Bee Diversity in an Peri-Urban Patch of “Restinga”
Bees gather food resources from angiosperm flowers, promoting pollination, cross-breeding, and increasing the genetic variability of many plant species. However, all over the world there has been a great decline in bee populations, with the most accepted hypotheses being the loss of habitat caused by urbanization, fungal contamination and pesticide residues. The collapse of bee colonies can unbalance ecosystems causing a reduction in plant abundance and directly impacting the agricultural economy. Peri-urban paths are considered refuges capable of sheltering part of the biodiversity, including communities of pollinators such as bees. On the Brazilian Atlantic coast, restinga ecosystems are fragmented and their remnants are threatened by anthropic actions and real estate speculation. In this work, the bee fauna was inventoried in a peri-urban patch of “restinga” in the Municipal Natural Park of Barreto’s Restinga in Macaé, Rio de Janeiro, Brazil, with the objective of understanding the structure of the bee community and its relationship with the availability of floral resources and seasonality. Collections were carried out monthly between February 2019 and February 2020. The methods included active collections with entomological nets between 08:00 and 17:00 hours. 363 individuals belonging to 31 bee species were sampled. The most abundant genera were Xylocopa, Eulaema and Eufriesia, with the highest richness and abundance during the summer. There was a significant relationship between the availability of floral resources and the richness and abundance of bees. There was also a positive relationship between bee richness and rainfall. The data generated may contribute to support public policies that ensure the maintenance and expansion of “restinga” areas in urban environments, contributing to the conservation of bee species in the Atlantic Forest.
Introduction
The Atlantic Forest is one of the six Brazilian biomes, being considered a national heritage and a diversity hotspot, that is, with a high degree of endemic species [1]. After several exploration cycles, the Atlantic Forest has become one of the most threatened and fragmented biomes in the world [2]. Among the physiognomies that make up this biome, we have the “restinga” formations, which are characterized by a predominantly shrubby and heterogeneous vegetation under a sandy soil, with low capacity to retain water, high incidence of solar radiation, wind and salinity [3, 4, 5].
The restingas shelter part of the biodiversity of the Atlantic Forest, therefore, they are priority areas in preservation actions [1, 6]. In addition, because it is a coastal environment, they are constantly threatened by the loss and fragmentation of habitats resulting from accelerated urban growth, port activities, real estate speculation and mineral extraction [7]. The size of the remaining restinga fragments and the management effort can influence the maintenance of local biodiversity Sofia, et al. [8] allowing the permanence of local fauna [2], such as rodents Cerqueira, et al. [9], lizards [10], birds Cogliatti-Carvalho, et al. [11] and insects, such as pollinating bees [12, 13].
Studies on the richness of bees in restingas in Brazil, although scarce, indicate the importance of this ecosystem for the maintenance of bee communities [14, 15, 16, 17, 18]. In a restinga region in the south of Bahia, 80 species of bees that visited Comolia ovalifolia bushes were recorded, in a sample area of 5 hectares, with the most predominant species being Centris, Xylocopa and Euglossa [19]. In the Lençóis Maranhenses National Park, 59 species of bees were sampled in a sandbank area of 200 hectares, the genera with the greatest richness being Megachile (12 spp.), Centris (10 spp.) and Xylocopa (5 spp) [20]. Still for sandbanks in northeastern Brazil, Silva, et al. [17] collected 80 species in an area of 450 hectares in Salvador, Bahia. In restingas in southern Brazil, in Santa Catarina, Kamke, et al. [21] inventoried 64 bee species in an area of approximately 40 hectares, while Patricio, et al. [22] sampled 39 species in an area of 18 hectares. In the state of Rio de Janeiro, in a restinga area in the north of the state of Rio de Janeiro, 16 bee species were collected only in the sandy cord and associated with the psammophile Canavalia rosea [13].
The composition of pollinator communities is related to the floral resources available in the environment, with greater richness and abundance occurring in periods when the availability of floral resources is greater [23, 24]. It was observed that the low productivity and availability of resources can influence the activity of bees, showing the importance of trophic relationships with the floral resource [23]. In addition, relationships also influence the overlapping of niches and possible competition between species (Nogueira-Ferreira and Augusto) [25], which may interfere with the frequency and timing of foraging in flowering plants [26].
Seasonal changes linked to radiation intensity, rainfall and temperature variation throughout the day and year are abiotic variables that influence the flowering intensity of plants (Lorenzon) and the richness and abundance of their pollinators [27]. In colder seasons, there is a lower supply of floral resources and a decrease in bee activity [27, 28, 29]. Throughout the day, pollinators can share the use of floral resources by modifying their foraging periods due to the influence of temperature and luminosity [30]. In Euglossinae bees, greater activity was observed in the morning, with an average temperature of 26.5°C [31]. Stingless bees and species with large body sizes also show greater activity in the early morning, when the temperature and luminosity are lower and the availability of resources is greater, while small bees start their activities when the temperature increases, around 11 am [32].
Bees are important pollinating agents, ensuring the reproduction of many plant species [33, 34]. Pollination increases the quantity and quality of fruits and grains that make up the diet of many animals, including 70% of crops intended for human consumption [35, 36]. In this sense, pollination by bees is fundamental to the economic success of several agricultural crops in the world [37]. Although the relevance of pollination ecosystem services is considered, with the approval of a series of international initiatives in defense of pollinators [38, 39, 40, 41], in recent years the rates of reduction in the abundance of several species have been alarming. Among the most likely causes for this worldwide phenomenon is the use of agricultural pesticides and the loss of habitats and resources, resulting from accelerated and disorderly urban growth [42, 43].
Thus, the creation and maintenance of urban parks could reduce the effects of degradation on the restinga fragments, making them refuges for bee species, thus minimizing the impacts of urbanization. When transforming a fragment of urban vegetation into a conservation unit, it is important to know its biological diversity so that management plans are well structured in order to promote the conservation of resources and ecosystem services [44]. In this sense, this study aims to inventory the community of bees in the Municipal Natural Park of Barreto’s Restinga, an peri-urban patch of restinga and analyze how the climatic seasonality influences the composition of this community.
Material and Methods
Study Area
The study was conducted in a restinga area in the Municipal Natural Park of Barreto’s Restinga - PNMRB - (22°20’22’’S and 41°44’28’’W) located in the municipality of Macaé, State of Rio de Janeiro, Brazil (Figure 1). The park has a peri-urban patch of restinga of approximately 32 hectares. The vegetation is predominantly shrubby mainly composed of species of Myrtaceae, Convolvulaceae, Cactaceae, Bignoneaceae and a sandy cord with a wide distribution of psammophilous vegetation. The climate in the region is tropical, classified as Aw according to Köppen [45].
The average temperature is 22.9°C and the annual average rainfall is 1126 mm, with most of the rain concentrated in the summer.
Sampling
Monthly collections were carried out between February 2019 and February 2020. The collections were carried out along a transect of 1500 meters. In each sampling, two collectors walked the transect from 8:00 am to 5:00 pm, staying for 10 minutes on each flowering plant. A total of 108 hours of field sampling were carried out. Bees were collected with entomological nets during visits to flowers and then killed in chambers containing ethyl acetate. All individuals were stored in envelopes with date and time identification, taken to the Integrated Laboratory of Invertebrate Biology of the Institute of Biodiversity and Sustainability of the Federal University of Rio de Janeiro (NUPEM/UFRJ), where they were mounted on an entomological pin and deposited in the collection. Species identification was performed using taxonomic keys and consulting specialists [46].
Climatic data of temperature, solar radiation and rainfall were recorded for each collection day (INMET – Instituto Nacional de Meteorologia) to assess how abiotic factors may influence the composition of the bee assemblage [47, 48].
Statistical Analysis
Data were analyzed in the statistical software R (R Development Core Team) using simple linear models. Analysis of Variance (ANOVA) was applied to verify whether the richness and abundance of bees vary according to the time of year. Analysis of variance was also carried out between abiotic factors (temperature, precipitation and solar radiation) and their relationship with the abundance and richness of bees. In order to verify if there was a temporal partition of floral resources between bee species, the Beta Diversity analysis was carried out (Betapart Package).
Results
A total of 363 bees belonging to 31 species were collected (Table 1). The most abundant genera were Xylocopa (177), Eulaema (48), Eufriesia (45), Epicharis (33) and Centris (30), which together make up 91.7% of the specimens collected.
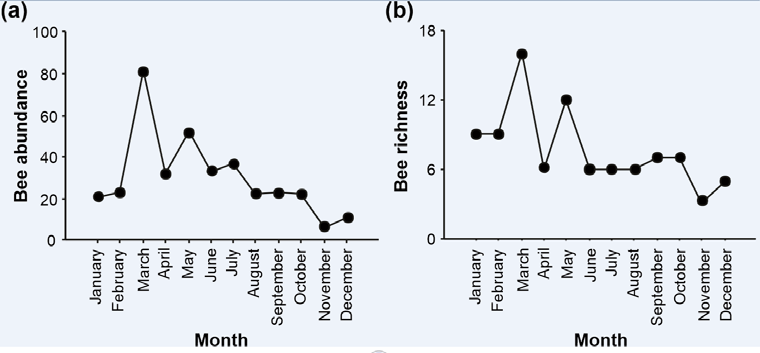
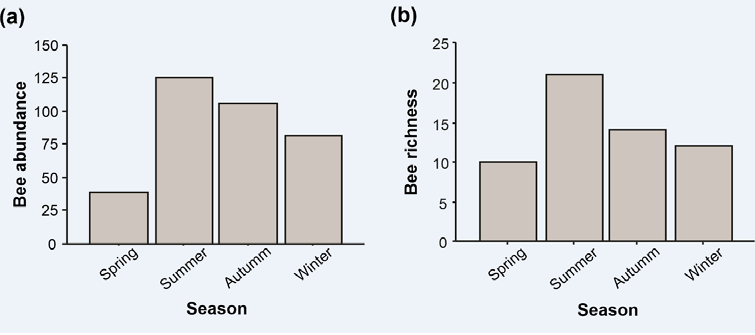
The month with the highest abundance and richness was March, with the lowest values occurring in November (Table 1 & Figure 2). The seasons with the highest abundance and richness of bees were summer and autumn (Figure 3).
| Abundance relative to the months sampled 2019/2020 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Specie | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Jan | Feb | |
| 1- Apis mellifera | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 2- Centris sp. 1 | 3 | 1 | 4 | |||||||||||
| 3- Centris sp. 2 | 1 | 1 | 3 | 5 | ||||||||||
| 4- Centris sp. 3 | 2 | 1 | 1 | 4 | ||||||||||
| 5- Centris sp. 4 | 4 | 4 | ||||||||||||
| 6- Centris sp. 5 | 3 | 3 | ||||||||||||
| 7- Centris sp. 6 | 2 | 3 | ||||||||||||
| 8- Centris sp. 7 | 2 | 1 | 3 | |||||||||||
| 9- Centris analis | 2 | 1 | 1 | 1 | 5 | |||||||||
| 10- Ceratina sp. 1 | 1 | 1 | ||||||||||||
| 11- Ceratina sp. 2 | 1 | 1 | ||||||||||||
| 12- Epicharis sp. 1 | 2 | 3 | 3 | 3 | 6 | 1 | 2 | 2 | 1 | 23 | ||||
| 13- Epicharis sp. 2 | 4 | 2 | 1 | 3 | 10 | |||||||||
| 14- Eufriesea sp. 1 | 1 | 1 | ||||||||||||
| 15- Eufriesea dentilabris | 9 | 14 | 8 | 6 | 7 | 44 | ||||||||
| 16- Euglossa sp. 1 | 1 | 1 | ||||||||||||
| 17- Eulaema nigrita | 19 | 10 | 3 | 4 | 7 | 2 | 45 | |||||||
| 18- Eulaema nigrohirta | 2 | 1 | 3 | |||||||||||
| 19- Megachile sp1 | 2 | 2 | 5 | |||||||||||
| 20- Megachile sp2 | 1 | 1 | ||||||||||||
| 21- Melipona quadrifasciata | 1 | 1 | ||||||||||||
| 22- Mesoplia sp.1 | 1 | 2 | 3 | |||||||||||
| 23- Pseudoaughlora sp. 1 | 1 | 1 | 1 | 3 | ||||||||||
| 24- Pseudoaughlora sp. 2 | 1 | 1 | ||||||||||||
Table 1: Diversity of bees and number of plant morphospecies collected over the months in the Municipal Natural Park of Table 1:
Table 1: Diversity of bees and number of plant morphospecies collected over the months in the Municipal Natural Park of Table 1: Diversity of bees and number of plant morphospecies collected over the months in the Municipal Natural Park of Barreto’s Restinga, Macaé-RJ between the years 2019-2020. The abundance of Apis mellifera was not counted due to the large number of individuals (greater than 100 individuals/month).
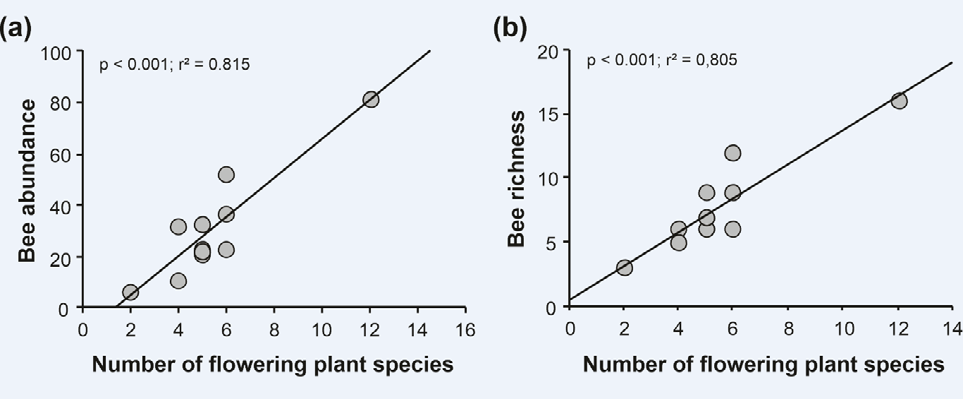
The months with the greatest diversity of flowering plants (March and May) were accompanied by the greatest richness and abundance of bees (Table 1). Significantly positive linear relationships were observed between the abundance of bees and the number of flowering species, where the predictor variable explained about 81% of the variation observed in the response variable (Figure 4a). We also found a significant positive linear relationship between bee richness and the number of flowering species, with the predictor variable explaining about 80% of the observed variation in the response variable (Figure 4b).
The bees collected floral resources at different times of the day, with the highest frequency of visits being between 9 and 11 am, and at 12 noon the lowest number of individuals sampled in flowers (Figure 5).
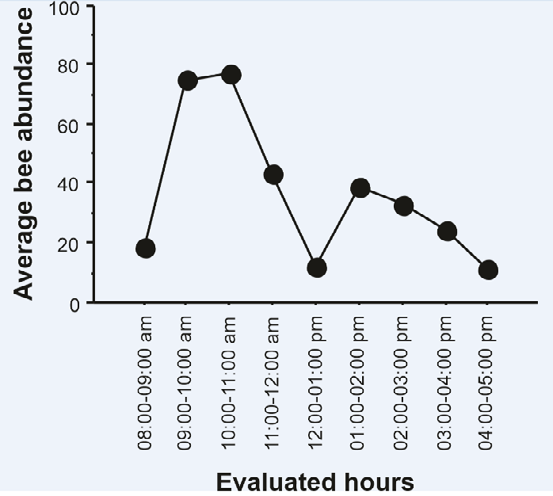
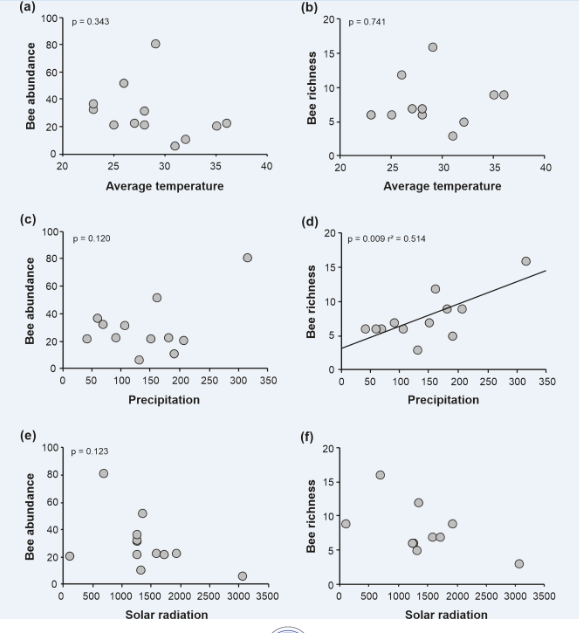
Figure 6: Relationship between abundance and richness of bees with different environmental variables, in the Municipal Natural Park of Barreto’s Restinga, Macaé, RJ, between 2019 and 2020. Average temperature (0C), precipitation (mm) and solar radiation (lux) The solar radiation indices on the sampling days showed two large peaks, with the lowest solar radiation in March, where the highest monthly abundance (81) and richness (16) of bees occurred, and the highest radiation index was in the month of November, when only 06 individuals belonging to 03 bee species were sampled. However, the abiotic variables temperature, rainfall and solar radiation were not significantly related to the abundance of bees. Also, no significant linear relationship was found between bee richness and average temperature and solar radiation. However, a significantly positive linear relationship was observed between bee richness and rainfall (r² = 0.514, p = 0.009; Figure 6).
Discussion
Apidae was the most representative group in this work, with the genera Xylocopa, Eulaema and Eufriesia being the most diverse, as also observed in other restinga areas [17, 22, 49, 50]. The prevalence of species of these genera in restinga can be related to generalist habits in the use of floral resources and body size, since most species of the genera in question have a medium to large body size [51] which enables them to support the strong and constant winds in the restinga [52].
Restinga areas in northeastern Brazil showed higher richness than that observed for the PNMRB. In southern Bahia, Oliveira-Rebouças, et al. [19] recorded 80 bee species. In Lençóis Maranhenses, 59 species were sampled Gostinsk, et al. [20] and in Salvador, Bahia, 80 species were collected [17]. In a restinga in southern Brazil, in Santa Catarina Patricio RS, et al. [22] sampled 39 species. The difference in richness can be attributed to several factors, such as collection time and areas of sampled fragments. However, within the cited works, the relative low richness of the current study can be attributed to factors of anthropic influences, since the area is highly urbanized in all its limits and with continuous use and traffic of people and vehicles.
The bees showed a temporal variation in foraging behavior throughout the day, with the highest foraging times being between 9 am and 11 am. This pattern is commonly observed for species of Xylocopa and Megachile when visiting flowers of Sophora tomentosa and Crotalaria vitellina [33]. This time preference has also been observed for the genera Mesoplia and Centris [49]. Insects need to reach a high metabolic rate to start foraging, but they tend to limit their activities to temperature ranges that have little influence on their energy expenditure [53].
The availability of floral resources influences the structure of the bee community, with greater abundance and richness being common in the months with greater supply of flowering plants [15, 29]. In the sampled study area, the Municipal Natural Park do Barretos Restinga, a work carried out in the sandy cord demonstrated that the richness and abundance of bees was positively related to the number of open flowers of Canavalia rosea [13]. Bees are sensitive to environmental responses associated with resource offers, and this factor may be a determining factor in community structure [54, 55]. Thus, climatic seasonality may have an effect on the availability of floral resources, such as the amount of flowers and the reward to visitors (nectar and pollen), which will influence the structure of the bee community [29, 34, 55].
During the hottest and rainiest months, a greater abundance of bees was observed, which may be related to the availability of water in the soil. In the restinga, the soils do not accumulate water so easily and with that the development of the plants is limited in the driest months, while in the rainy periods there is a greater emission of leaf buds and flower buds, which means a greater supply of resources flowers in this period [56, 57]. In the study area, it was observed that as rainfall increased, there was a significant increase in bee species richness, which possibly occurred due to the increase in the supply of floral resources, but can also be explained as a response to less overlapping of niches and less competition for resources between different species [58].
As for the effects of solar radiation, a lower richness and abundance of bees foraging in the period of the year with the highest radiation (November) was observed, as was also observed by Souza [59] who correlated higher air temperature and solar radiation to a lower number of Melipona subnitida bees foraging. In the study area, the five bees that were observed foraging during this period of higher radiation, out of a total of six specimens observed, are species of Xylocopa. This genus is one of the most diverse in restinga areas, being considered a potential pollinator for several plants, because, due to their large body size, these bees are able to forage even under strong winds and high temperatures, which is an important adaptation for a pollinator in the restinga environment [19, 50].
Conclusion
Knowledge about the structure of bee communities and their ecosystem services can contribute to actions aimed at preserving remaining areas of the Atlantic Forest and its phytophysiognomies, such as restingas. Considering the accelerated process of urbanization and fragmentation of restinga environments, as occurred in the PNMRB, it is important to inventory and understand the trophic relationships between fauna and flora, generating subsidies for management plans that aim to maintain or expand these protected areas, ensuring the ecosystem conservation.
Our study demonstrated that even urban and anthropized fragments can harbor plant species and pollinators, being fundamental for the maintenance of sandbank species and, therefore, contributing to the conservation of the Brazilian Atlantic Forest.
Acknowledgments: The authors thank the Federal University of Rio de Janeiro (UFRJ) for the logistical support and YA thanks CNPq for the scholarship
References
-
Myers N, Mittermeier RA, Mittermeier CG, DaFonseca GA, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
-
Tabarelli M, Rod SA (2005) Desafios e oportunidades para a conservação da biodiversidade na Mata Atlântica brasileira. Megadiversidade 1: 132-138.
-
Holzer W, Crichyno J, Pires AC (2004) Sustentabilidade da urbanização em áreas de restinga: uma proposta de avaliação pós-ocupação. Paisagem e Ambiente 19: 49-65.
-
Instituto Brasileiro de Geografia e Estatística (2010) Censo demográfico.
-
Barcelos MEF, Riguete JR, Silva LTP, Ferreira PD (2012) Uma visão panorâmica sobre os solos das restingas e seu papel na definição de comunidades vegetais nas planícies costeiras do sudeste do Brazil. Natureza 10: 71-76.
-
Ministério do Meio Ambiente (2000) Avaliação e Ações prioritárias para a Conservação da Biodiversidade da Mata Atlântica e Campos Sulinos. Brazília: MMA/SBF.
-
Martins SE, Rossi L, Sampaio PSP, Magenta AG (2008) Caracterização florística de comunidades vegetais de restinga em Bertioga, SP, Brazil. Acta Botanica Brasilica 22(1): 249-274.
-
Sofia SH, Suzuki KM (2004) Comunidades de machos de abelhas Euglossina (Hymenoptera: Apidae) em fragmentos florestais no sul do Brazil. Neotropical Entomology 33(6): 693-702.
-
Cerqueira R, Fernandez FAS, Quintela MFS (1990) Mamíferos da restinga de Barra de Maricá, Rio de Janeiro. Papéis Avulsos de Zoologia 37(9): 141-157.
-
Rocha CFD (2000) Biogeografia de Répteis de Restingas: Distribuição, Ocorrência e Endemismos. In: Esteves FA, Lacerda LD (Eds.), Ecologia de restingas e lagoas costeiras. NUPEM/UFRJ, Macaé, Rio de Janeiro, Brasil.
-
Cogliatti-Carvalho L, Freitas AFN, Rocha FD, Sluys MV (2001) Variação na estrutura e na composição de Bromeliaceae em cinco zonas de restinga no Parque Nacional da Restinga de Jurubatiba, Macaé, RJ. Revista Brasileira de Botânica 24(1): 1-9.
-
Oliveira-Rebouças P, Gimenes M (2011) Potential pollinators of _Comolia_ _ovalifolia_ DC Triana (Melastomataceae) and _Chamaecrista ramosa_ (Vog.) HS Irwin and Barneby var. ramosa (Leguminosae- Caesalpinioideae), in restinga, Bahia, Brazil. Brazilian Journal of Biology 71(2): 343-351.
-
Meirelles J, Miranda SM, Soares MO, Antonini Y, Bacci L et al., (2022) Bee Diversity Associated with Canavalia rosea (Sw.) DC. (Fabaceae, Papilionoideae) in an Peri-Urban Patch of “Restinga”. International Journal of Zoology and Animal Biology 5: 000407.
-
Marques MCM, Oliveira PEAM (2004) Fenologia de espécies do dossel e do sub-bosque de duas Florestas de Restinga na Ilha do Mel, sul do Brasil. Brazilian Journal of Botany 27(4): 713-723.
-
Silva MCM, Martins CF (1999) Flora apícola e relações tróficas de abelhas (Hymenoptera: Apoidea) em uma área de restinga (Praia de Intermares, Cabedelo-PB, Brasil). Principia 7: 40-51.
-
Silva O, Rego MMC, Albuquerque PMC, Ramos MC (2009) Abelhas Euglossina (Hymenoptera: Apidae) em área de restinga do nordeste do Maranhão. Neotropical Entomology 38(2): 186-196.
-
Silva M, Ramalho M, Aguiar CM, Silva MD (2015) Apifauna (Hymenoptera, Apoidea) em uma área de restinga arbórea-mata atlântica na costa atlântica do Nordeste do Brasil. Magistra 27(1): 110-121.
-
Magnago LFS, Martins SV, Schaefer CEGR, Neri AV (2012) Restinga forests of the Brazilian coast: richness and abundance of tree species on different soils. Anais da Academia Brasileira de Ciências 84(3): 807-822.
-
Oliveira-Rebouças P, Gimenes M (2004) Abelhas (Apoidea) visitantes de flores de _Comolia ovalifolia_ DC Triana (Melastomataceae) em uma área de restinga na Bahia. Neotropical Entomology 33(3): 315-320.
-
Gostinski LF, Carvalho GCA, Rêgo MMC, Alburquerque PMC (2016) Species richness and activity pattern of bees (Hymenoptera, Apidae) in the restinga area of Lençóis Maranhenses National Park, Barreirinhas, Maranhão, Brazil. Revista Brasileira de Entomologia 60(4): 319- 327.
-
Kamke R, Zillikens A, Steiner J (2011) Species richness and seasonality of bees (Hymenoptera, Apoidea) in a restinga area in Santa Catarina, southern Brazil. Studies on Neotropical Fauna and Environment 46: 35-48.
-
Patricio RS, Pereira AS, Elias SG, Furlanetto CB, Cascaes MF, et al. (2014) Apifauna (Hymenoptera: Apidae) em área de restinga no sul de Santa Catarina, Brasil. Tecnologia E Ambiente pp: 20.
-
Gonçalves SJM, Rego MMC, Araújo A (1996) Abelhas sociais (Hymenoptera: Apidae) e seus recursos florestais em uma região de mata secundária, Alcântara, MA, Brasil. Acta Amazônica. 26(1-2): 55-68.
-
Araújo VA, Araújo APA, Antonini Y (2010) Impact of Resource Availability on Bee Diversity. Sociobiology 55(3): 1-13.
-
Nogueira-Ferreira FH, Augusto SC (2007) Amplitude de nicho e similaridade no uso de recursos florais por abelhas eussociais em uma área de cerrado. Bioscience Journal 23: 45-51.
-
Ribeiro EKMD, Rêgo MMC, Machado ICS (2008) Cargas polínicas de abelhas polinizadoras de Byrsonima chrysophylla Kunth. (Malpighiaceae): fidelidade e fontes alternativas de recursos florais. Acta Botanica Brasilica 22(1): 165-171.
-
Lorenzon MCA, Matrangolo CAR, Schoereder JH (2003) Flora visitada pelas abelhas ussociais (Hymenoptera, Apidae) na Serra da Capivara, em Caatinga do Sul do Piauí. Neotropical Entomology 32(1): 27-36.
-
Jamhour J, Laroca S (2004) Uma comunidade de abelhas silvestres (Hymenopter, Apoidea) de Pato Branco (PR-Brasil): diversidade, fenologia, recursos florais e aspectos biogeográficos. Acta Biológica Paranaense 33: 27-119.
-
Araújo VA, Antonini Y, Araújo A (2006) Diversity of bees and their floral resources at altitudinal areas in the Southern Espinhaço Range, Minas Gerais, Brazil. Neotropical Entomology 35(1): 30-40.
-
Roubik DW (1978) Competitive interactions between neotropical pollinators and Africanized honey bees. Science 201: 1030-1032.
-
Oliveira ML (1999) Sazonalidade e horário de atividade de abelhas Euglossinae (Hymenoptera, Apidae), em florestas de terra firme na Amazônia Central. Revista Brasileira de Zoologia 16: 83-90.
-
Teixeira LV, Campos FNM (2005) Início da atividade de vôo em abelhas sem ferrão (Hymenoptera, Apidae): influência do tamanho da abelha e da temperatura ambiente. Revista Brasileira de Zoociências 7: 195-202.
-
Brito VLG, Pinheiro M, Sazima M (2010) Sophora tomentosa e Crotalaria vitellina (Fabaceae): biologia reprodutiva e interações com abelhas na restinga de Ubatuba, São Paulo. Biota Neotropica 10(1): 185-192.
-
Garratt MPD, Coston DJ, Truslove CL, Lappage MG, Polce C, et al. (2014) The identity of crop pollinators helps target conservation for improved ecosystem services. Biological Conservation 169: 128-135.
-
Klein AM, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA (2006) Importance of pollinators in changing landscapes for world crops. Proceedings of the royal society B: biological sciences 274: 303-313.
-
Gallai N, Salles JM, Settele J, Vaissiere BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological economics 68: 810-821.
-
Willcox BK, Aizen MA, Cunningham SA, Mayfiel MM, Rader R (2017) Deconstructing pollinator community effectiveness. Current opinion in insect science 21: 98- 104.
-
Imperatriz-Fonseca VL, Saraiva AM, Gonçalves LS (2007) A iniciativa brasileira de polinizadores e os avanços atuais para a compreensão do papel dos polinizadores como produtores de serviços ambientais. Bioscience Journal 23: 100-105.
-
VanEngelsdorp D, Evans JD, Saegerman C, Mullin C, Haubruge E, et al. (2009) Colony Collapse Disorder: A Descriptive Study. PLoS ONE 4(8): e6481.
-
Pires S, Pereira CS, Lopes FMR, Nocelli MTF, Malaspina RC et al., (2016) Weakness and collapse of bee colonies in Brazil: are there cases of CCD? Pesquisa Agropecuária Brasileira 51(5): 422-442.
-
Nielsen A, Reitan T, Rinvoll AW, Brysting AK (2017) Effects of competition and climate on a crop pollinator community. Agriculture, Ecosystems & Environment 246: 253-260.
-
Andena SR, Bego LR, Mechi MR (2005) A comunidade de abelhas (Hymenoptera, Apoidea) de uma área de cerrado (Corumbataí, SP) e suas visitas às flores. Revista Brasileira de Zoociências 7(1): 55-91.
-
Kerr WE, Carvalho GA, Silva AC, Assis MGP (2001) Aspectos pouco mencionados da biodiversidade amazônica. Parcerias Estratégicas 12: 20-41.
-
Brito DMC (2008) Conflitos em unidades de conservação. PRACS: Revista Eletrônica de Humanidades do Curso de Ciências Sociais da UNIFAP 1: 1-12.
-
Alvares CA, Stape JL, Sentelhas PC, Leonardo GMJ, Sparovek GJMZ (2013) Köppen’s climate classification map for Brazil. 22(6): 711-728.
-
Silveira FA, Melo GAR, Almeida EAB (2002) Abelhas brasileiras. Sistemática e Identificação. Fundação Araucária, Belo Horizonte.
-
Instituto Nacional de Meteorologia do Brasil-Normas Climatológicas, Brasília-DF, Brasil.
-
R Development Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
-
Goes LP, Paz JRL, Silva WP, Silveira SRB, Neves EL, et al. (2021) Comunidades de abelhas em diversos métodos de amostragem: Fenologia e sazonalidade de uma restinga costeira em mangue seco, litoral norte da Bahia. Candombá-Revista virtual 1: 31-34.
-
Gimenes M, Oliveira-Rebouças P, Almeida GF (2007) Estudo das interações abelhas (Hymenoptera-Apoidae) e flores em um ecossistema de restinga no estado da Bahia, Brasil. SITIENTIBUS série Ciências Biológicas 7(4): 347-353.
-
Viana BF (1999) A Comunidade de abelhas (Hymenoptera, Apoidea) das Dunas Interiores do Rio São Francisco, Bahia, Brasil. Anais da Sociedade Entomológica do Brasil 28(4): 635-645.
-
Gottsberger G, Camargo JMF, Silberbauer-Gottsberger I (1988) A bee pollinated tropical community: The beach dune vegetation of Ilha de São Luís, Maranhão, Brazil. Botanische Jahrbücher fur Systematik 109(4): 469-500.
-
Proni EA, Nogueira MF, Macieira OJD (2002) Ritmo Circadiano da Taxa Respiratória de Operárias Isoladas de Plebeia droryana (Humenoptera: Apidae: Meliponinae), Durante o Inverno e Verão. Arquivos de Ciências Veterinárias e Zoologia da UNIPAR 5(1): 51-58.
-
Michener CD (2000) The bees of the world. The John Hopkins University Press, Baltimore.
-
Potts SG, Vulliamy B, Dafni A, Ne’eman G, Willmer P (2003) Linking Bees and Flowers: How do floral communities structure pollinator communities?. Ecology 84(10): 2628-2642.
-
Henriques R, Araujo D, Hay J (1986) Descrição e classificação dos tipos de vegetação da restinga de Carapebus, Rio de Janeiro. Revista Brasileira de Botânica 9(2): 173-189.
-
Rosado BHP, Mattos EA (2007) Variação temporal de características morfológicas de folhas em dez espécies do Parque Nacional da Restinga de Jurubatiba, Macaé, RJ, Brasil. Acta botânica brasílica 21(3): 741-752.
-
Menezes C, Silva CI, Singer RB, Kerr WE (2007) Competição entre abelhas durante forrageamento em Schefflera arboricola (Hayata). Bioscience Journal 23: 63-69.
-
Souza-Júnior JBF (2019) Termorregulação e atividade de forrageamento de melipona subnitida no bioma caatinga. Tese de Doutorado. UFERSA, Mossoró/ RN pp: 82.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework