Navigational Competency is Mediated by Foraging Experience and Light Conditions in the Nocturnal Bull Ant Myrmecia midas
The nocturnal bull ants of the genus Myrmecia, Myrmecia midas travel in both bright and dim settings on the ground and also vertically up on trees navigating towards targets. It is known that foragers retain multiple panoramic memories of terrestrial landmarks as well as celestial cues along their foraging route. It is likely that they can compare these stored views with their current visual input to return to their nest. Here we examined whether nocturnal M. midas foragers’ navigational efficiency and success at view-based navigation suffer at low light levels (night time) compared with daylight levels of brightness. When exposed to panoramic cues in dim conditions, which had dimmer light levels than their peak foraging period, they were less well oriented and less likely to find the nest, with the catchment area smaller at night than during the day. At different displacement distances from the nest, initial navigational behaviours differed. Foragers displayed more scanning near the starting point at shorter distances, while with increasing distance they switched more to loops of systematic searching or heading off in some direction without scans. In the dim conditions, landmark guidance became less effective, perhaps explaining why foragers limit their navigational activities mostly to the twilight hours, when light levels are still adequate, and mostly forage in a tree through the night.
Introduction
Like most central-place foragers, ants rely on a variety of strategies to navigate in their environment [1, 2, 3]. First, path integration (PI) is one major mechanism supporting the navigational success of ants, especially for those that inhabit areas with little to no prominent visual landmarks or are unfamiliar with their environment [1, 4]. Path integration enables ants to compute the direction home by a process equivalent to adding all increments of movement to obtain a continually updated representation of direction and distance from the starting point, information from which ants return on a direct path to their nest [4, 5, 6, 7, 8]. Secondly, ants use landmark-based navigation, especially in a visually cluttered environment. Ants acquire the visual panorama [9, 10, 11, 12] and information in other modalities (olfactory cues [13], tactile cues [14] magnetic cues [15], vibration cues [15] thermal cues [16]) from the surrounding to navigate habitual routes and to pinpoint the goal. When naïve ants first come out from the nest, they perform multiple learning walks around the nest entrance to learn the cues [17, 18, 19, 20]. The ants stop and display stereotypical orientation behaviours towards the nest and other locations during these learning walks to obtain landmark information to navigate back to home [19, 20, 21, 22]. When these two navigation strategies fail to return ants to their nest, which happens frequently due to navigational errors, ants undertake a systematic search as a back-up mechanism, forming loops around the starting point of the search and with loops increasing in size as the search continues [6, 23, 24, 25].
View-based models of ant navigation suggest that there should be some catchment area surrounding the nest where views collected during learning walks can direct the forager back to the nest [26, 27, 28, 29]. According to the view-based models, the catchment area of stored panoramic images is smaller in dense or cluttered environments where there are multiple landmarks blocking visual cues. In contrast, the catchment area is larger in open environments where distant landmarks are visible and are much more stable and catchment areas become systematically wider as the average distance of objects increases [30]. Foragers retain long-term memories of the visual cues of the nest panorama and may compare these retained views with their present views while they are away from the nest [31]. The ability of solitary foragers to generalise their views after learning visual information from a limited area has been studied in the desert ants Cataglyphis noda and Melophorus bagoti. The research showed that they were able to generalise their views to other distant scenes with stable landmark features and home from those never-visited locations after learning the nest panorama in an area extending 0.5m (C. noda [32] or 1m M. bagoti [33, 34]) around their nest. Like desert ants, night- active bull ants Myrmecia midas also memorise panoramic views and generalise those views up to 10.5m from their nest [35]. In the nocturnal ant, Myrmecia pyriformis, navigational performance and efficiency suffer at lower light levels [36, 37]. The subdued light levels during their active periods pose additional challenges for navigation. It is unclear whether M. midas foragers’ navigational efficiency change likewise with light levels. Further studies are required to understand navigational knowledge and view generalisation in bright and dim conditions in M. midas. Testing this ant living in a habitat with rich panoramic views would probe the generality of the experimental ethology of learning views.
Myrmecia foragers travel on the ground and also vertically up on trees to forage. Our study on the nocturnal bull ants M. midas examines how the navigational efficiency and catchment area of views suffer at low light levels (night time) compared with daylight levels of brightness. Individual M. midas foragers head out to a particular tree in the evening twilight when visual cues are less salient, either to their nest tree (< 30cm from the nest entrance) or to foraging trees (3 to 15m away) and return back to the nest in the morning twilight [38, 39, 40, 41, 42]. Therefore, the ants usually acquire and use scene information in dim conditions. We compared the initial orientations of the nocturnal bull ants in light and dim conditions at a range of distant displacement sites, in both ants that foraged on their nest tree and on those that foraged on other trees. We measured their ability to reach the destination, their nest, in such conditions and the ants’ route characteristics in these light conditions.
Methods
Study Subjects and Species
The night-active bull ants Myrmecia midas live in cluttered habitats, forage solitarily on nearby trees for insect prey and sugar sap, and do not show signs of recruitment by trail pheromones or contact with other foragers. To understand the navigational knowledge of M. midas, experiments were conducted at the Fauna Park, Macquarie University campus in Sydney from August 2020 to October 2020. Two nests were selected. From the first nest, only ants that foraged on the nest tree, which was 20cm away to the South from the nest (most of the foraging force), were selected, whereas from the second nest, only ants that foraged on a nearby tree 7 m away to the West (most of the foraging force) were selected. Both nest-tree and foraging-tree foragers were tested in both dim and daylight conditions. Ants usually came out from each nest and travelled straight to their foraging trees in the evening twilight and returned in the morning twilight. They were captured in the dusk 7pm – 8pm at the base of their foraging- or nest-tree trunk using foam-stoppered transparent glass tubes. After being collected, ants were kept in the laboratory overnight and fed a small amount of honey water. Later, ants were subjected to displacement experiments. After completing the experiment ants were painted with a tiny dot of yellow colour on their abdomen to avoid repeated testing. Australia does not have any ethical requirements for working with ants and in addition, the experimental procedures were completely non-invasive and as with previous behavioural research, they had no notable adverse effects on the individuals or on the colonies.
Navigational Efficiency of the Nest-Tree Foragers in day and Dim Light Conditions
To determine the navigational efficiency and catchment area of the nest-tree foragers in different light conditions, we tested the ants in two separate time periods: after sunrise, between 6am and 10am in the morning after capture (daylight condition) or between 9pm and 12am the next evening after the sunset (dim condition). The dim testing period was later (and hence dimmer) than the peak foraging period of the species, which is shortly after sunset. The field site, however, lay on campus near a suburban street, so that some human-made lights from the campus and from the street provided dim illumination through the night. From the nest entrance, we marked distant points 5 m, 10 m, and 20 m to the North, South, East and West directions (Figure 1). In the daylight condition, we conducted tests at 5m, 10m, and 20m distances. In the dim condition we tested only at 5m and 10m distances from the 4 different directions. At each displacement site n ≥ 15 ants were released individually. Each ant was tested only once. To quantify the compass direction of the initial headings, ants were released on a goniometer individually, a circle 45cm in radius drawn on a wooden board which was separated into 24 15° wedeges. We made a circular indentation in the centre with a 15mm diameter and 25mm depth to release the ants into. After releasing a forager at the centre of the goniometer we covered the hole for 30 seconds to allow the ant to settle. The first crossing observed at 45 cm was considered as the initial heading direction and the wedge that the ant crossed was recorded manually.
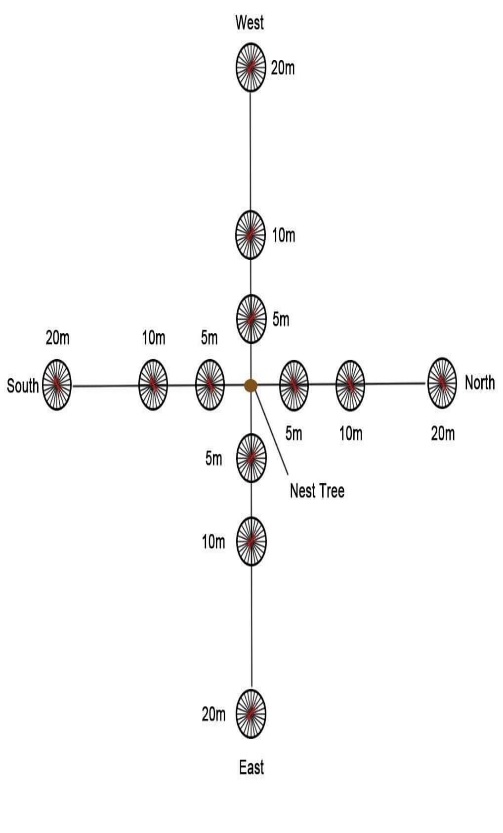
We also recorded the ants’ paths within a 45cm radius by using a video camera (GoPro HERO7) at each displacement site in the daylight condition. (The camera was not suitable for night conditions.) The camera was mounted on a tripod about 60cm away and pointed at the goniometer. The HD video camera recorded the nest area of about 60×50cm at 60 frames per second. This video camera recording was remotely operated via GoPro voice remote.
We quantified the proportions of ants that exhibited what we called Immediate Scanning, Systematic Searching and Direct Navigation on tests. In Immediate Scanning, an ant coming out from the displacement hole onto the goniometer platform showed stereotypical orientations with pauses to look at some portion of the panorama, within 5 cm of the release point, before heading off in a chosen direction without looping back. When ant foragers coming onto the goniometer performed scanning and then made one or more loops that brought the forager back to the starting point of the loop on the displacement board, that behaviour was considered Systematic Searching. The rest, ant foragers that headed off in a direction without any scans near the start and without looping back, was considered as showing Direct Navigation. After an ant left the goniometer, we followed it for up to 20 minutes. If the ant reached halfway to the nest within 20 minutes, we continued to follow the ant until it reached the nest (which all ants that reached this point eventually did). These ants were counted as successful. If the ant was unable to get to half-way to the nest within 20 minutes, it was deemed unsuccessful. Such ants were captured and released at the nest, after which all ants entered their nest.
Data Analysis
We separated ants that showed each type of initial navigational behaviour (Immediate Scanning, Systematic Searching, and Direct Navigation) from the video tapes for detailed analyses from first appearance until the ant first crossed the goniometer boundary. We generated the x-y co- ordinates of each ant’s path by using a customized Python program. We used OpenCV-based CSRT (Channel and Spatial Reliability Tracking) and Lucas-Kanade Optical Flow to track the x-y coordinates of the ant path in the video. Based on the video coordinates we calculated the path straightness of each path, defined as the ratio of the straight-line distance from the releasing point to the end point of the path that the ant travelled on the goniometer to the forager’s total path length. For each nest, a two-way ANOVA was conducted to examine the influence of two independent variables (distance and initial behaviour) on the path straightness of foragers.
We compared success rate and initial navigation behaviour with Fisher’s exact tests.
Ants’ initial orientations on displacement tests were analyzed with circular statistics using the statistical program Oriana Version 4 (Kovach Computing Services™). To test for a uniform distribution of headings (p > 0.05) for each condition, a Rayleigh’s Test was conducted. To test if initial orientations were significantly clustered around the nest direction at 0°, we examined if 0° fits within the 95% confidence interval (CI) of orientations (Watson tests) and conducted V tests, with alpha set at p = 0.05. A V test delivers a significant result when a distribution of headings is notably clustered around a specified target direction.
Results
Foraging Tree Foragers’ Behaviour in Daylight and Dim Conditions
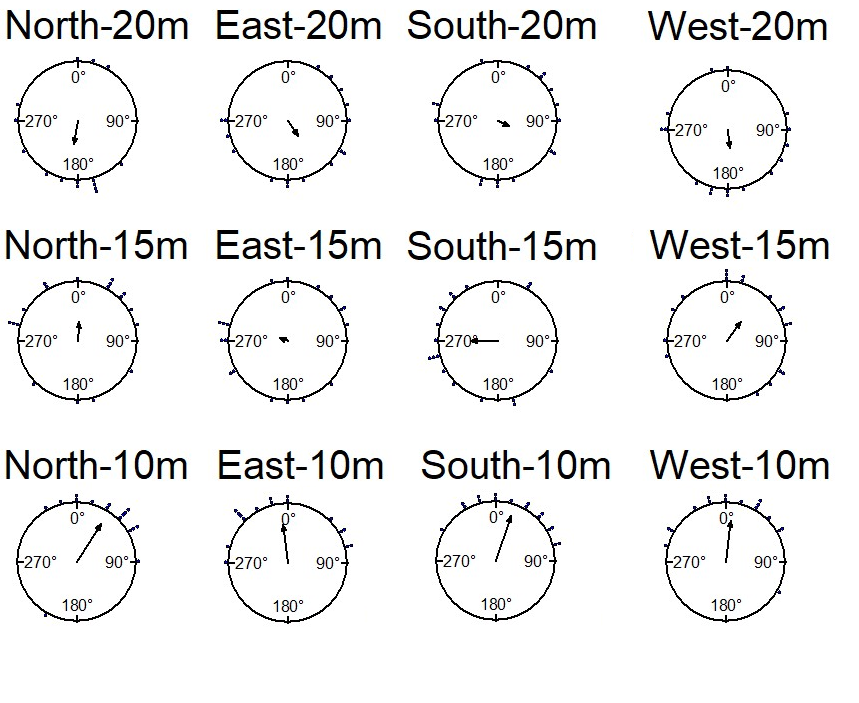
The foraging-tree foragers were collected in the evening twilight and were displaced at 10m, 15m, and 20m in 4 cardinal directions, each tested individually on the goniometer at one particular location in the early morning in daylight. The goniometer paths show that the distributions of initial (45cm) headings at 10m displacement sites were oriented towards the home vector direction (Watson test, E:p<0.05,W:p<0.05, N:p<0.05, S:p<0.05). At 15m sites ants oriented towards the nest only from the North and West directions (E:p>0.05,W:p<0.05, N:p<0.05, S:p>0.05; Figure 2, Table 1), West being the direction of the foraging tree. By contrast, all distributions of headings were uniformly distributed at 20m displacement sites (Rayleigh tests all locations p>0.05; Table 1). The V test showed that the initial orientations of ants were significantly towards the nest direction in all 10m displacement tests and in 15m North and West displacement tests (Table 1).
| Test | Mean vector | 95% confidence intervel | Rayleigh test | V test detection 00 | |||
|---|---|---|---|---|---|---|---|
| µ(0) | Minus(0) | Plus(0) | Z | P | Z | P | |
| Nest- Tree foragers displacement | |||||||
| 5m North | 3.55° | 336.49° | 30.6° | 6.76 | < 0.001 | 3.67 | < 0.001 |
| 5m South | 11.8° | 347.25° | 36.35° | 7.78 | < 0.001 | 3.86 | < 0.001 |
| 5m West | 49.92° | 269.69° | 190.15° | 5.71 | < 0.001 | 2.98 | < 0.001 |
| 5m East | 355.13° | 309.16° | 41.10° | 10.8 | < 0.001 | 3.56 | < 0.001 |
| 10m North | 352.74° | 308.61° | 36.86° | 2.96 | 0.05 | 2.41 | 0.007 |
| 10m South | 163.31° | 232.09° | 94.53° | 0.07 | 0.93 | -0.4 | 0.64 |
| 10m West | 335.37° | 276.93° | 33.82° | 1.74 | 0.17 | 1.7 | 0.04 |
| 10m East | 168.02° | 93.89° | 243.95° | 1.09 | 0.34 | -1.4 | 0.92 |
| 20m North | 202.52° | 172.01° | 234.03° | 5.32 | 0.004 | -3 | 0.99 |
| 20m South | 221.03° | 173.69° | 268.37° | 2.6 | 0.07 | -1.7 | 0.95 |
| 20m West | 151.97° | 102.77° | 201.16° | 2.42 | 0.087 | -1.9 | 0.97 |
| 20m East | 137.82° | 97.46° | 178.14° | 3.48 | 0.028 | -2 | 0.97 |
| Foraging Tree foragers displacement | |||||||
| 10m North | 32.74° | 11.27° | 54.22° | 8.82 | < 0.001 | 3.53 | < 0.001 |
| 10m South | 18.53° | 358.82° | 38.23° | 9.67 | < 0.001 | 4.17 | < 0.001 |
| 10m West | 7.09° | 342.43° | 31.75° | 7.44 | < 0.001 | 0.67 | < 0.001 |
| 10m East | 351.88° | 324.73° | 19.04° | 6.51 | < 0.001 | 3.57 | < 0.001 |
| 15m North | 4.08° | 305.65° | 62.51° | 1.75 | 0.17 | 1.86 | 0.031 |
| 15m South | 269.2° | 229.09° | 309.3° | 3.51 | 0.02 | -0 | 0.51 |
| 15m West | 337.78° | 351.57° | 84° | 2.72 | 0.06 | 1.84 | 0.03 |
| 15m East | 168.02° | 156.° | 16.92° | 0.33 | 0.72 | 0.31 | 0.37 |
| 20m North | 190.61° | 135.49° | 245.74° | 2.33 | 0.09 | -2.1 | 0.98 |
| 20m South | 114.42° | 16.92° | 211.91° | 0.65 | 0.5 | -0.5 | 0.68 |
| 20m West | 171.73° | 108.73° | 238.73° | 1.42 | 0.24 | -1.7 | 0.95 |
| 20m East | 146.94° | 83.01° | 210.86° | 1.48 | 0.23 | -1.4 | 0.92 |
| Night Nest Tree foragers displacement | |||||||
| 5m North | 222.57° | 179.29° | 265.86° | 3.19 | 0.041 | -1.9 | 0.96 |
| 5m South | 42.38° | 2.14° | 82.63° | 3.61 | < 0.02 | 1.98 | < 0.02 |
| 5m West | 316.34° | 278.71° | 353.97° | 4.14 | < 0.01 | 2.08 | < 0.01 |
| 5m East | 222.81° | 143.54° | 302.05 | 0.97 | 0.38 | -1 | 0.84 |
| 10m North | 19.01° | 309.05° | 88.97° | 1.24 | 0.29 | 1.48 | 0.06 |
| 10m South | 234.79° | 144.8° | 324.77° | 0.76 | 0.47 | -0.7 | 0.75 |
| 10m West | 143.35° | 102.05° | 184.65° | 3.3 | 0.03 | -2.1 | 0.98 |
| 10m East | 71.38° | 286.35° | 216.42° | 0.29 | 0.74 | 0.24 | 0.4 |
| Night-Foraging Tree foragers displacement | |||||||
| 10m North | 225.81° | 1.59° | 62.72° | 0.18 | 0.83 | -0.4 | 0.66 |
| 10m South | 62.85° | 4.5° | 121.21° | 2.14 | 0.11 | 0.94 | 0.17 |
| 10m West | 163.51° | 96.98° | 230.03° | 1.77 | 0.17 | 1.8 | 0.96 |
| 10m East | 32.15° | 1.59° | 62.72° | 5.45 | < 0.003 | 2.79 | < 0.002 |
Table 1: Statistical results for initial heading direction in all conditions in dispacement tests of nest-tree foragers and forag
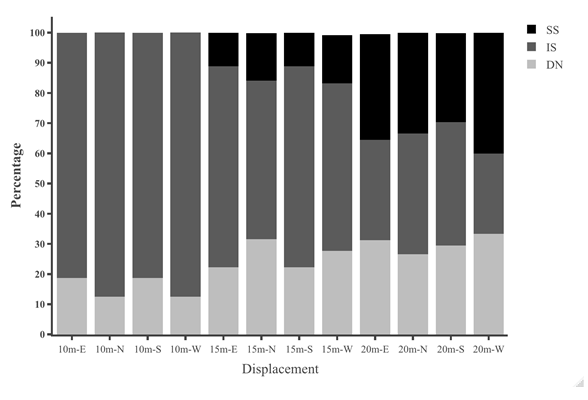
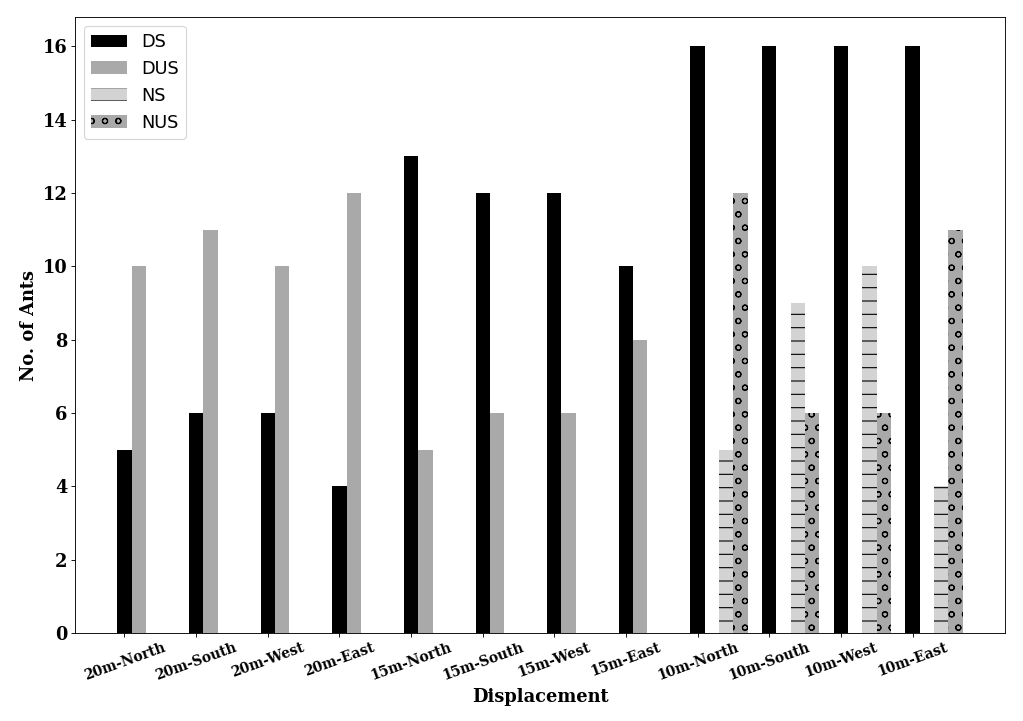
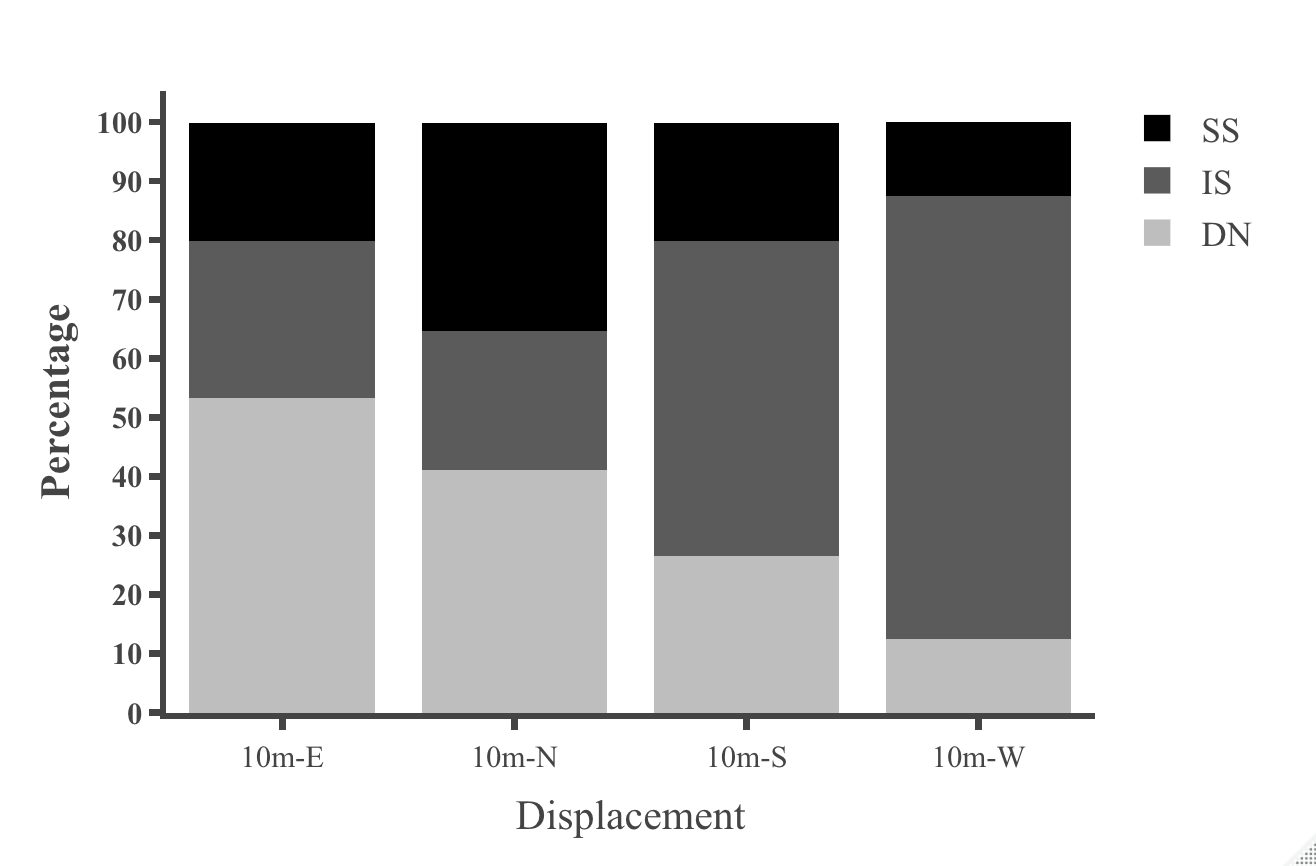
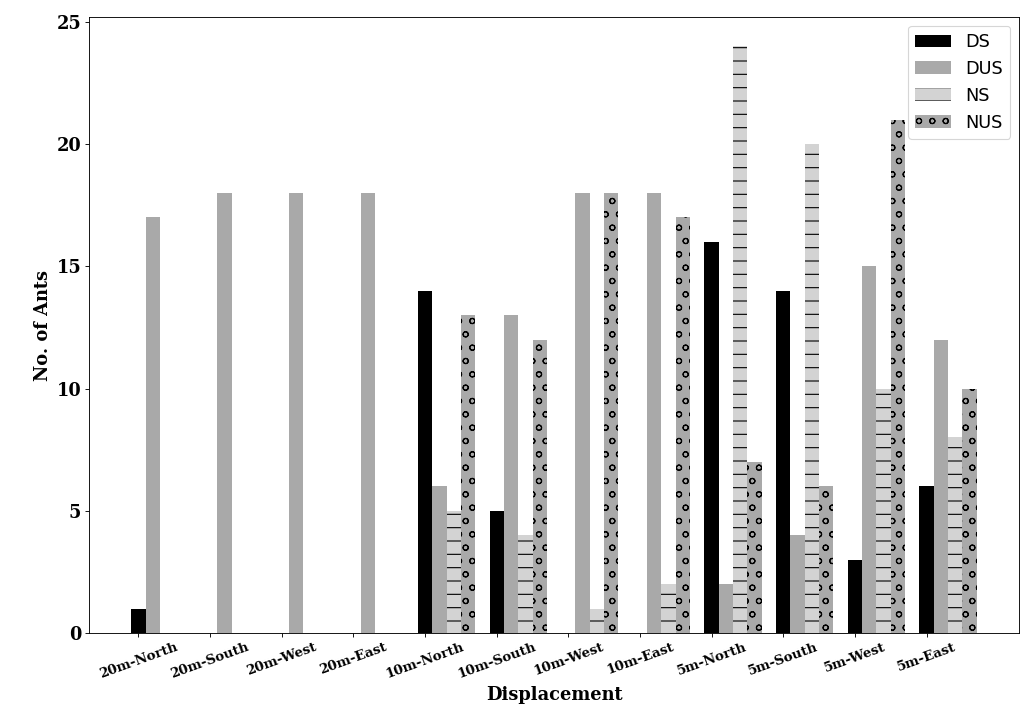
Table 1: Statistical results for initial heading direction in all conditions in dispacement tests of nest-tree foragers and foraging- tree foragers Categorising the ants behaviours within the 45cm radius using the video recordings shows that at 10 m, the majority of the foragers in the daylight condition perform Immediate Scanning (IS), while at greater distances more ants use Systematic Searches (SS) or Direct Navigation (DN) (Figure 3). The rate of successful returns also decreased as displacement distance grew (Figure 4, Fisher’s exact test, p = 0.001). While we have not recorded their paths and scans along the routes, our observations suggest that the ants walked faster and straighter in daylight conditions than in dim conditions (described next) and chose a heading direction more quickly at the shorter distances. With increasing distance, they stopped more frequently and searched a broader area to find the home route.
The foraging tree foragers were collected at evening twilight and also displaced the following night at 10m displacement sites (Figure 5, Table 1). Only those from the East oriented significantly towards the nest (Table 1, V test) and were nonrandomly distributed by the Rayleigh test (Table 1).
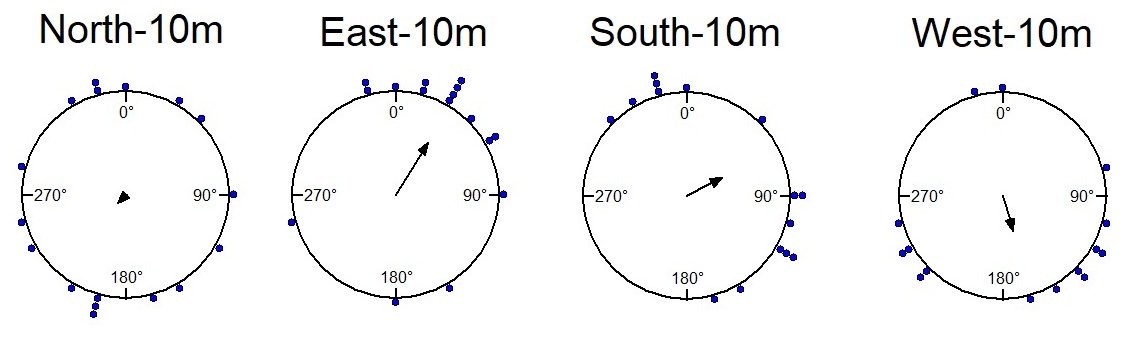
We also examined the proportions of Systematic Searching, Immediate Scanning, and Direct Navigation at 10m displacement sites. In the dim condition the proportion of individual ants exhibiting different initial navigational behaviours differed from those of ants tested in the daylight condition. Immediate Searching decreased relative to a Systematic Search pattern or Direct Navigation (Figure
6, Fisher’s exact test, p = 0.006). In dim conditions ants appeared to walk more slowly and stop more frequently compared to the daylight condition. Ants took more time to move on from the displacement site in the dim condition and paths appeared less straight than in the daylight condition. In addition, ants were less successful in reaching home in the dim condition compared to the daylight condition (Figure 9).
The path straightness of foraging-tree foragers on the goniometer at different displacement sites was measured. The foraging-tree foragers which performed Direct Navigation displayed a straighter path (Figure 10) compared to those exhibiting Immediate Scanning and Systematic Search. There was a significant interaction between the distance and the initial behaviour of the foraging-tree foragers, which appears to be due to foragers changing their navigational mode from Direct Navigation or Immediate scanning to Systematic search when displaced further away from the nest or foraging tree (F = 199.9, df = 4,193, p = 0.005). The two-way ANOVA analysis also showed a statistically significant difference in path straightness with the distance (F= 4.5, df = 4,193, p = 0.01) and across different initial navigational behaviours (F =395.84, df = 4,193, p = 0.0005).
Nest-Tree Foragers’ Behaviour in Daylight and Dim Conditions
The nest-tree foragers, with the nest tree to the South of their nest, were displaced 5m, 10m and 20m from the nest in various directions. In initial headings, ants tested at 5m displacement sites were oriented towards the direction of the nest (Watson test, E:p<0.05,W:p<0.05, N:p<0.05, S:p<0.05), while at 10m, only ants tested at the North and West sites were significantly oriented towards the direction of the nest (Figure 7). In addition, the nest-tree foragers’ mean vector did not show orientation towards the nest at any of the 20m displacement sites (V-tests). Rayleigh tests showed non- random distributions at all 5m displacement sites, but only selected 10m displacement sites (Table 1).
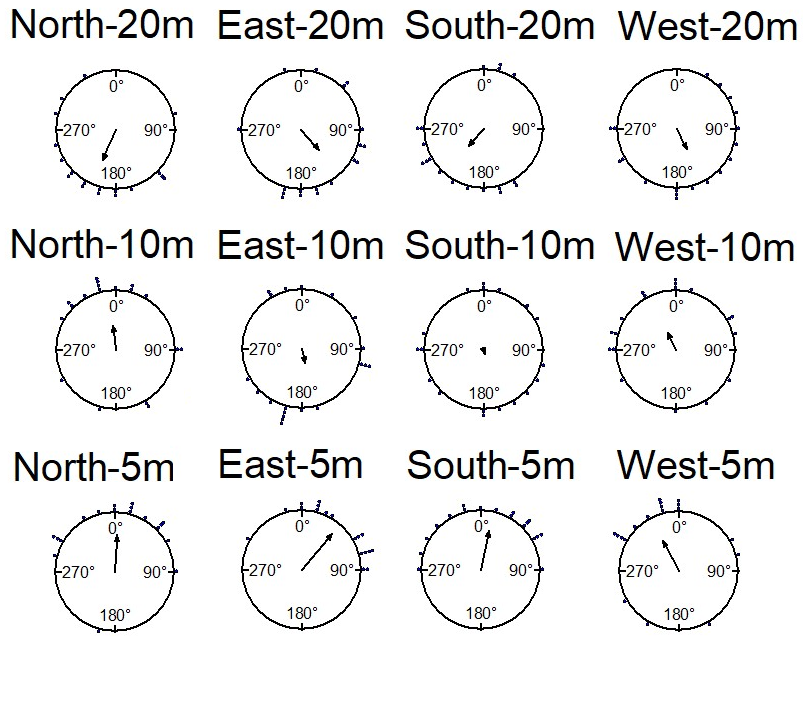
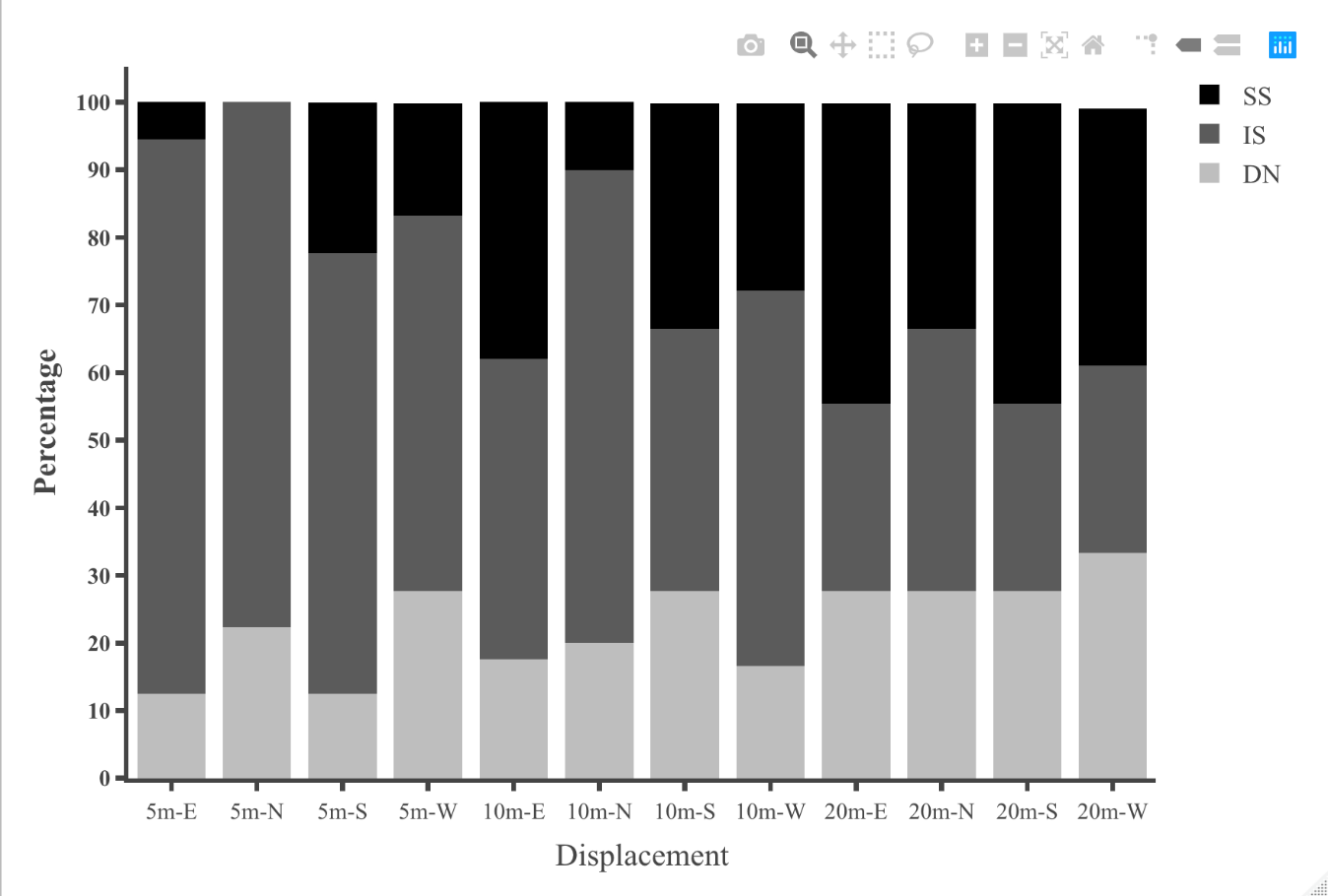
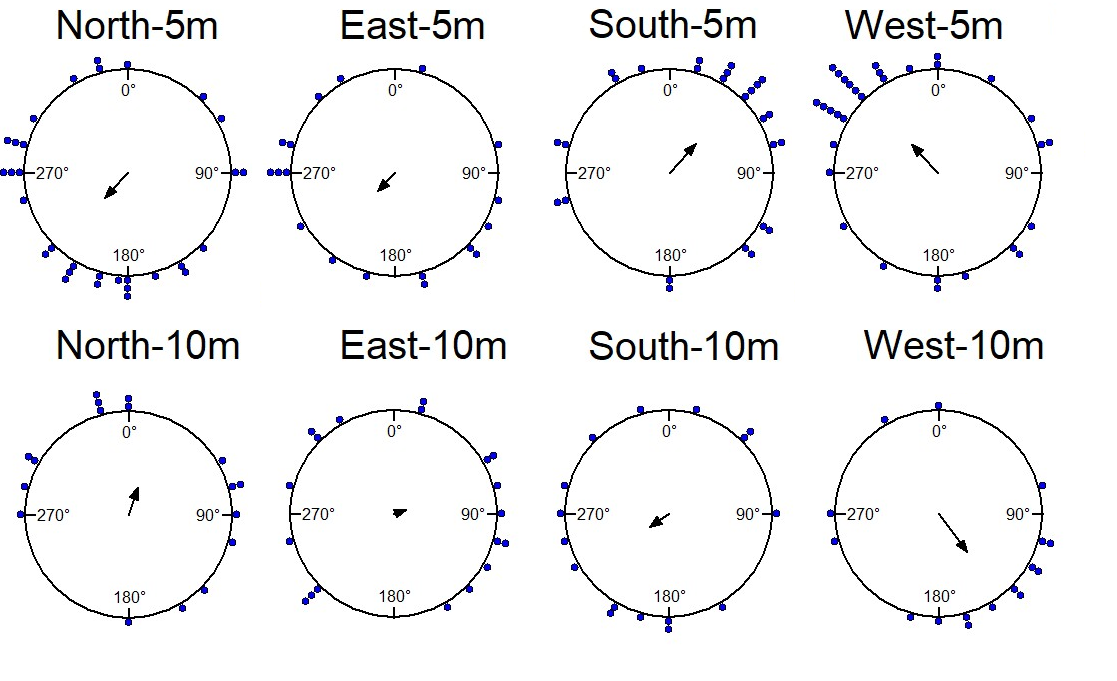
With regard to navigational behaviours at the start of displacement tests in the daylight condition, at 5m distance the majority of the nest-tree foragers showed Immediate Scanning (Figure 8). As the distance increased nest-tree foragers also switched more to Systematic Searching or Direct Navigation. In addition, the proportion of ants that returned home successfully within the 20-minute duration decreased with increasing displacement distances (Figure 9). In the daylight condition, the ants appeared to walk faster and straighter and chose a heading direction quicker at the shorter distances, with the paths becoming wider at longer distances. The path straightness of nest-tree foragers at different displacement sites was measured. The nest-tree foragers which performed direct navigation displayed a straighter path (Figure 12B) compared to ants that performed Immediate Scanning and Systematic Search. There was a significant interaction between the distance and the initial navigational behaviour of the nest-tree foragers (F = 96.38, df = 8, 209, p = 0.005). With increase in distance ants showed less path straightness and switched the initial home finding strategy to systematic search. The two-way ANOVA also showed statistically significant differences in path straightness with distance (F= 4.5, df = 4, 213, p = 0.01) and across initial navigational behaviours (F =367.14, df = 4, 213, p = 0.0005). The nest-tree foragers were collected at evening twilight and tested the following night at 5m and 10m displacement sites (Figure 10). Only ants from two 5m displacements tests oriented significantly towards the nest. None of the tests at 10 m showed significant nest ward initial orientation (Table 1).
In addition, on the goniometer, we recorded the ants’ initial navigational behaviours within the 45cm radius. In dim conditions ants exhibited more Systematic Searching and Direct Navigation and less Immediate Scanning, compared with daylight conditions (Figures 11 & 12). Ants appeared to take more time to depart from the displacement site in the dim condition. In addition, ants were numerically less successful in reaching home compared with ants in daylight conditions (Figure 9). Inferential statistics of nest-tree daylight vs dim conditions, however, showed no significant difference in success rate with Fisher’s exact test (p = 0.7). However, the proportions of initial navigational behaviours were significantly different in the dim condition compared to the daylight condition (Fisher’s exact test, p = 0.04).
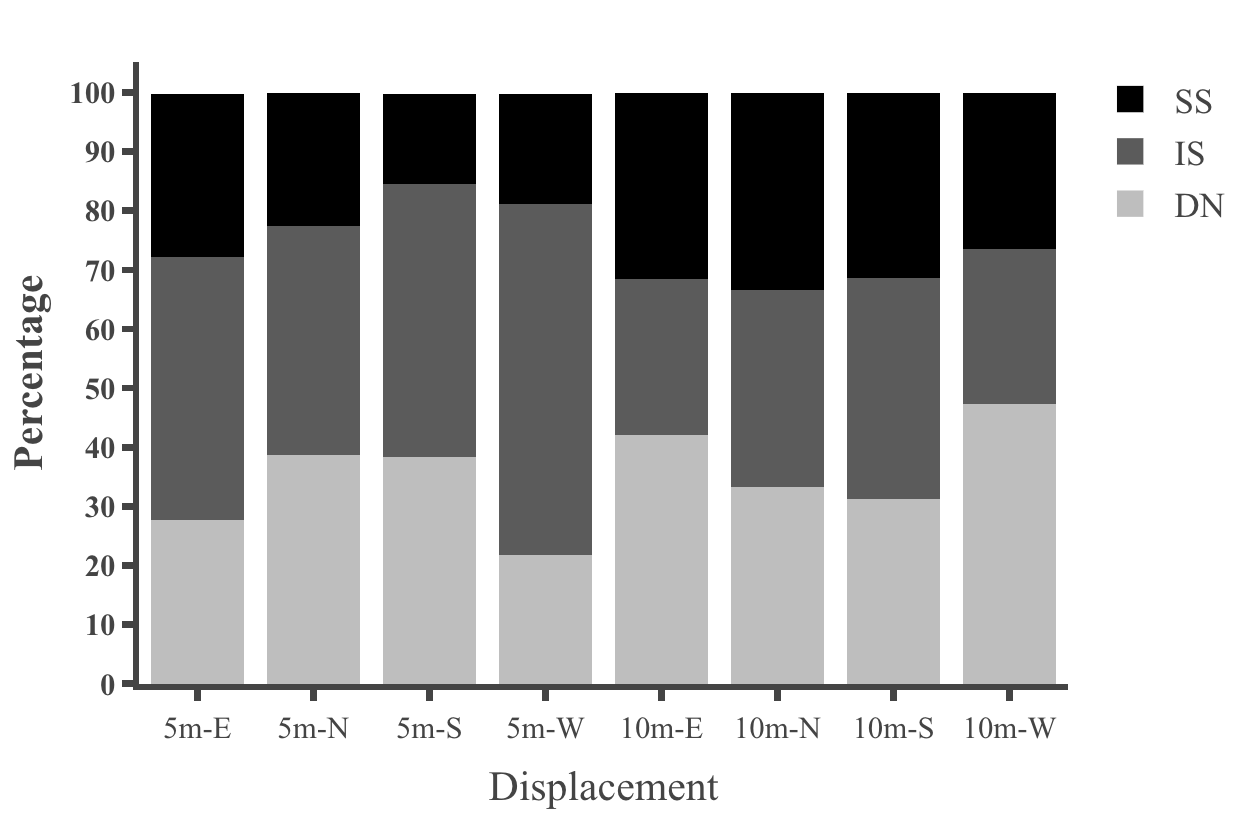
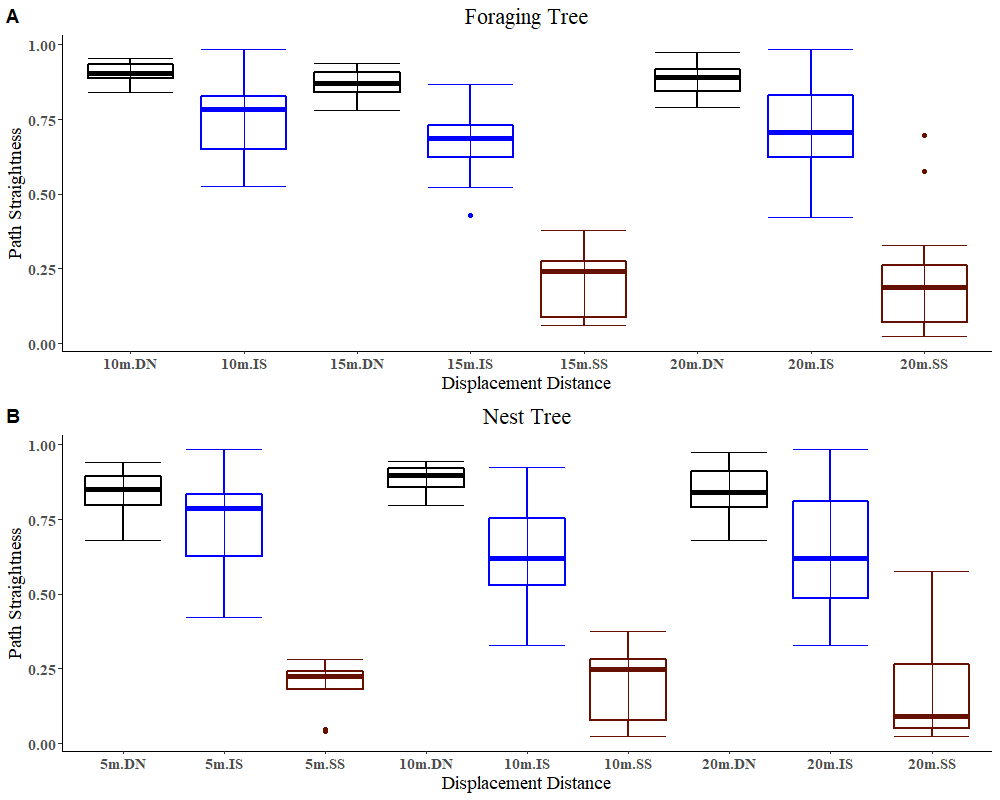
Figure 12: The path straightness of ants showing different initial navigational behaviours on tests in daylight. The x-axis shows different initial behaviours, Direct Navigation (DN), Immediate Scanning (IS), and Systematic Searching (SS), in the displacement tests. The box plots show the median (middle line in the box), the lower and upper quartiles of the box, and 1.5 times the interquartile range (whiskers).
Discussion
The current study examined the initial orientation, path straightness, and the initial navigational behaviours of M. midas foragers at displacement sites at different distances from the ants’ nest under daylight and dim (night) conditions. In the daylight condition, foraging-tree foragers were oriented towards the nest direction even from some 15m displacement sites, but not from any 20m displacement site. In dim conditions, these foragers fared much worse, with initial orientations directed towards the nest from only one 10m displacement site. In daylight, nest-tree foragers exhibited nest-oriented initial directions at 5m while at night, these ants were only oriented at 2 of 45m displacement sites. Additionally, at shorter distances, both nest-tree and foraging-tree foragers displayed more Immediate Scanning (scanning near the starting point) in their initial navigational behaviour, while with increasing distance they switched more to Systematic Searching (looping behaviour) or Direct Navigation (heading off in some direction without scans). Success at finding the nest decreased with increasing displacement distance and with decreasing light (night vs. day). While these ants do not go foraging in the day time in their normal lives, our results suggest that their orientation and navigation improve in daylight conditions compared with evening twilight and night conditions.
Ants are known to compare stored views with current views to identify their familiar heading direction to pinpoint their goal and generalise their views to some distant sites that they have not visited [32, 33, 34, 35, 36]. In evening twilight conditions, the normal foraging time of M. midas, ants that foraged at a foraging tree could home successfully from 10.5m away in roughly the same sector as their foraging tree when viewed from the nest [35]. Most of those ants homed successfully. Here, the results show that in daylight conditions, a majority of M. midas foragers could home successfully even from 15m, although the success rate dropped at 20m displacement distance. In night conditions, which have less light than during evening twilight [36, 37], the success rate was not high even at 10m displacement distance. Increased light levels led to better generalisation of views, even if the views were learned in twilight conditions.
In evening twilight conditions, the normal foraging time of M. midas, nest-tree foragers show high navigational success up to 4.5m away from the nest [35]. Here, the results show that in daylight conditions, a majority of M. midas foragers could home successfully even from one of the 10m displacement sites, although the success rate dropped at 20m displacement distance. In night conditions, which have less light than during evening twilight [37], the success rate was not high even on some of the 5m displacement tests. Again, increased light levels improve navigation while decreased light levels hinder navigation.
The initial navigational behaviour changed as displacement distance increased. The ants switched their navigational behaviour from Immediate Scanning to more Systematic Searching or Direct Navigation. Desert ants engage in systematic search when view familiarity is very low and they do not have a path-integrated vector to run off [25]. It makes sense that this behaviour increases with larger displacement distances because the scenes would be less familiar. The increase in Direct Navigation with increasing displacement distance demands a different explanation. In our observations, at shorter distances ants headed in the nest direction whereas at longer distances they headed off in random directions. The straight-line navigation or Direct Navigation is reminiscent of attempts to move away quickly from a starting point, well studied in ball-rolling dung beetles, which maximize the efficiency of an escape in any random direction after they have obtained their valuable ball of dung [43, 44, 45]. Were ants in very unfamiliar terrain perhaps heading off quickly in the hopes of finding terrain with some familiarity? Why some ants exhibited Direct Navigation at distant test sites is a question requiring more investigation.
Conclusion
In conclusion, this study found that M. midas foragers exhibit differences at initial orientation, path straightness, and initial navigational behaviours at displacement sites under daylight and dim (night) conditions. At night, they had more problems in finding their nest, perhaps explaining why these ants mostly remain in a tree throughout the night, even at a site with much light pollution.
Acknowledgements
We thank the Macquarie University for giving us access and support in the field site on campus.
Funding
This research was supported by a grant from the Australian Research Council (DP 1598700) and by Macquarie University. The work was also supported by the Australian Research Council [DP200102337] and by the Australian Defence [AUSMURIB000001 associated with ONR MURI grant N00014-19-1-2571].
Author Contributions
Experiments conceived and designed by Sudhakar Deeti and Ken Cheng. Sudhakar Deeti, Vito Lionetti and Muzahid Islam carried out experimentation and collected data. Sudhakar Deeti analysed the data. Sudhakar Deeti, Trevor
Murray and Ken Cheng drafted and revised the manuscript.
Ethics Standards
Australia has no ethical regulations regarding work with insects. The study was non-invasive and no long-term aversive effects were found on the nests or on the individuals studied.
Competing Interests
The authors declare no competing or financial interests.
References
-
Wehner R (2008) The Desert Ant’s Navigational Toolkit: Procedural Rather than Positional Knowledge. Navigation 55(2) 101-114.
-
Cheng K, Schultheiss P, Schwarz S, Wystrach A, Wehner R (2014) Beginnings of a synthetic approach to desert ant navigation. Behavioural Processes 102: 51-61.
-
Wehner R (2020) Desert navigator: the journey of an ant. Harvard University Press.
-
Wittlinger M, Wehner R, Wolf H (2007) The desert ant odometer: a stride integrator that accounts for stride length and walking speed. Journal of Experimental Biology 210: 198-207.
-
Mittelstaedt M, Mittelstaedt (1980) Homing by path integration in a mammal. Naturwissenschaften 67(11): 566-567.
-
Wehner R, Srinivasan MV (1981) Searching behaviour of desert ants, genus Cataglyphis (Formicidae, Hymenoptera). Journal of Comparative Physiology A 142: 315-338.
-
Schmidt I, Collett T, Dillier F, Wehner R (1992) How desert ants cope with enforced detours on their way home. Journal of Comparative Physiology A 171(3): 285- 288.
-
Wittlinger M, Wehner R, Wolf H (2007) The desert ant odometer: a stride integrator that accounts for stride length and walking speed. Journal of Experimental Biology 210: 198-207.
-
Collett TS (1992) Landmark Learning and Guidance in Insects. Philosophical Transactions: of the Royal Society Biological Sciences 337(1281): 295-303.
-
Graham P, Cheng K (2009) Which portion of the natural panorama is used for view-based navigation in the Australian desert ant?. Journal of Comparative Physiology A 195(7): 681-689.
-
Collett M (2010) How desert ants use a visual landmark for guidance along a habitual route. Proceedings of the National Academy of Sciences 107(25): 11638.
-
Schwarz S, Cheng K (2010) Visual associative learning in two desert ant species. Behavioural Ecology and Sociobiology 64(12): 2033-2041.
-
Steck K, Hansson BS, Knaden M (2009) Smells like home: Desert ants, Cataglyphis fortis, use olfactory landmarks to pinpoint the nest. Frontiers in Zoology 6(1): 1-8.
-
Seidl T, Wehner R (2006) Visual and tactile learning of ground structures in desert ants. Journal of Experimental Biology 209(17): 3336-3344.
-
Buehlmann C, Hansson BS, Knaden M (2012) Desert ants learn vibration and magnetic landmarks. Plos One 7(3): e33117.
-
Kleineidam CJ, Ruchty M, Casero-Montes ZA, Roces F (2007) Thermal radiation as a learned orientation cue in leaf-cutting ants (Atta vollenweideri). Journal of Insect Physiology 53(5): 478-487.
-
Müller M, Wehner R (2010) Path integration provides a scaffold for landmark learning in desert ants. Current Biology 20(15): 1368-1371.
-
Fleischmann PN, Christian M, Müller VL, Rössler W, Wehner R (2016) Ontogeny of learning walks and the acquisition of landmark information in desert ants, Cataglyphis fortis. Journal of Experimental Biology 219(19): 3137.
-
Jayatilaka P, Murray T, Narendra A, Zeil J (2018) The choreography of learning walks in the Australian jack jumper ant Myrmecia croslandi. Journal of Experimental Biology 221(20): jeb185306.
-
Deeti S, Cheng K (2021) Learning walks in an Australian desert ant, Melophorus bagoti. Journal of Experimental Biology 224(16): 242177.
-
Fleischmann PN, Grob R, Wehner R, Rössler W (2017) Species-specific differences in the fine structure of learning walk elements in Cataglyphis ants. Journal of experimental Biology 220(13): 2426-2435.
-
Deeti S, Cheng K, Graham P, Wystrach A (2023a) Scanning behaviour in ants: an interplay between random- rate processes and oscillators. Journal Comparative Physiology A 209: 625-639.
-
Schultheiss P, Cheng K (2011) Finding the nest: inbound searching behaviour in the Australian desert ant, Melophorus bagoti. Animal Behaviour 81(5): 1031-1038.
-
Schultheiss P, Wystrach A, Legge ELG, Cheng K (2013) Information content of visual scenes influences systematic search of desert ants. Journal of Experimental Biology 216(4): 742-749.
-
Schultheiss P, Cheng K, Reynolds AM (2015) Searching behaviour in social Hymenoptera. Learning and Motivation 50: 59-67.
-
Zeil J, Hofmann MI, Chahl JS (2003) Catchment areas of panoramic snapshots in outdoor scenes. Journal of the Optical Society of America A 20(3): 450-469.
-
Zeil J (2012) Visual homing: an insect perspective. Current Opinion in Neurobiology 22(2): 285-293.
-
Narendra A, Gourmaud S, Zeil J (2013) Mapping the navigational knowledge of individually foraging ants, Myrmecia croslandi. Proceedings of the Royal Society B: Biological Sciences 280(1765): 20130683.
-
Stürzl W, Zeil J, Boeddeker N, Hemmi JM (2016) How wasps acquire and use views for homing. Current Biology 26(4): 470-482.
-
Stürzl W, Zeil J (2007) Depth, contrast and view-based homing in outdoor scenes. Biological Cybernetics 96(5): 519-531.
-
Collett TS, Lent DD, Graham P (2014) Scene perception and the visual control of travel direction in navigating wood ants. Philosophical Transactions of the Royal Society B: Biological Sciences 369(1636): 20130035.
-
Fleischmann PN, Rössler W, Wehner R (2018) Early foraging life: spatial and temporal aspects of landmark learning in the ant Cataglyphis noda. Journal of Comparative Physiology A 204(6): 579-592.
-
Wystrach A, Beugnon G, Cheng K (2012) Ants might use different view-matching strategies on and off the route. Journal of Experimental Biology 215(1): 44.
-
Deeti S, Fujii K, Cheng K (2020) The effect of spatially restricted experience on extrapolating learned views in desert ants, Melophorus bagoti. Animal Cognition 23: 1063-1070.
-
Freas C, Cheng K (2019) Panorama similarity and navigational knowledge in the nocturnal bull ant Myrmecia midas. Journal of Experimental Biology 222(11).
-
Narendra A, Reid S, Raderschall C (2013) Navigational efficiency of nocturnal Myrmecia ants suffers at low light levels. Plos One 8(3) e58801.
-
Narendra A, Kamhi JF, Ogawa Y (2017a) Moving in dim light: behavioral and visual adaptations in nocturnal ants. Integrative and Comparative Biology 57(5): 1104- 1116.
-
Narendra A, Ramirez-Esquivel F (2017b) Subtle changes in the landmark panorama disrupt visual navigation in a nocturnal bull ant. Philosophical Transactions of the Royal Society B: Biological Sciences 372(1717): 20160068.
-
Islam M, Deeti S, Kamhi JF, Cheng K (2021) Minding the gap: learning and visual scanning behaviour in nocturnal bull ants. Journal of Experimental Biology 224(14).
-
Islam M, Deeti S, Mahmudah Z, Kamhi JF, Cheng K (2023) Detouring while foraging up a tree: What bull ants (Myrmecia midas) learn and their reactions to novel sensory cues. Journal of Comparative Psychology 137(1): 4-15.
-
Deeti S, Islam M, Freas C, Murray T, Cheng K (2023b) Intricacies of running a route without success in night-active bull ants (Myrmecia midas). Journal of Experimental Psychology: Animal Learning and Cognition 49(2): 111.
-
Lionetti VAG, Deeti S, Murray T, Cheng K (2023) Resolving conflict between aversive and appetitive learning of views: how ants shift to a new route during navigation. Learnng and Behaviour 51: 446-457.
-
Dacke M, Nilsson DE, Scholtz CH, Byrne M, Warrant EJ (2003) Animal behaviour: insect orientation to polarized moonlight. Nature 424(6944): 33.
-
Dacke M, Nordstrom P, Scholtz CH (2003) Twilight orientation to polarised light in the crepuscular dung beetle Scarabaeus zambesianus. Journal of Experimental Biology 206(9): 1535-1543.
-
Smolka J, Baird E, Jundi BEl, Reber T, Byrne M (2016) Night sky orientation with diurnal and nocturnal eyes: dim-light adaptations are critical when the moon is out of sight. Animal Behaviour 111: 127-146.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework