA, Chaudhary A, Rubtsova NY, Singh HS (2021a) Redescription and molecular analysis of _Pallisentis (Pallisentis) nandai_ Sarkar, 1953 (Acanthocephala: Quadrigyridae) in India. J Helminthol 95: 1-20.
-
Tadros G (1966) On three new Acanthocephala of the genera _Pallisentis_ Van Cleave, _Saccosentis_ gen. nov. and _Acanthocephalus koelreuther_, from fish. J Helminthol 40: 155-180.
-
Mital RP, Lal SS (1976) Two new acanthocephalan worms _Pallisentis croftoni_ sp. nov. and _P. indica_ sp. nov. (Family-Pallisentidae) from fresh-water fishes of the genus _Ophicephalus._ Indian Journal of Zootomy 17(3): 169-175.
-
Pichelin S, Cribb TH (2001) The status of the Diplosentidae (Acanthocephala: Palaeacanthocephala) and a new family of acanthocephalans from Australian wrasses (Pisces: Labridae). Folia Parasitol 48(4): 289- 303.
-
Gupta R, Maurya R, Saxena AM (2015) Two new species of the genus _Pallisentis_ Van Cleave, 1928 (Acanthocephala: Quadrigyridae) from the intestine of _Channa punctatus_ (Bloch, 1793) from the River Gomti at Lucknow, India. Iran J Parasitol 10(1): 116-121.
-
Gautam NK, Upadhyay MK, Maurya R, Verma SK, Saxena AM (2017) Molecular and morphological study of a new species _Pallisentis anandai_ n. sp. (Quadrigyridae, Van Cleave, 1920) of _Channa punctatus_. Trends in Biosciences 10(7): 1540-1543.
-
Lee RE (1992) Scanning Electron Microscopy and X-Ray Microanalysis. Prentice Hall, Englewood Cliffs, New Jersey, pp: 458.
-
Littlewood DTJ, Olson PD (2001) Small subunit rDNA and the phylum Platyhelminthes: Signal, noise, conflict and compromise.
-
Suzuki N, Hoshino K, Murakami K, Takeyama H, Chow S (2008) Molecular diet analysis of phyllosoma larvae of the Japanese spiny lobster _Palinurus japonicus_ (Decapoda: Crustacea). Mar Biotechnol 10(1): 49-55.
-
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22(22): 4673-4680.
-
Posada D (2008) jModelTest: phylogenetic model averaging. Mol Biol Evol 25(7): 1253-1256.
-
Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol Biol Evol 33(7): 1870-1874.
-
Milne I, Lindner D, Bayer M, Husmeier D, Mcguire G, et al. (2009) TOPALiv2: A rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25(1): 126-127.
-
Talwar PK, Jhingran AG (1992) Inland fishes of India and adjacent countries, Vols. 1-2. Inland fishes of India and adjacent countries 1-2: 1158.
-
Bhuiyan AL (1964) _Fishes of Dacca._ Asiatic Society of Pakistan, Dacca 13: 107-108.
-
Rahman AKA (1989) Freshwater fishes of Bangladesh. Zoological Society of Bangladesh. Department of Zoology, pp: 364.
-
Rainboth WJ (1996) Fishes of the Cambodian Mekong. FAO species identification field guide for fishery purposes, pp: 265.
-
Amin OM, Heckmann RA, Ha NV, Luc PV, Doanh PN (2000) Revision of the genus _Pallisentis_ (Acanthocephala: Quadrigyridae) with the erection of three new subgenera, the description of _Pallisentis_ _(Brevitritospinus) vietnamensis_ subgen. et sp. n., a Key to species of _Pallisentis_, and the description of a new quadrigyrid genus. _Pararaosentis_ gen n. Comparative Parasitology 67(1): 40-50.
-
Amin OM, Heckmann RA, Shareef PAA (2017) Redescription of _Pallisentis_ (_Brevitritospinus_) _indica_ (Acanthocephala: Quadrigyridae) from _Channa_ _punctatus_ Bloch & Schneider (Channidae) in Aligarh, India with New Understandings of Old Structures. J Parasitol 103(3): 251-256.
-
Wongkham W, Whitfield PJ (1999) _Pallisentis rexus_ n. sp. (Eocanthocephala: Quadrigyridae) from snakehead fish, _Channa striata_ Bloch, from Chiang Mai Basin, Thailand. Thai J Agric Sci 32(2): 241-261.
-
Gautam NK, Misra PK, Saxena AM, Monks S (2020) Description of _Pallisentis_ _thapari_ n. sp. and a re- description of _Acanthosentis_ _seenghalae_ (Acanthocephala, Quadrigyridae, Pallisentinae) using morphological and molecular data, with analysis on the validity of the sub- genera of _Pallisentis._ Zootaxa 4766(1): 139-156.
-
Chaudhary A, Amin OM, Singh HS (2019e) Molecular characterization and phylogenetic relationships of _Pallisentis_ (_Brevitritospinus_) _indica_ (Acanthocephala: Quadrigyridae), a parasite of the spotted snakehead (_Channa punctatus_). J Parasitol 105(1): 180-185.
-
Amin OM, Heckmann RA, Sharifdini M, Albayati NY (2019d) _Moniliformis cryptosaudi_ n. sp. (Acanthocephala: Moniliformidae) from the Long-eared Hedgehog _Hemiechinus auritus_ (Gmelin) (Erinaceidae) in Iraq; A Case of Incipient Cryptic Speciation Related to _M. saudi_ in Saudi Arabia. Acta Parasitol 64(1): 195-204.
-
Near JT, Garey JR, Nadler SA (1998) Phylogenetic relationships of the acanthocephala inferred from 18S ribosomal DNA sequences. Mol Phylogenetics Evol 10(3): 287-298.
-
Amin OM, Saoud MFA, Alkuwari KSR (2002) _Neoechinorhynchus qatarensis_ sp. n. (Acanthocephala: Neoechinorhynchidae) from the blue-barred flame parrot fish, _Scarus gobban_ Forsskal, 1775, in Qatar waters of the Arabian Gulf. Parasitol Int 51(2): 171-196.
-
Amin OM (2005) Occurrence of the subgenus _Acanthosentis_ Verma & Datta, 1929 (Acanthocephala; Quadrigyridae) in Japan, with the description of _Acanthogyrus (Acanthosentis) alternatspinus_ sp. n. and _A_. (_A_.) _parareceptaclis_ sp. n. from Lake Biwa drainage fishes and a key to species of the subgenus. Syst Parasitol 60(2): 125-137.
-
Amin OM, Ha NV, Ngo HD (2011a) First report of _Neoechinorhynchus_ (Acanthocephala: Neoechinorhynchidae) from marine fish of the eastern seaboard of Vietnam, with the description of six species. Parasite 18(1): 21-34.
-
Amin OM, Chaudhary A, Heckmann R, Ha NV, Singh HS (2019a) Redescription and molecular analysis of _Neoechinorhynchus_ (_Neoechinorhynchus_) _johnii_ Yamaguti, 1939 (Acanthocephala: Neoechinorhynchidae) from the Pacific Ocean off Vietnam. Parasite 26: 43.
-
Amin OM, Chaudhary A, Heckmann R, Ha NV, Singh HS (2019b) The morphological and molecular description of _Acanthogyrus_ (_Acanthosentis_) _fusiformis_ n. sp. (Acanthocephala: Quadrigyridae) from the catfish _Arius_ sp. (ariidae) in the Pacific Ocean off Vietnam, with notes on zoogeography. Acta Parasitol 64: 779-796.
-
Amin OM, Sharifdini M, Heckmann RA, Ha NV (2019c) On three species of _Neoechinorhynchus_ (Acanthocephala: Neoechinorhynchidae) from the Pacific Ocean off Vietnam with the molecular description of _Neoechinorhynchus_ (_N._) _dimorphospinus_ Amin and Sey, 1996. J Parasitol 105(4): 606-618.
-
Amin OM, Evans RP, Boungou M, Heckmann R (2016) Morphological and molecular description of _Tenuisentis_ _niloticus_ (Meyer, 1932) (Acanthocephala: Tenuisentidae) from _Heterotis_ _niloticus_ (Cuvier) (Actinopterygii: Arapaimidae), in Burkina Faso, with Emendation of the family diagnosis and notes on new features, cryptic genetic diversity and Histopathology. Syst Parasitol 93(2): 173-191.
-
Amin OM, Heckmann RA, Sist B, Basso WU (2021b) A review of the parasite fauna of the black bellied pangolin _Phataginus tetradactyla_ Lin. (Manidae) from Central Africa with the description of _Intraproboscis sanghae_ n. gen., n. sp. (Acanthocephala: Gigantorhynchidae). J Parasitol 107(2): 222-238.
-
Amin OM, Heckmann RA, Standing MD (2007) Structural- functional relationship of the para-receptacle structure in Acanthocephala. Comp Parasitol 74(2): 383-387.
-
Heckmann RA, Amin OM, El-Naggar AM (2013) Micropores of Acanthocephala, a scanning electron microscopy study. Sci Parasitol 14(3): 105-113.
-
Amin OM, Heckmann RA, Radwan NA, Mantuano JS, Alcivar MAZ (2009) Redescription of _Rhadinorhynchus_ _ornatus_ (Acanthocephala: Rhadinorhynchidae) from skipjack tuna, _Katsuwonus pelamis_, collected in the Pacific Ocean off South America, with special reference to new morphological features. J Parasitol 95(3): 656- 664.
-
Wright RD, Lumsden RD (1969) Ultrastructure of the tegumentary pore-canal system of the acanthocephalan _Moniliformis dubius_. J Parasitol 55(5): 993-1003.
-
Byram JE, Fisher FM (1973) The absorptive surface of _Moniliformis dubius_ (Acanthocephala). 1. Fine structure. Tissue Cell 5(4): 553-579.
-
Whitfield PJ (1979) The biology of parasitism: An introduction to the study of associating organisms. University Park Press, Baltimore, Maryland, pp: 27.
-
Amin OM, Heckmann RA, Halajian A, El-Naggar AM (2011b) The morphology of an unique population of _Corynosoma_ _strumosum_ (Acanthocephala, Polymorphidae) from the Caspian seal, _Pusa caspica_, in the land-locked Caspian Sea using SEM, with special notes on histopathology. Acta Parasitol 56(4): 438-445.
-
Heckmann RA, Amin OM, Standing MD (2007) Chemical analysis of metals in Acanthocephalans using energy dispersive X-ray analysis (EDXA, XEDS) in conjunction with a scanning electron microscope (SEM). Comp Parasitol 74(2): 388-391.
-
Heckmann RA, Amin OM, Radwan NAE, Standing MD, Eggett DL, et al. (2012) Fine structure and energy dispersive X-ray analysis (EDXA) of the proboscis hooks of _Radinorynchus ornatus_, Van Cleave 1918 (Rhadinorynchidae: Acanthocephala). Sci Parasitol 13(1): 37-43.
-
Standing MD, Heckmann RA (2014) Features of Acanthocephalan hooks using dual beam preparation and XEDS phase maps. Microscopy and Microanalysis Meeting, Hartford, Connecticut, U.S.A. Poster.
-
Amin OM, Heckmann RA (2017) _Neoandracantha_ _peruensis_ n. gen. n. sp. (Acanthocephala: Polymorphidae) described from cystacanths infecting the ghost crab _Ocypode gaudichaudi_ on the Peruvian coast. Parasite 24: 40.
-
Ha NV, Amin OM, Ngo HD, Heckmann RA (2018) Descriptions of acanthocephalans, _Cathayacanthus_ _spinitruncatus_ (Rhadinorhynchidae) male and _Pararhadinorhynchus magnus_ n. sp. (Diplosentidae), from marine fish of Vietnam, with notes on _Heterosentis_ _holospinus_ (Arhythmacanthidae). Parasite 25: 35.
-
Amin OM, Heckmann RA, Ha NV (2018a) Descriptions of _Acanthocephalus_ _parallelcementglandatus_ (Echinorhynchidae) and _Neoechinorhynchus_ (_N_.) _pennahia_ (Neoechinorhynchidae) (Acanthocephala) from amphibians and fish in Central and Pacific coast of Vietnam, with notes on _N._ (_N_.) _longnucleatus_. Acta Parasitol 63(3): 572-585.
-
Amin OM, Heckmann RA, Ha NV (2018b) Descriptions of _Neorhadinorhynchus nudum_ (Cavisomidae) and _Heterosentis paraholospinus_ n. sp. (Arhythmacanthidae) (Acanthocephala) from fish along the Pacific coast of Vietnam, with notes on biogeography. Journal of Parasitology 104(5): 486-495.
-
Amin OM (2013) Classification of the Acanthocephala. Folia Parasitol 60(4): 273-305.
-
Gautam NK, Maurya R, Saxena AM (2015) Two new species of the genus _Pallisentis_ Van Cleave, 1928 (Acanthocephala: Quadrigyridae) from the intestine of _Channa punctatus_ (Bloch, 1793) from the River Gomti at Lucknow, India. Iranian J Parasitol 10(1): 116-121.
-
Gautam NK, Misra PK, Saxena AM (2019) Four New Species of the Genus _Pallisentis_ (Quadrigyridae, Van Cleave, 1920) from Freshwater Fish in Uttar Pradesh, India. Acta Parasitol 64(1): 71-85.
-
García Varela M, Mendoza Garfias B, Choudhury A, Pérez Ponce de León G (2017) Morphological and molecular data for a new species of _Pomphorhynchus_ Monticelli, 1905 (Acanthocephala: Pomphorhynchidae) in the Mexican redhorse _Moxostoma austrinum_ Bean (Cypriniformes: Catostomidae) in central Mexico. Syst Parasitol 94(9): 989-1006.
-
Li L, Chen HX, Yang Y (2018) Morphological and molecular study of _Neorhadinorhynchus nudus_ (Harada, 1938) (Acanthocephala: Cavisomidae) from _Auxis_ _thazard_ Lacepede (Perciformes: Scombridae) in the South China Sea. Acta Parasitol 63(3): 479-485.
-
García Varela M, Park JK, Hernández Orts JS, Pinacho- Pinacho CD (2019) Morphological and molecular data on a new species of _Plagiorhynchus_ Lühe, 1911 (Acanthocephala: Plagiorhynchidae) from the long- billed curlew (_Numenius americanus_) from northern Mexico. J Helminthol 94: e61.
-
Sharifdini M, Amin OM, Heckmann RA (2020) The Molecular Profile of _Paratrajectura longcementglandatus_ Amin, Heckmann Et Ali, 2018 (Acanthocephala: Transvenidae) from Percid Fishes in the Marine Waters of Iran and Iraq. Helminthologia 57: 1-11.
-
Sharifdini M, Amin OM, Heckmann RA (2021) The molecular profile of _Acanthogyrus_ (_Acanthosentis_) _kashmirensis_ from the Indian subcontinent. Acta Parasitol.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework