The Polymorphism in MC1R Gene and Human the Skin Health
The human melanocortin receptor 1(MC1R) gene has a complex structure; it possesses rich polymorphism and variable spliceosome and participates in several key biological pathways. This paper reviews the structure and function of the human MC1R gene and its polymorphisms on the increased risk of cutaneous carcinoma, nevus, Parkinson's disease, vitiligo, and other diseases to provide a theoretical basis for incorporating the MC1R gene into the risk prediction model of human-related diseases.
Introduction
The melanocortin receptor 1 (MC1R) gene is also named melanocyte-stimulating hormone receptor (MSHR), which encodes MC1R protein, have been described as belonging to the G Protein-coupled receptors (GPCRs) superfamily, a member of the adrenal cortex (MC) receptor family. MC1R protein contains seven transmembrane structures, including one glycosylation site at the extracellular N-terminal and one palmitoylation site at the intracellular C-terminal. It was first discovered in melanocytes and mainly regulated animals and humans’ hair color and skin color [1], and other traits related to pigment deposition [2, 3, 4]. Mammalian MC1R gene is highly conserved with only one exon, but the human MC1R gene has a complex structure with multiple splicing variants and is highly polymorphic. MC1R-001 is the major transcript that encodes 317 amino acids and possesses all the structural characteristics of GPCRs of the A family; it contains exons 2, 3 and 4, and retains two un-spliced insertion sequences between exons 2-3 and 3-4 [5]. In this splicing isoform, the N-terminal segment, cytoplasmic tail, and transmembrane fragment-connecting loops are shorter than most GPCRs [6]. MC1R-002 contains exons 1-4, with the same first 316 amino acids as MC1R-001, but with the extension of 66 amino acids at the C-terminal, encoding a total of 382 amino acids [7]. MC1R-350 was isolated from normal cultured melanocytes and skin sections, with the same first 315 amino acids identical to MC1R-001 and MC1R-002, followed by 35 different amino acid fragments at the C-terminal [8]. MC1R-003 lacks a functional open reading framework [9]. In addition, the human MC1R gene has a highly complex and inefficient Poly (A) site, resulting in intergenic splicing with its immediate downstream neighbor Tubulin-β-Ⅲ (TUBB3) to produce two MC1R isoforms [10, 11]. MC1R gene polymorphisms, particularly red-hair color (RHC) variants associated with red hair, are linked to cutaneous carcinoma, freckle, nevus, vitiligo and Parkinson’s disease. This article reviews the function of the human MC1R gene and its effect on the risk of developing related diseases.
Function of the MC1R Gene
MC1R plays an essential role in pigmentation and regulates eumelanin and pheomelanin synthesis by binding to its natural ligand αmelanocyte-stimulating hormone (α-MSH) or its antagonist agouti signaling protein (ASIP), thereby regulating pigment deposition, which has been widely reported. When MC1R binds to α-MSH, it leads to increased adenylate cyclase (AC) activity and intracellular cyclic adenosine monophosphate(CAMP), then activates protein kinase A(PKA), leading to phosphorylation of cAMP response element-binding protein (CREB), followed by activating the promoter of microphthalmia-associated transcription factor (MITF), promote the expression of MITF, and then activate tyrosinase (TYR), tyrosinase-related protein 1 (TYRP1) and dopachrome tautomerase(DCT) simultaneously promote their expression, resulting in eumelanin synthesis [12]. When MC1R binds to ASIP, this process will be silenced, resulting in functional impairment of MC1R downstream signal and finally a synthesis of pheomelanin. Beta defensin 3(HBD3) is another antagonist of human MC1R, which also blocks the effects of α-MSH on cAMP and tyrosinase activities in human melanocytes [13] (Figure 1).
Receptor protein-tyrosine kinase (C-kit) also plays a role in pigmentation; It binds to stem cell factor (SCF), resulting the recruitment of adapter proteins that contain SRC homology 2(SH2) domain which is associated with the guanine nucleotide exchange factor (GEF) [14, 15]. SH2/GEF complex activates the NRAS/BRAF/MEX/ERK cascade and then activates MITF expression [16]. In melanocytes, MC1R can transactivate C-KIT by SRC, then transmit the signal to extracellular signal-regulated kinase (ERK) via SCF/ c-KIT pathway independent of cAMP [16].
Pheomelanin has a weaker ability to shield ultraviolet radiation (UVR) compared to eumelanin [17] and has been proved to amplify UVA-induced reactive oxygen species (ROS) [10]. Data also suggest that melanin may contribute to DNA oxidative damage and melanoma by producing free radicals in melanocytes, even in the absence of UV radiation [18, 19]. Therefore, the fairer the skin, the greater the damage from UV radiation and the higher the risk of skin diseases.
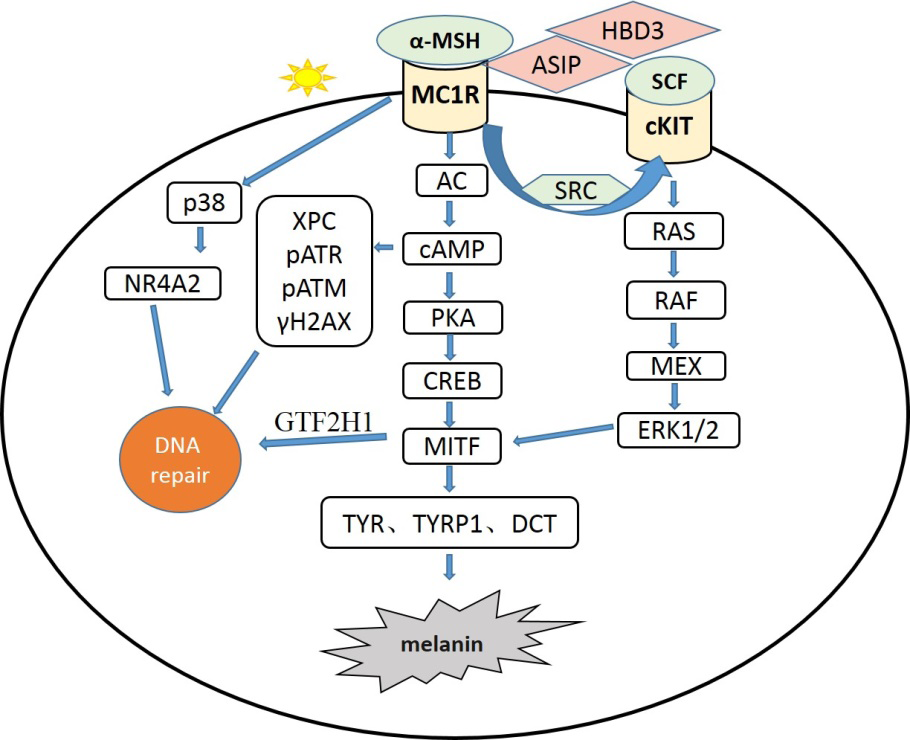
Figure 1: MC1R gene regulates the signal pathway of melanogenesis and DNA repair. Note: The melanocyte membrane receptor MC1R binds to its agonist ɑ-MSH or antagonists ASIP / HBD3 to promte or inhibit melanogenesis by affecting expression of the downstream melanogenic enzymes(TYR,TYRP1,DCT),which are also regulated by SCF/ c-KIT pathway transactivated by MC1R. Meanwhile, the process of DNA repair can be activated by the p38 and/or Ras pathways (independent of cAMP) and those signals dependent on cAMP.
MC1R gene and DNA Repair
UVR consists of about 95% UVA (320-400 nm) and 5% UVB (290-320 nm) [20]. UVA penetrates the skin and reaches the dermis to induce pigmentation through oxidation or distribution of existing melanin; UVB acts on the epidermis to induce skin pigmentation by increasing melanin synthesis [21]. UVR is one of the main environmental factors that damage DNA. The most significant injury caused by UVB is the change at the epidermal level. It leads to covalent bonding between two adjacent pyrimidines, resulting in the formation of cyclobutane pyrimidine dimer (CPD) and 6-4 pyrimidine photoproduct (6-4PP) [22]. Both of these photoproducts distort the structure of DNA by introducing bends or kinks, leading to transcription and replication errors which can eventually lead to permanent mutations if not effectively repaired [23]. By inducing oxidative stress, UVA produces 7, 8-dihydro-8-oxyguanine (8-OXODG) in the dermis that presents in clusters on stretched chromatin fibers, and disruption of chromatin structure may lead to single-strand breaks and double-strand breaks. UVR-induced light leakage and DNA breakage lead to increased genomic instability and the formation of mutations, which are the driving force of melanoma [20].
After UVR, MC1R expression is up-regulated in melanocytes through the synergistic action of p38 protein kinase and upstream transcription factor 1(USF1) [24]. In turn, wild-type MC1R can enhance the activation of P38 in melanocytes after UVR [25]. Activation of P38 promotes phosphorylation of members of the nuclear receptor 4A (NR4A) subfamily and transfers them to the DNA damage site. At the same time, the level of α-MSH also increased after UVR, and α-MSH activated MC1R to trigger the cAMP cascade (Figure 1).On the one hand, the cascade promotes the expression of MITF, and MITF directly controls general transcription and UVR-induced nucleotide excision repair by transactivation of general transcription factor IIH polypeptide 1(GTF2H1) as a core element of DNA transcription factor ⅡH(TFIIH) [26].On the other hand, the increased level of XPC activates DNA damage receptors ATR and ATM, promotes the formation of histone H2AX (γH2AX), and induces the expression of NR4A subfamily members [27, 28]. The increase of γH2AX can recruit DNA repair proteins, including XPC, DDB2 and NR4A, etc., to DNA damage sites for DNA repair [20]. Furthermore, MC1R is involved in many other key biological pathways [12, 29, 30], including cell cycle control of apoptosis and brown adipose tissue metabolism regulation by pro-inflammatory cytokines.
Cutaneum Carcinoma
Cutaneous carcinoma is common cancer that refers to malignant tumors of the skin originating in epithelial tissue, and about 5 million new cases are occurring annually, mainly including melanoma and non-melanoma skin cancers (NMSCs) such as basal cell carcinoma (BCC) and squamous cell carcinoma (SCC). BCC is usually confined, while SCC and melanoma can be potentially metastatic and challenging to treat. The incidence rate of cutaneous carcinoma has increased over the past ten years.
Genome-wide association analysis and meta-analysis showed that RHC variants are associated with an increased risk of melanoma and NMSCs. Compared with the wild-type allele subjects of MC1R, two subjects carrying RHC variants showed a 3- to 5-fold increase in overall cancer incidence. In a study of 220 subjects in Queensland, the prevalence of NMSCs was associated with the presence of the MC1R alleles p.R151C, p.R160W and p.D294H. However, P.v60L, P.V92m and P.r163Q had little effect on BCC and SCC risk. These findings were confirmed and extended in a case-control study of Dutch patients by showing the relative risk of NMSCs in p.D84E and p.H260P carriers was the highest while p.R142H had a slightly lower risk. Data from the M-SKIP project highlighted the association of MC1R variants with the risk of NMSCs development in people living in different geographic regions. It had a greater significant effect on dark-skinned people [31, 32]. Fair-skinned carriers of the MC1R variant had almost twice the risk of BCC compared with their non- carriers. Carriers of p. R163Q variants of medium skin color had a 3-fold higher risk than non-carriers [33].
The subjects that carried any MC1R R allele variant had had a 2- to 4-fold increased risk of melanoma compared with those who had wild-type MC1R allele. In particular, among individuals with multiple alleles of variation, the risk of melanoma was twice as high in carriers of two mutations as in carriers of one mutation [34]. According to the meta- analysis of literature before 2007, the development of melanoma and seven mutations, namely p.D84E, p.R142H, p.R151C, p.I155T, p.R160W, p.R163Q, p.D294H had an association with RHC phenotype; among them, p.D84E, p.R151C, p.I155T, p.R160W and p.R163Q reduced the cell surface expression of the receptor and damaged the cAMP coupling; the surface expression of p.R142H and p.D294H cells was normal but functional response was attenuated [35]. Therefore, the relationship between these MC1R variants and melanoma can be explained by the pigmentary pathway. Individuals with red hair and fair skin cannot normally produce melanin themselves through the pigment synthesis pathway, the mutations are cytotoxic if UVR elevates melanin levels, and the result has also been confirmed and extended in later studies. Recent studies have shown that the number of MC1R variants is also positively associated with an increased risk of melanoma in non- RHC individuals [36]. A pooled analysis of 3830 melanoma cases and 2619 controls showed that the presence of any MC1R variant had a direct effect on melanoma regardless of phenotypic characteristics, increasing the risk of carriers by 60% compared to non-carriers [37]. Therefore, MC1R mutations may also be involved in melanoma development through a pathway independent of pigmentation.
Pigment Spots
Pigment spots and naevus are common biomarkers of melanoma risk [38]. Several studies have reported an association between the freckle and melanoma formation, and the denser the freckle, the higher the risk of illness [39, 40, 41]. A study of 423 primary melanoma patients and 678 controls in the U.S. found that people who have black spots before the age of 15 are more likely to develop melanoma [39].
Pigment spots are a common skin pigmentation feature, mainly including ephelides, solar lentigines and chloasma, which are benign. Freckles are small pigmented spots (generally 1-2 mm) that range in color from red to hazel [42, 43]. Fair-skinned people are more prone to developing freckles, which are believed to be genetically determined and usually first appear at the age of 2-3 yr, then increase during adolescence and often partially disappear with age. They are most frequently found on the face, arms, neck and chest, and become more pigmented during summertime. Solar lentigines are larger than freckles, ranging in diameter from millimetres to centimetres and usually dark brown, unaffected by the season [42, 43, 44]. It often appears on the face, the back of the hand and the lateral forearm and other chronic sunshine skin after 50 years old. Chloasma is an acquired, irregular pattern of facial melanoma, which is the size of the fingertip to a coin or the size of the palm, which is light brown or dark brown [43], mostly related to endocrine, usually symmetrical distribution in the face, forehead, cheeks, nose and back on both sides of the lip around the neck skin, it is more likely to occur in people after puberty, pregnancy women, menopause women and people with darker skin, and can be aggravated after the sun.
The MC1R gene has been proved to be a major factor in the formation of freckles. The impaired function or reduced cell surface receptor expression are two important factors for the occurrence of freckles and solar lentigines [45]. In Caucasians, MC1R genes mutation P.R151C, P.R160W, and P.D294H are strongly associated with red hair, skin cancer, freckles, and solar lentigines. Among them, p.R151C and P. R160W variants showed abnormal cis-transport and intracellular retention, leading to abnormal transport of melanosomes, which accumulated in melanocytes of sunburn [46, 47]. In the Japanese population, the p.92Met allele and the p.163Arg allele of MC1R were positively associated with freckles and severe solar lentigines [48, 49], and homozygote subjects had an extremely increased risk of freckles and solar lentigines. In spite of p.R163Q did not affect MC1R function, it was negatively associated with freckles and severe solar lentigines. In the Indonesian population, the MC1R gene p.V92M was significantly higher in patients with chloasma than in those without chloasma. It is a risk factor for the development of melasma, along with sun exposure and family history [50]. A study of 962 Cutaneous carcinoma patients and 385 non-cutaneous carcinoma controls in the Netherlands showed that people with one or two MC1R variants had a 3-fold and 11-fold increased risk of childhood freckles and a 1.5-fold and 2-fold increased risk of severe sunburn, solar lentigines [51]. Compound heterozygous (R/r) and compound homozygous (R/R and r/r) have an 8-fold increased risk of freckles compared to individual variants (R/- and R/-) [52] Therefore, the risk of freckles is also related to the number of MC1R variants.
Nevus
In addition to pigmentation related features, the most important risk factors for melanoma were the high number of nevi and the presence of atypical nevi, which are usually asymmetrically shaped, ill-defined, irregular brown or red spots greater than or equal to 5 mm in diameter [53]. The number of nevi is inversely correlated with age, usually increasing and then decreasing from adolescence to middle age [52]. MC1R gene is one of the genetic factors affecting nevus formation. Carriers of MC1R mutation, especially those of R allele, are prone to spot and spherical nevi [35]. The nevi with spherical cobblestone pattern are derived from immature dermal melanocytes proliferating in the dermis during infancy, and MC1R is crucial for the proliferation regulation and differentiation of melanocytes. Therefore, the reduced function of MC1R may lead to the immaturity of melanocytes, leading to the punctiform and sphericity of nevi [52].
In a study of 1131 melanoma patients and 869 healthy controls, 79 different MC1R variants were identified, including 69 rare r alleles and five common r alleles(p.V60L, p.V92M, p.I155T, p.R163Q and p.T314T); and 5 R alleles(p. D84E, p.R142H, p.R151C, p.R160W and p.D294H), among them, the r allele was significantly correlated with the number of nevi [54]. Another study explored whether a high number of nevi, red hair, and the MC1R allele act synergistically on melanoma risk and found that nevi with a total number of more than 20 (diameter>5 mm) combined with R/R genotype leading high risk of melanoma for patients living in areas with intense solar irradiation, and R/r genotype was significantly associated with a total number of nevi [55]. A recent study found that although there was no significant association between the number of nevi and phenotypic characteristics in subjects from Leiden, the MC1R gene P. v60L was highly positively associated with the number of moles, especially in women. Therefore, p.V60L variation can be used as an independent predictor of the high number of nevi [56]. MC1R variants affect nevus size and UV radiation effect, resulting in a larger nevi size in carriers of MC1R variants than in wild-type individuals [52].
Other Diseases
Vitiligo is a common acquired depigmentation skin disease caused by the loss of melanocyte function or a decrease in the number of cells in the skin. It is characterized by localized white patches on the skin. Approximately 1% of the world’s population has vitiligo [57]. Although vitiligo is neither physically harmful nor contagious, it is often psychologically devastating and extremely harmful [58]. Blood MC1R gene expression levels were down-regulated in patients with vitiligo compared with controls, while MC1R gene expression levels were also down-regulated in vitiligo diseased tissue compared with healthy controls [59]. The p.V60L, p.V92M, p.R151C and p.R163Q alleles of MC1R gene have been found in the vitiligo population. Still, there is no significant difference in gene frequency between the vitiligo group and the control group [60]. The p.R160W allele is an RHC allele located in the second intracellular loop of MC1R protein, and it’s a component of the protein kinase C (PKC) target sequence consisting of four amino acids. Its frequency is significantly higher in fair-skinned healthy Hungarians than in vitiligo patients, suggesting that this allele has a protective effect on vitiligo [61].
Parkinson’s disease (PD) is a neurodegenerative disease characterized by motor and mental abnormalities. It tends to occur in middle-aged and elderly people and is clinically characterized by loss of dopaminergic neurons in the substantia nigra compact, which can cause great harm [61, 62]. It has been reported that the risk of PD in red hair is twice as high as that in the general population compared with black hair. Therefore, the MC1R gene that regulates the red hair phenotype has also been associated with PD [63]. By a whole-genome sequencing of 870 PD patients and 736 controls from Spain, the p.R160W mutation in MC1R gene was found to be slightly associated with PD [64]. Another study of 272 PD cases and 1185 normal subjects from different races (Caucasian, African American, Asian and others) found that homozygous participants with the p.R151C variant of the MC1R gene had a greater risk of PD than wild-type carriers [65]. Interestingly, other studies found that p. V60L carriers had a lower risk of PD than those without MC1R variants. Both MC1R carriers had a lower risk of PD, leading to the inference that the risk of PD was negatively impacted correlated with the number of MC1R variants [62]. However, studies on Chinese and Singaporean populations found no correlation between MC1R variation and PD [66]. In short, the correlation between MC1R gene variation and PD may be different in different populations.
Expectation
Although MC1R has not been used as a targeted therapy for human-related diseases to date, in vitro and animal studies have shown that it induces improvements in NER function and DNA repair by increasing cAMP levels [67]. It has been reported that MC1R agonists and antagonists may be used as a potential therapeutic approach. For instance, an α-MSH analogue has been used in clinical practice to treat photosensitivity in patients with erythropoietic protoporphyria [68]. Based on the information available in the literature to date, further studies are needed to provide data on the role of MC1R genomic risks that have potential clinical implications for the early detection of treatment and prevention strategies.
With the rapid advancement of knowledge, future directions should be to determine the biological mechanisms of non-pigment MC1R function evaluate gene-gene and gene- environment interactions, and incorporate MC1R variants into skin cancer and Parkinson’s disease risk prediction models for diseases such as vitiligo and test their effectiveness as prevention strategies in eliciting risk-reducing behaviors to tailor treatment options for patients.
Funding
This article was supported by the Project of the Chongqing Natural Science Foundation (cstc2019jcyj-msxmX0063), the Project of the Chongqing Fundamental Research and Frontier Exploration (cstc2018jcyjAX-0042).
References
-
Mountjoy KG, Robbins LS, Mortrud MT (1992) The cloning of a family of genes that encode the melanocortin receptors. Science 257(5074): 1248-1251.
-
Yu W, Wang C, Xin Q (2013) Non-synonymous SNPs in MC1R gene are associated with the extended black variant in domestic ducks (Anas platyrhynchos). Anim Genet 44(2): 214-216.
-
Van Raamsdonk CD, Barsh GS, Wakamatsu K (2009) Independent regulation of hair and skin color by two G protein-coupled pathways. Pigment Cell Melanoma Res 22(6): 819-826.
-
García Borrón JC, Abdel Malek Z, Jiménez Cervantes C (2014) MC1R, the cAMP pathway, and the response to solar UV: extending the horizon beyond pigmentation. Pigment Cell Melanoma Res 27(5): 699-720.
-
Katritch V, Cherezov V, Stevens RC (2013) Structure- function of the G protein-coupled receptor superfamily. Annu Rev Pharmacol Toxicol 53: 531-556.
-
Herraiz C, Garcia Borron JC, Jiménez Cervantes C (2017) MC1R signaling. Intracellular partners and pathophysiological implications. Biochim Biophys Acta Mol Basis Dis 1863(10): 2448-2461.
-
Tan CP, Mckee KK, Weinberg DH (1999) Molecular analysis of a new splice variant of the human melanocortin-1 receptor. FEBS Lett 451(2): 137-141.
-
Rouzaud F, Costin GE, Yamaguchi Y (2006) Regulation of constitutive and UVR-induced skin pigmentation by melanocortin 1 receptor isoforms. FASEB J 20(11): 1927-1929.
-
Herraiz C, Garcia Borron JC, Jiménez Cervantes C (2017) MC1R signaling. Intracellular partners and pathophysiological implications. Biochim Biophys Acta Mol Basis Dis 1863(10PtA): 2448-2461.
-
Dalziel M, Kolesnichenko M, Das Neves RP (2011) Alpha- MSH regulates intergenic splicing of MC1R and TUBB3 in human melanocytes. Nucleic Acids Res 39(6): 2378- 2392.
-
Herraiz C, Olivares C, Castejón Griñán M (2015) Functional Characterization of MC1R-TUBB3 Intergenic Splice Variants of the Human Melanocortin 1 Receptor. PLoS One 10(12): e0144757.
-
Manganelli M, Guida S, Ferretta A (2021) Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention. Genes 12(7): 1093.
-
Swope VB, Jameson JA, Mcfarland KL (2012) Defining MC1R regulation in human melanocytes by its agonist α-melanocortin and antagonists agouti signaling protein and β-defensin 3. J Invest Dermatol, 132(9): 2255-2262.
-
Lemmon MA, Pinchasi D, Zhou M (1997) Kit receptor dimerization is driven by bivalent binding of stem cell factor. J Biol Chem 272(10): 6311-6317.
-
Kuang D, Zhao X, Xiao G (2008) Stem cell factor/c-kit signaling mediated cardiac stem cell migration via activation of p38 MAPK. Basic research in cardiology 103(3): 265-273.
-
Herraiz C, Journé F, Abdel Malek Z (2011) Signaling from the human melanocortin 1 receptor to ERK1 and ERK2 mitogen-activated protein kinases involves transactivation of cKIT. Basic Res Cardiol 25(1): 138- 156.
-
Vincensi MR, D’ischia M, Napolitano A (1998) Phaeomelanin versus eumelanin as a chemical indicator of ultraviolet sensitivity in fair-skinned subjects at high risk for melanoma: a pilot study. Melanoma Res 8(1): 53- 58.
-
Mitra D, Luo X, Morgan A (2012) An ultraviolet-radiation- independent pathway to melanoma carcinogenesis in the red hair/fair skin background. Nature 491(7424): 449-453.
-
Prota G (2000) Melanins, melanogenesis and melanocytes: looking at their functional significance from the chemist’s viewpoint. Pigment Cell Res 13(4): 283-293.
-
Yin K, Sturm RA, Smith AG (2014) MC1R and NR4A receptors in cellular stress and DNA repair: implications for UVR protection. Exp Dermatol 23(7): 449-452.
-
Wolber R, Schlenz K, Wakamatsu K (2008) Pigmentation effects of solar-simulated radiation as compared with UVA and UVB radiation. Pigment Cell Melanoma Res 21(4): 487-491.
-
Besaratinia A, Yoon JI, Schroeder C (2011) Wavelength dependence of ultraviolet radiation-induced DNA damage as determined by laser irradiation suggests that cyclobutane pyrimidine dimers are the principal DNA lesions produced by terrestrial sunlight. FASEB J 25(9): 3079-3091.
-
Pleasance ED, Cheetham RK, Stephens PJ (2010) A comprehensive catalogue of somatic mutations from a human cancer genome. Nature 463(7278): 191-196.
-
Corre S, Primot A, Sviderskaya E (2004) UV-induced expression of key component of the tanning process, the POMC and MC1R genes, is dependent on the p-38- activated upstream stimulating factor-1 (USF-1). J Biol Chem 279(49): 51226-51233.
-
Wong SS, Ainger SA, Leonard JH (2012) MC1R variant allele effects on UVR-induced phosphorylation of p38, p53, and DDB2 repair protein responses in melanocytic cells in culture. J Invest Dermatol 132(5): 1452-1461.
-
Seoane M, Buhs S, Iglesias P (2019) Lineage-specific control of TFIIH by MITF determines transcriptional homeostasis and DNA repair. Oncogene 38(19): 3616- 3635.
-
Scully R, Xie A (2013) Double strand break repair functions of histone H2AX. Mutat Res 750(1-2): 5-14.
-
Jarrett SG, Wolf Horrell EM, Christian PA (2014) PKA- mediated phosphorylation of ATR promotes recruitment of XPA to UV-induced DNA damage. Mol Cell 54(6): 999- 1011.
-
Hill RP, Macneil S, Haycock JW (2006) Melanocyte stimulating hormone peptides inhibit TNF-alpha signaling in human dermal fibroblast cells. Peptides 27(2): 421-430.
-
Vivas-García Y, Falletta P, Liebing J (2020) Lineage- Restricted Regulation of SCD and Fatty Acid Saturation by MITF Controls Melanoma Phenotypic Plasticity. Mol Cell 77(1): 120-137.e129.
-
Pasquali E, García Borrón JC, Fargnoli MC (2015) MC1R variants increased the risk of sporadic cutaneous melanoma in darker-pigmented Caucasians: a pooled- analysis from the M-SKIP project. International journal of cancer 136(3): 618-631.
-
Tagliabue E, Fargnoli MC, Gandini S (2015) MC1R gene variants and non-melanoma skin cancer: a pooled- analysis from the M-SKIP project. Br J Cancer 113(2): 354-363.
-
Scherer D, Bermejo JL, Rudnai P (2008) MC1R variants associated susceptibility to basal cell carcinoma of skin: interaction with host factors and XRCC3 polymorphism. Int J Cancer 122(8): 1787-1793.
-
Ozola A, Azarjana K, Doniņa S (2013) Melanoma risk associated with MC1R gene variants in Latvia and the functional analysis of rare variants. Cancer Genet 206(3): 81-91.
-
Raimondi S, Sera F, Gandini S (2008) MC1R variants, melanoma and red hair color phenotype: a meta-analysis. International journal of cancer 122(12): 2753-2760.
-
Williams PF, Olsen CM, Hayward NK (2011) Melanocortin 1 receptor and risk of cutaneous melanoma: a meta- analysis and estimates of population burden. Int J Cancer 129(7): 1730-1740.
-
Tagliabue E, Gandini S, Bellocco R (2018) MC1R variants as melanoma risk factors independent of at-risk phenotypic characteristics: a pooled analysis from the M-SKIP project. Cancer Manag Res 10: 1143-1154.
-
Duffy DL, Box NF, Chen W (2004) Interactive effects of MC1R and OCA2 on melanoma risk phenotypes [J]. Hum Mol Genet 13(4): 447-461.
-
Titus Ernstoff L, Perry AE, Spencer SK (2005) Pigmentary characteristics and moles in relation to melanoma risk. Int J Cancer 116(1): 144-149.
-
Mackie RM (1998) Incidence, risk factors and prevention of melanoma. Eur J Cancer 3: 3-6.
-
Bliss JM, Ford D, Swerdlow AJ (1995) Risk of cutaneous melanoma associated with pigmentation characteristics and freckling: systematic overview of 10 case-control studies. The International Melanoma Analysis Group (IMAGE). Int J Cancer 62(4): 367-376.
-
Ezzedine K, Mauger E, Latreille J (2013) Freckles and solar lentigines have different risk factors in Caucasian women. J Eur Acad Dermatol Venereol 27(3): 345-356.
-
Plensdorf S, Martinez J (2009) Common pigmentation disorders. Am Fam Physician 79(2): 109-116.
-
Cario-Andre M, Lepreux S, Pain C (2004) Perilesional vs. lesional skin changes in senile lentigo. J Cutan Pathol 31(6): 441-447.
-
Papp T, Pemsel H, Zimmermann R (1999) Mutational analysis of the N-ras, p53, p16INK4a, CDK4, and MC1R genes in human congenital melanocytic naevi. J Med Genet 36(8): 610-614.
-
Sánchez-Laorden BL, Herraiz C, Valencia JC (2009) Aberrant trafficking of human melanocortin 1 receptor variants associated with red hair and skin cancer: Steady-state retention of mutant forms in the proximal golgi. J Cell Physiol 220(3): 640-654.
-
Flanagan N, Healy E, Ray A (2000) Pleiotropic effects of the melanocortin 1 receptor (MC1R) gene on human pigmentation. Hum Mol Genet 9(17): 2531-2537.
-
Motokawa T, Kato T, Hashimoto Y et al. (2007) Effect of Val92Met and Arg163Gln variants of the MC1R gene on freckles and solar lentigines in Japanese . Pigment Cell Res 20(2): 140-143.
-
yamaguchi K, Watanabe C, Kawaguchi A (2012) Association of melanocortin 1 receptor gene (MC1R) polymorphisms with skin reflectance and freckles in Japanese. J Hum Genet 57(11): 700-708.
-
suryaningsih BE, Sadewa AH, Wirohadidjojo YW (2019) Association between heterozygote Val92Met MC1R gene polymorphisms with incidence of melasma: a study of Javanese women population in Yogyakarta .Clin Cosmet Investig Dermatol 12: 489-495.
-
Bastiaens M, Ter huurne J, Gruis N (2001) The melanocortin-1-receptor gene is the major freckle gene. Hum Mol Genet 10(16): 1701-1708.
-
Vallone MG, Tell Marti G, Potrony M (2018) Melanocortin 1 receptor (MC1R) polymorphisms’ influence on size and dermoscopic features of nevi . Pigment Cell Melanoma Res 31(1): 39-50.
-
kennedy C, Bajdik CD, Willemze R (2003) The influence of painful sunburns and lifetime sun exposure on the risk of actinic keratoses, seborrheic warts, melanocytic nevi, atypical nevi, and skin cancer. J Invest Dermatol 120(6): 1087-1093.
-
Hu HH, Benfodda M, Dumaz N (2014) A large French case-control study emphasizes the role of rare Mc1R variants in melanoma risk. Biomed Res Int pp: 925-716.
-
Duffy DL, Lee KJ, Jagirdar K (2019) High naevus count and MC1R red hair alleles contribute synergistically to increased melanoma risk. Br J Dermatol 181(5): 1009- 1016.
-
Van Der Poel LAJ, Bergman W, Gruis NA (2020) The role of MC1R gene variants and phenotypical features in predicting high nevus count. Melanoma Res 30(5): 511- 514.
-
Whitton M, Pinart M, Batchelor JM (2016) Evidence- based management of vitiligo: summary of a Cochrane systematic review. Br J Dermatol 174(5): 962-969.
-
Wang JY, Chen H, Wang YY (2017) Network pharmacological mechanisms of Vernonia anthelmintica (L.) in the treatment of vitiligo: Isorhamnetin induction of melanogenesis via up-regulation of melanin- biosynthetic genes. BMC Syst Biol 11(1): 103.
-
Yuan X, Meng D, Cao P (2019) Identification of pathogenic genes and transcription factors in vitiligo. Dermatol Ther 32(5): e13025.
-
Széll M, Baltás E, Bodai L (2008) The Arg160Trp allele of melanocortin-1 receptor gene might protect against vitiligo. Photochem Photobiol 84(3): 565-571.
-
Disse M, Reich H, Lee PK (2016) A Review of the Association Between Parkinson Disease and Malignant Melanoma. Dermatol Surg 42(2): 141-6.
-
Elincx-Benizri S, Inzelberg R, Greenbaum L (2014) The melanocortin 1 receptor (Mc1r) variants do not account for the co-occurrence of Parkinson’s disease and malignant melanoma. J Mol Neurosci 54(4): 820-825.
-
Gao X, Simon K C, Han J (2009) Genetic determinants of hair color and Parkinson’s disease risk. Ann Neurol 65(1): 76-82
-
Tell Marti G, Puig Butille JA, Potrony M (2015) The MC1R melanoma risk variant p.R160W is associated with Parkinson disease. Ann Neurol 77(5): 889-894.
-
Gao X, Simon KC, Han J (2009) Family history of melanoma and Parkinson disease risk. Neurology 73(16): 1286-1291.
-
Shi CH, Wang H, Mao CY (2016) MC1R variants in Chinese Han patients with sporadic Parkinson’s disease. Neurobiol Aging 42: 217.e215-216.
-
D’orazio JA, Nobuhisa T, Cui R (2006) Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature 443(7109): 340-344.
-
Langendonk JG, Balwani M, Anderson KE (2015) Afamelanotide for Erythropoietic Protoporphyria. N Engl J Med 373(1): 48-59.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind