Interactive Effect of Honey Bees [Apis mellifera, Apis cerana (Hymenoptera: Apidae)] and Native Bees for Pollination Services in Radish, Raphanus sativus
Radish flowers, being self-incompatible are highly reliant on insect visitors for pollination. Honey bees are effective pollinators of radish. Pollination is influenced by visitation rates, behaviour and pollination effectiveness of pollinators which aid in seedset. We explored bee behaviour known to favour pollination; flowers visited per unit time and time required to process a flower by honey bees and non-Apis bees visiting radish. During our study, 54 species/morphospecies were observed to visit radish flowers, of which Apis cerana indica is predominant. Pollinator effectiveness was measured by means of seed set by flowers receiving a single visit of a specific pollinator as compared to seed set by flowers allowed for open pollination (multiple visits) and was found to be 0.31 and 0.36 for A. cerana and Apis mellifera, respectively. Yield enhancement studies through entomophily showed that, flowers receiving unrestricted visit of pollinators (open control) recorded 265.5% higher yield followed by interaction of two pollinators (A. c. indica + A. mellifera) 198.3%, A. mellifera (163.8%) and A. c. indica (178.3%) with respect to radish flowers allowed only for self-pollination. Pollinator diversity enhanced pollination and seed set in radish. These advantages of pollinator synergies and benefits of biodiversity are increasingly recognized.
Introduction
There are numerous examples of positive relationship between biodiversity and ecosystem services like pollination [1, 2]. Pollination is an essential ecosystem service that can improve the quality and quantity of fruits as well as seeds of 39 of the world’s 57 main crops [3]. Percent fruit set is enhanced when a diverse pollinator community exists within an ecosystem [4, 5]. Few studies show that the native bees are more efficient than honey bees in pollination when the visitation rate is considered or can complement the dominant pollinators [6, 7, 8, 9]. A recent meta-analysis by Foldesi, et al. [10] stated that the amount of pollen deposited on a stigma by a flower visitor varies greatly, as it depends upon the morphological traits of the flower visitor. Within diverse pollinator communities, interspecific interactions [11], or resource competition [12] modifies the pollination behaviour in such a way to increase pollination efficiency. Such alterations in pollinator behaviour are essential in crops bearing distinct male and female flowers or crops with self-incompatibility (e.g., Radish, Raphanus sativus L.). Though most studies find that pollinator diversity is largely considered for effective plant pollination, some exceptions are noted [13, 14], suggesting that it is important to study crop-pollinator interactions on a case-by-case basis.
The flowers of radish are sporophytically self- incompatible and considered allogamous [15, 16]. For effective pollination and successful fertilization, radish depends upon synchronous flowering of male and female flowers and efficient pollinator visits [17]. Insects, particularly of the order Hymenoptera (Apis sp., Andrena savignyi, Ceratina sp. Bombus sp. Megachile sp. and Halictus sp.), some dipterans (e.g., Syrphidae) as well as lepidopterans are regarded as the main pollinators of radish [7]. Both honey bees i.e., Apis cerana (Asian honey bee) [18] and Apis mellifera (European honey bee) [19, 20] are primary pollinating agents in radish.
Apart from honey bees, the pollinator community visiting radish has been recorded as highly diverse [17]. Yield deficits due to inadequate pollination have been demonstrated in a number of crops worldwide and concern about overreliance on a small number of managed species underscores the importance of verifying the pollination performance of flower-visiting insects to provide guidelines regarding pollination management [21]. Many researchers aim to correlate pollinator effectiveness with crop yield. With increased diversity of bee pollinators in radish, there was a significant increase in the production of seeds in radish [17]. Although there are many different ways to measure the efficiency, the method proposed by Spears [22] is particularly useful in that it considers not only seed production following a single visit but also how single visit seed production compares to the seed set of unvisited flowers and flowers receiving unrestricted pollinator visits. With this methodology, the effectiveness of particular flower visitors and frequency of visits required for optimum seed set can be described. Davis [23] discovered the enormous value of assessing single visits to virgin flowers of Echium plantagineum in order to screen flower-visiting insects as pollinators, and to rank pollinator effectiveness. Fruit or seed set from the flowers receiving a single visit by a particular species reveals its innate capacity for pollen vectoring efficiency [24]. This method can also be utilized to find the efficiency of various pollinators [25].
In radish, we assessed the diversity, abundance, and foraging behavior of flower visitors to radish. We evaluated single visit efficiency of A. c. indica, A. mellifera, Andrena savignyi, C. similima along with the pollination services of Apis supplemented with non-Apis native wild bees.
Materials and Methods
Study Area
Research was conducted at ICAR-Vivekananda Parvatiya Krishi Anusandhan Sansthan; Almora at the. Experimental farm, Hawalbagh (altitude 1250 amsl), Uttarakhand, India (29º38´01˝ N and 79º37´49˝ E). Seeds of radish, Raphanus sativus var. Dunagiri were sown in the field during September of each year, 2017-2019, following recommended agronomic practices [26]. Root cutting were obtained by November and transplanted into the main field (~0.5 ha area) with 50x50 cm spacing and allowed to bloom during March-April the following year.
Quantification of Diversity and Abundance of Pollinators
In-Situ Counts: To assess the diversity of flower visiting insects in radish during the flowering period, the fields were monitored regularly from flower initiation up to crop maturity stage by walking in a zigzag manner in the field. All flower visiting insects were recorded by scan sampling method [27] and one respective specimen of each flower visitor was collected through hand net, it was identified and preserved as voucher specimen in the repository of ICAR- VPKAS, Almora. Flower visitors belonging to hymenopterans were morphologically characterized based on pictorial, interactive and dichotomous keys [28, 29, 30]. Dipterans were identified through taxonomic keys (pictorial, linear and dichotomous) prepared by Buck M, et al. [31] and Marshall SA [32]. Furthermore, Lepidopterans were identified with the help of technical bulletin on butterflies of Almora designed by Stanley, et al. [33]. Coleopterans were characterized with the help of specimens of white grub beetle (Himalayan white grub species) preserved in compendium at ICAR-VPKAS, Almora, Uttarakhand, India.
Pollinator abundance was assessed three times a day (i.e. 10 AM, 1 PM and 4 PM) across different flower densities (i.e., low=20%, medium=50%, high=100%, medium post- peak=48% and low post-peak=25%) each for four sunny days. These flower densities were selected according to the number of flowers present/m2 after the initiation of flowering in radish. Different 1 m2 quadrants were marked on the field and the flowers were counted in 10 places to arrive at a range which implies low, medium and high flower densities. Further, two more situations after peak flowering were taken as medium post-peak and low post-peak (end of the flowering season). For estimating the abundance, four 1 m2 quadrants were marked in the field and the insects visiting the radish flowers in one-minute time were counted Stanley J, et al. [34] under a total of 12 replications. The 1 m2 quadrants chosen were completely randomized. During the observation, the insect moving out of the marked area and returning back within a span of one minute was counted as fresh entry while the insects which visited different flowers within the same quadrant were counted only once.
Forager Recruitment for Pollen and Nectar Collection
One hundred individuals of A. c. indica and A. mellifera were counted manually in radish fields and differentiated into two categories, i.e., bees with pollen and bees without pollen at three different time frames of the day (10 AM, 1 PM and 4 PM). The data was recorded across various flower densities (i.e., low=20%, medium=50%, high=100%, medium post- peak=48% and low post-peak=25%) each for four bright sunny days. The plot chosen to study the individuals of A. c. indica and A. mellifera were completely randomized and the data collected was transformed into percentage. The aim of recording bees without pollen indicates that, those bees are seeking for nectar only.
Quantification of Pollination Behaviour
Pollination behaviour of honey bees (A. c. indica and A. mellifera) were studied across various flower densities at different hours of the day each for four bright sunny days. The following bee foraging behaviour parameters were recorded; peak period of bee visitation each day, number of flowers per minute, and time spent (in seconds) per flower by pollinators with and without pollen. A total of 10 observations on the flower visitation and flowers processed per minute were made at three different time intervals (10 AM, 1 PM and 4 PM) of the day across different flower densities and mean values were calculated [34, 35]. Similarly pollination behaviour was also calculated for non-Apis bees. Further, flower searching time was also calculated from the data on the time spent per flower and flowers visited/min (60 sec) [34].
60 seconds Processing time Time spent for searching the flower= Flowers visited per minute − ∗ Note*: Processing time = Flowers visited per minute x time spent per flower.
Quantification of Pollination Efficiency
Pollination efficiency of A. c. indica and A. mellifera in radish was studied by imposing six treatments viz., T1- A. cerana single visit, T2- A. mellifera single visit, T3-Andrena savignyi single visit, T4-Ceratina similima single visit, T5- no bee visit, and (T6) multiple or unrestricted pollinator visits. About 100 flower buds were randomly selected, tagged with red threads for the first treatment and closed one day before anthesis to prevent undesirable pollinator visits using small cloth covers. The size of the covers was small enough to accommodate petiole of single flower and large enough to provide optimum space for the flowers to open within it. Similarly 100 flower buds (2nd, 3rd, 4th, 5th, and 6th treatment) were tagged with green, orange, blue, white and yellow threads for single bee visit and closed one day before flowering. On the day of anthesis of red tagged flowers, one pollen foraging A. c. indica was captured from same radish field in a transparent glass tube and allowed to visit/pollinate the flower once as per Stanley, et al. [34, 35]. The flower was carefully covered immediately to prevent other pollinator visits. The same procedures was followed for green, orange, blue tagged flowers but were pollinated with forager of A. mellifera, Andrena savignyi and C. similima forager respectively. Another set of 100 flower buds under the 5th treatment were tagged with white threads covered to exclude pollinator visit while for 6th treatment flower buds were tagged one day before anthesis with yellow threads and allowed for multiple or unrestricted visits of pollinators. On the fourth day of pollination, the covers were removed and the pods were allowed to grow till maturity with the coloured thread tags still tied on to the petiole. The pods with tags were monitored at regular intervals to have a track on the pod development and the tagged flowers which did not produce pods were noted. At the time of full maturity, the pods tagged with similar coloured threads were collected separately, brought to laboratory and counted for the seeds. The average number of seeds per pod was calculated for each treatment. Pollinator effectiveness was calculated separately for A. c. indica, A. mellifera, Andrena savignyi, and C. Similima based on the number of seeds per treatment using the Spears formula [22]: Pollinator effectiveness (PE) = (SB – NB)/ (MB– NB) Where, PE – pollinator effectiveness SB – mean number of seeds set per flower received single bee visit NB – mean number of seeds per flower restricted for pollinator visits. MB – mean number of seeds per flower with unrestricted (multiple) pollinator visits. No. of bee visits required for optimum seed set = 1/ PE.
Interaction of Apis and Non-Apis Bee Pollinators in Pollinating Radish
To study the interaction of honey bees with non-Apis bees in pollinating radish, a total of twelve treatments with three replications were imposed (T1- A. c. indica alone, T2- A. mellifera alone, T3-Andrena savignyi alone, T4-Ceratina similima alone, T5- A. c. indica + A. mellifera, T6- A. c. indica + Andrena savignyi, T7- A. c. indica + Ceratina similima, T8- A. mellifera + Andrena savignyi, T9- A. mellifera + Ceratina similima, T10- Andrena savignyi + Ceratina similima, T11- control open and T12- control close). The treatments were assigned in a completely randomized block design (CRBD). The interaction was calculated based on number of seeds per square metre and the variation in seed weight among different treatments. With the onset of flower bud in radish, cages of 1m3 were set up in the radish field and covered with mosquito net (mesh size was 1.2×1.2mm2 which was small enough to prevent insect access). Next day, 10 bees of particular treatment were released for pollination in each covered plot. The Apis and non-Apis bees were collected from same radish field and put into the cages. At harvest, the respective pods among different treatment (T1-T12) were collected and seed obtained from each treatment were weighed seperately. Additionally 100 pods from each treatment were randomly selected and average seeds/ siliqua, seed weight/10 silique, 100 seed weight (test weight) and length of siliqua were analyzed by utilizing ANOVA and average separated by LSD.
Yield in treated plots yield in closed control plots Interaction among different pollinators 100 Yield in closed control plots − = ×
Data Analysis
All the field experiments were set up in a completely randomized block design (CRBD) with twelve treatments and four replications each. The quantitative yield data was analyzed by calculating the average values through Microsoft Office Excel 2019 (Microsoft corp., USA) and the ANOVA was assessed at p < 0.05 level of significance. Further, LSD test was conducted through SPSS software for WINDOWS version 16.0 (SPSS Inc, Chicago) for comparison of statistically significant yield data in different treatments. A generalized linear mixed-effect model (GLMM) was applied to the data of pollination behaviour using R [36]. Package ‘lme4′ [37] was used for the GLMM.
Results
Diversity and Abundance of Flower Visitors Of Radish
A total of 54 species/morphospecies of flower visitors Table 1 were recorded in the radish crop, which were collected and preserved as voucher specimens. The study revealed that visitors to radish flowers were generally represented by three insect orders: Hymenoptera (47.77%), Diptera (37.08%) and Lepidoptera (15.05) (Table 2). This pollinator community comprised fifteen bees, five wasps, twelve flies, eighteen butterflies, one skipper and three moth species (Table 1).
| Order of Insect Pollinators | List of Insect Pollinators Vsiting Onion Flowers |
|---|---|
| Hymenoptera | Apis cerana, A. mellifera, A. florea, Bombus haemorrhoidalis, Xylocopa amethystina, X. fenestrata, X. pubescens, Ceratina similima, Ceratina sp., C. smaragdula, Andrena savignyi, Nomia sp., Megachile bicolor, M. relata, Vespula sp., Vespa velutina, Delta uniguiculata, Ischnojoppaluteator, and Megascolia azurea |
| Diptera | Episyrphus balteatus, Syrphus ribesii, Ischiodon sp., Eupeodes sp., Eristalinus sp., Eristalis arbustorum, Eristalis tenax, Chrysoma sp., Chrysomya sp., Musca domestica, Sarcophaga dux and Tabanus sp. |
| Lepidoptera | Aglais caschmirensis, Vanessa cardui, Vanessa indica, Tirumala limniace, Danaus chrysippus, Argynnis hyperbius, Lampides boeticus, Pachliopta aristolochiae, Papilio bianor, Graphium sarpedon, Graphium cloanthus, Pieris brassicae, P. canidia, Pontia daplidice, Colias erate, Colias fieldii, Gonepteryx nepalensis, Catopsilia Pomona, Celaenorrhinus leucocera, Helicoverpa armigera, Thysanoplusia orichalcea, Macroglossum sp, |
| A. c. indica outnumbered all other flower visitors on all days of observation. They represented 18.92% of all the pollinators followed by Syrphus ribesii (15.91 %), butterflies and moths combined (15.04%), non-Apis native bees 13.61%, houseflies (13.26%), A. mellifera (9.58%), and wasps (family vespidae, ichneumonidae, scoliidae) (5.04%) (Table 2). | |
| Relative abund ance | |
| F-cal. pvalue | |
| Low (30- | |
| 53)* | |
| Medium | |
| (80-119) * | |
| High (198-239) | |
| Flower density (No. of flower/m2) | |
| Mean No/ m2/min | |
| Insect species | |
| Medium | |
| Post-peak | |
| (115-129) * | |
| Low postpeak(34-57) * | |
| (%) | |
| * | |
| A. c. indica | |
| 0.64±0.17 b 1.53±0.34 ab 2.25±0.46 a | |
| 1.56±0.30 ab | |
| 1.00±0.25 b | |
| 1.39±0.19 | |
| 18.92 | |
| 3.641 | |
| 0.044 | |
| A. mellifera | |
| 0.33±0.05 | |
| 0.56±0.18 | |
| 0.97±0.31 | |
| 1.11±0.29 | |
| 0.55±0.15 | |
| 0.71±0.11 | |
| 9.58 | |
| 2.183 | |
| 0.144 | |
| A. florea | |
| 0 | |
| 0 | |
| 0 | |
| 0.17±0.17 | |
| 0.06±0.06 | |
| 0.04±0.026 | |
| 0.61 | |
| 2.137 | |
| 0.15 | |
| Xylocopa sp. | |
| 0.14±0.03 cd | |
| 0.28±0.03 bc 0.42±0.05 ab | |
| 0.44±0.06 a | |
| 0.06±0.06 d | |
| 0.27±0.04 | |
| 3.62 | |
| 13.63 0.0004 | |
| Ceratina smaragdula | |
| 0 | |
| 0 | |
| 0.06±0.06 | |
| 0 | |
| 0 | |
| 0.01±0.01 | |
| 0.15 | |
| 1 | |
| 0.451 | |
| Ceratina similima | |
| 0 | |
| 0.11±0.03 | |
| 0.14±0.06 | |
| 0.17±0.09 | |
| 0 | |
| 0.08±0.027 | |
| 1.12 | |
| 2.311 | |
| 0.128 | |
| Ceratina sp. | |
| 0 | |
| 0.03±0.03 | |
| 0.03±0.03 | |
| 0 | |
| 0 | |
| 0.01±0.007 | |
| 0.14 | |
| 0.75 | |
| 0.58 | |
| Bombus haemorrhoidalis | |
| 0.17±0.09 | |
| 0.36±0.07 | |
| 0.53±0.12 | |
| 0.44±0.19 | |
| 0.28±0.05 | |
| 0.35±0.056 | |
| 4.81 | |
| 1.417 | |
| 0.297 | |
| Andrena savignyi | |
| 0.20±0.12 | |
| 0.25±0.13 | |
| 0.53±0.21 | |
| 0.22±0.22 | |
| 0.11±0.11 | |
| 0.26±0.074 | |
| 3.55 | |
| 0.899 | |
| 0.5 | |
| Megachile sp. | |
| 0.00b | |
| 0.00b | |
| 0.08±0.08a | |
| 0.00b | |
| 0.00b | |
| 0.02±0.009 | |
| 0.22 | |
| NA | |
| NA | |
| Vespidae | |
| 0 | |
| 0 | |
| 0.11±0.03 | |
| 0.33±0.19 | |
| 0.11±0.06 | |
| 0.11±0.047 | |
| 1.51 | |
| 2.23 | |
| 0.138 | |
| Ichneumonidae | |
| 0.19±0.19 | |
| 0.28±0.27 | |
| 0.20±0.03 | |
| 0.33±0.03 | |
| 0.11±0.11 | |
| 0.22±0.073 | |
| 3.01 | |
| 0.214 | |
| 0.924 | |
| Scoliidae | |
| 0.00b | |
| 0.05±0.05ab 0.14±0.01a | |
| 0.00b | |
| 0.00b | |
| 0.04±0.018 | |
| 0.52 | |
| 4.558 | |
| 0.023 | |
| Syrphus ribesii | |
| 1.20±0.22 | |
| 1.11±0.20 | |
| 1.39±0.30 | |
| 1.11±0.33 | |
| 1.05±0.40 | |
| 1.17±0.12 | |
| 15.91 | |
| 0.189 | |
| 0.938 | |
| Eristalinus sp., Eristalis arbustorum and Eristalis tenax | |
| 0.42±0.12 | |
| 0.55±0.47 | |
| 0.67±0.09 | |
| 0.83±0.09 | |
| 0.44±0.11 | |
| 0.58±0.06 | |
| 7.91 | |
| 1663 | |
| 0.234 | |
| Houseflies | |
| 1.22±0.14 | |
| 0.80±0.16 | |
| 1.19±0.09 | |
| 0.83±0.09 | |
| 0.83±0.25 | |
| 0.98±0.08 | |
| 13.26 | |
| 2.129 | |
| 0.151 | |
| Butterflies | |
| 0.36±0.07b 0.75±0.21ab 1.11±0.14a | |
| 0.33±0.19b | |
| 0.28±0.11b | |
| 0.57±0.10 | |
| 7.69 | |
| 5.236 | |
| 0.015 | |
| Hawk moth | |
| 0.22±0.06 | |
| 0.55±0.17 | |
| 0.56±0.13 | |
| 0.17±0.16 | |
| 0.06±0.05 | |
| 0.31±0.07 | |
| 4.22 | |
| 3.275 | |
| 0.058 | |
| Helicoverpa armigera | |
| 0.07±0.07 c 0.11±0.11 b 0.88±0.28 a | |
| 0.00 d | |
| 0.11±0.11 b | |
| 0.23±0.10 | |
| 3.18 | |
| 5.894 | |
| 0.01 | |
Table 1: Diversity of Flower Visitors in Radish.
In rows, SE (Standard error) followed by different alphabetical letters (a, b or c) represent statistically significant differences for insect abundance according to flower density, with ‘a’ representing a superior group means followed with the same letter are not different statistically by LSD (P=0.05);*range of flower density; NA- not available; Low = 20% bloom, Medium = 50% bloom, High = 100% bloom, Medium (post peak) = 48% and Low (post peak) = 25%.
Forager Recruitment for Pollen and Nectar Collection
A. c. indica (Indian Honey Bee): A. c. indica reached maximum density 2.25±0.45 bees/min/m2 (P=0.010, F=10.91) at peak bloom. Foragers with pollen exceeded foragers without pollen at the end of bloom. Foragers without pollen were greater in number during peak flowering at 4 PM i.e., 59.73% (P <0.001, F=199.7) and minimum during medium post-peak flower density i.e., 32.67% (P <0.001, F=83.38) (Table 3).
| Flowers density | Density of A. c. indica (No. of bees/ m2) | Proportion of A. c. indica # | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Forager with pollen | Foragers without pollen | |||||||||||||||||
| 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | |
| Low (30-53)* | 0.67±0.14 | 0.93±0.08 | 0.33±0.14 | 0.64 | 2.481 | 0.163 | 45.3 | 47.4 | 51 | 47.9 | 8.549 | 0.057 | 54.7 | 52.6 | 49 | 52.1 | 9.443 | 0.05 |
| Medium (80- 119)* | 1.58±0.13 b | 2.08±0.16 a | 0.92±0.08 c | 1.53 | 12.31 | 0.007 | 58.1 | 65.7 | 50.1 | 57.97 | 87.56 | 0.002 | 41.90 b | 34.30c | 49.90 a | 42.03 | 29.14 | 0.01 |
| High (198- 239)* | 2.33±0.24 b | 3.00±0.14 a | 1.42±0.16 c | 2.25 | 10.91 | 0.01 | 41.22 b | 54.39 a | 40.27 c | 45.29 | 40.24 | 0.006 | 58.78 | 45.61 | 59.73 | 54.71 | 199.7 | 0.0006 |
| Medium post-peak (115-129 )* | 1.33±0.19 b | 2.17±0.17 a | 1.17±0.17 b | 1.56 | 19.19 | 0.002 | 40.99c | 67.33b | 52.99a | 53.77 | 176.9 | 0.007 | 59.01 | 32.67 | 47.01 | 46.23 | 83.38 | 0.002 |
| Low post- peak (34- 57)* | 0.83±0.17 b | 1.50±0.17 a | 0.67±0.00 b | 1 | 11.49 | 0.008 | 59.42 | 54.62 | 52.32 | 55.45 | 7.743 | 0.065 | 40.58 | 45.38 | 47.68 | 44.55 | 15.33 | 0.026 |
| Average | 1.35 b | 1.94 a | 0.90 c | 1.4 | 49.01 | 57.89 | 49.34 | 52.08 | 50.99 | 42.11 | 50.66 | 47.92 | ||||||
| F-cal. | 3.309 | 2.111 | 2.111 | |||||||||||||||
| p-value | 0.071 | 0.163 | 0.163 |
Table 2: Pollination behaviour of Indian Honey Bee, A. C. Indica in Radish, Raphanus sativus.
In column and amid particular rows, average followed by a common letter(s) are not significantly different by LSD (P=0.05); #100 bee foragers counted in the field at the particular time; * Flower density range; NA – not available; Low = 20% bloom, Medium = 50% bloom, High = 100% bloom, Medium (post peak) = 48% and Low (post peak) = 25%.
Apis mellifera (European Honey Bee): The overall density of foraging A. mellifera was highest at 1 PM followed by 10 AM with lowest density observed at 4 PM (i.e., 1.07±0.22,0.65±0.14 and 0.40±0.07 bees/min/m2 respectively, P=0.038, F=4.337). At the commencement of flowering, A. mellifera had more foragers with pollen (59.70%) (Table 4). In general, foragers with pollen outnumbered foragers without pollen at all-time point except during the medium post-peak bloom time.
| Flowers density | Density of A. mellifera (No. of bees/ m2) | Proportion of A. mellifera# | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pollen forager | Foragers without pollen | |||||||||||||||||
| 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | 10:00 AM | 1:00 PM | 4:00 PM | Mean | F-cal. | p- value | |
| Low (30- 53)* | 0.33±0.14 | 0.42±0.16 | 0.25±0.06 | 0.33 | 0.228 | 0.802 | 52.7 | 70.1 | 56.3 | 59.7 | 33.39 | 0.008 | 47.30a | 29.90c | 43.30b | 40.2 | 83.78 | 0.002 |
| Medium (80-119)* | 0.42±0.08 b | 0.92±0.08 a | 0.33±0.14 b | 0.56 | 6.011 | 0.036 | 73.1 | 64.3 | 59.92 | 65.8 | 45.59 | 0.005 | 26.9 | 35.7 | 40.08 | 34.2 | 50.58 | 0.004 |
| High (198- 239)* | 1.00±0.24 ab | 1.50±0.22 a | 0.42±0.08 b | 0.97 | 8.824 | 0.016 | 60.1 | 50.95 | 64.32 | 58.5 | 41.74 | 0.006 | 39.90b | 49.05a | 35.68c | 41.5 | 28.87 | 0.01 |
| Medium post-peak (115-129 )* | 1.00±0.19 | 1.67±0.00 | 0.67±0.00 | 1.11 | 3.886 | 0.082 | 40.38c | 51.08b | 49.02a | 46.8 | 30.05 | 0.01 | 59.62 | 48.92 | 50.98 | 53.2 | 19.1 | 0.019 |
| Low post- peak (34- 57)* | 0.50±0.17 | 0.83±0.17 | 0.33±0.00 | 0.55 | 3.274 | 0.109 | 72.52 | 63.67 | 56.72 | 64.3 | 46.76 | 0.005 | 27.48 | 36.33 | 43.28 | 35.7 | 85.35 | 0.002 |
| Average | 0.65±0.14 b | 1.07±0.2 a | 0.40±0.07 c | 0.71 | 59.76 | 60.02 | 57.26 | 59 | 40.24 | 39.98 | 42.66 | 41 | ||||||
| F-cal. | 4.337 | 0.118 | 0.111 | |||||||||||||||
| p-value | 0.038 | 0.89 | 0.896 |
Table 3: Pollination behaviour of European honey bees, A. mellifera in radish, Raphanus sativus.
In column and amid particular rows, SE followed by a common letter(s) are not significantly different by LSD (P=0.05); # Proportion of A. mellifera observed with and without pollen (of 100 bees) at each time point.; * Flower density range; NS – not available; Low = 20% bloom, Medium = 50% bloom, High = 100% bloom, Medium (post peak) = 48% and Low (post peak) = 25%.
Pollination Behaviour- Flowers Visited Per Minute
A. c. indica (Indian Honey Bee): Foragers of A. c. indica with pollen visited statically more radish flowers at 1 PM followed at 4 PM and least at 10 AM (9.7±0.17, 8.9±0.11 and 8.67±0.5 flowers/min, respectively, P=0.03, F=11.96) (Figure 1). Foragers without pollen visited most flowers at 10 AM when flower density was medium (8.99±0.12 flowers/min, P<0.001, F=123.6) with lowest visits occurring at 1 PM and 4 PM during low and low post-peak bloom (7.4±0.44 flowers/ min, P<0.001, F=56.47) (Figure 1). Apis Mellifera (European Honey Bee): Foragers with pollen visited statically more flowers during high flower density at 1 PM (9.2±0.22, P<0.005, F=109.2) flowers/min whereas they visited fewer flowers at 10 AM (i.e., 6.5±0.45, P<0.001, F=141.2) during the low flower density periods (Figure 1). Non-Apis Bees: During the observation periods, Ceratina sp. visited more flowers compared to other non-Apis bee pollinators i.e., 11.02±0.12 (P=0.006, F=112.9), whereas, A. savignyi visited the fewest flowers i.e., 6.30±0.34 flowers/min (P<0.001, F=460.0) during low flower density (Supplementary Table 1).
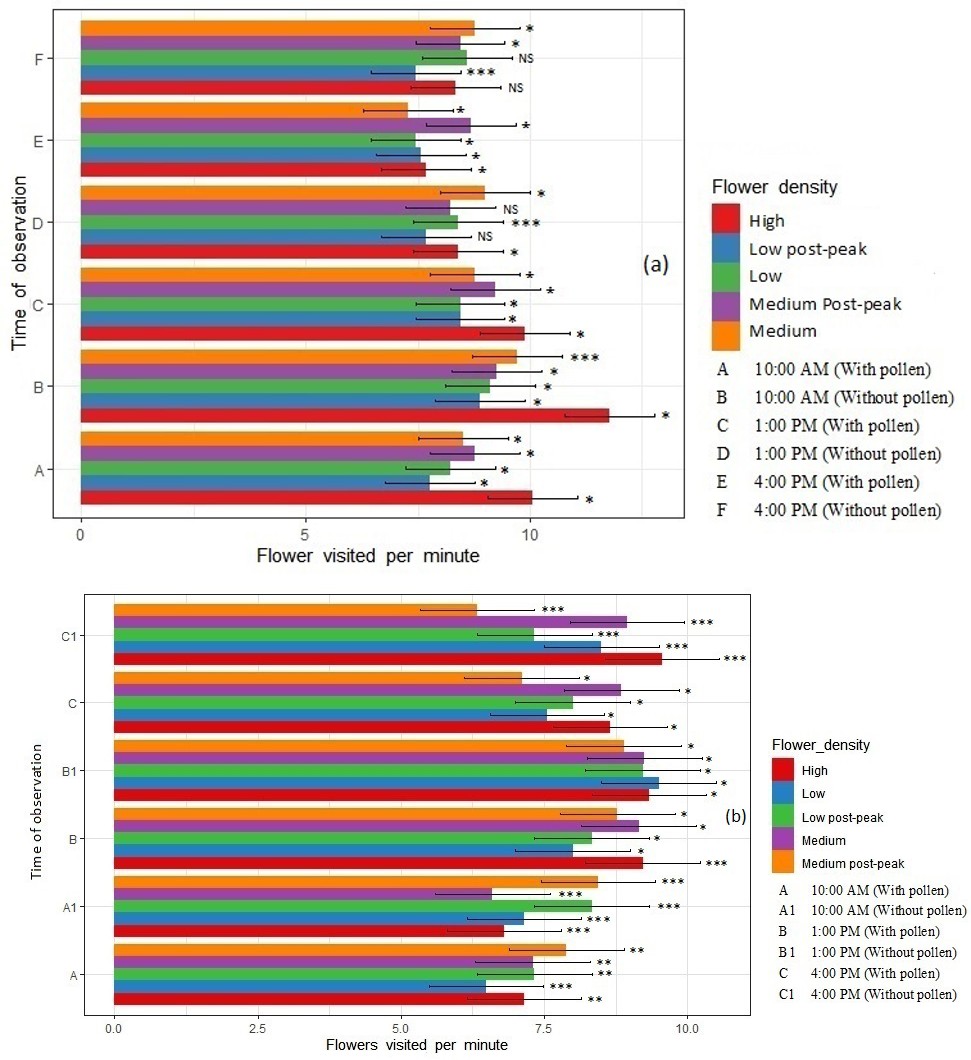
Figure 1: Mean (+/- SE) Number of Flowers Visited Per Minute and Time Spent Per Flower by A) Indian Honey Bee, Apis Cerana Indica and B) European Honey Bees, A. mellifera, in Radish (Raphanus Sativus) Plots Observed Across Various Flower Densities at Three Times during the Day (Low = 20% Bloom, Medium = 50% Bloom, High = 100% Bloom, Medium (Post Peak) = 48% and Low (Post Peak) = 25%) *, , * are Significant At P=0.05, 0.01, 0.001.
Pollination Behaviour- Time Spent Per Flower
A. c. indica (Indian Honey Bee): Foragers with pollen required more time per flower at 1 PM during low post- peak flower density (4.7±0.25 sec, P=0.004, F=28.21) than during peak flower density at 1 PM (3.1±0.56 sec, P=0.002, F=72.4). Foragers without pollen spent the statically greatest time per flower during low flower density at 1 PM (5.6±0.42 sec, P=0.003, F=69.8) and the least amount of time during medium flower density at 4 PM (3.33±0.43 sec, P=0.001, F=102.8) (Figure 1).
Apis mellifera (European Honey Bee): Foragers with pollen spent statically more time per flower during peak flower density (4.8±0.31 sec, P=0.002, F=28.23) but less time during medium flower density (4.5±0.50 sec, P=0.004, F=32.56). Foragers without pollen were observed to spend 5.1±0.41 sec (P=0.001, F=45.89), 4.4±0.22 sec (P=0.002, F=39.80) and 4.2±0.38 sec (P=0.004, F=58.1) per flower at 10 AM, 1 PM and 4 PM, respectively, across all flower densities (Figure 1). Non-Apis Bees: During the study, it was found that A. savignyi spent greatest time per radish flower (6.33±0.46 sec, P=0.001, F=128.2) at low bloom (Supplementary Table 1).
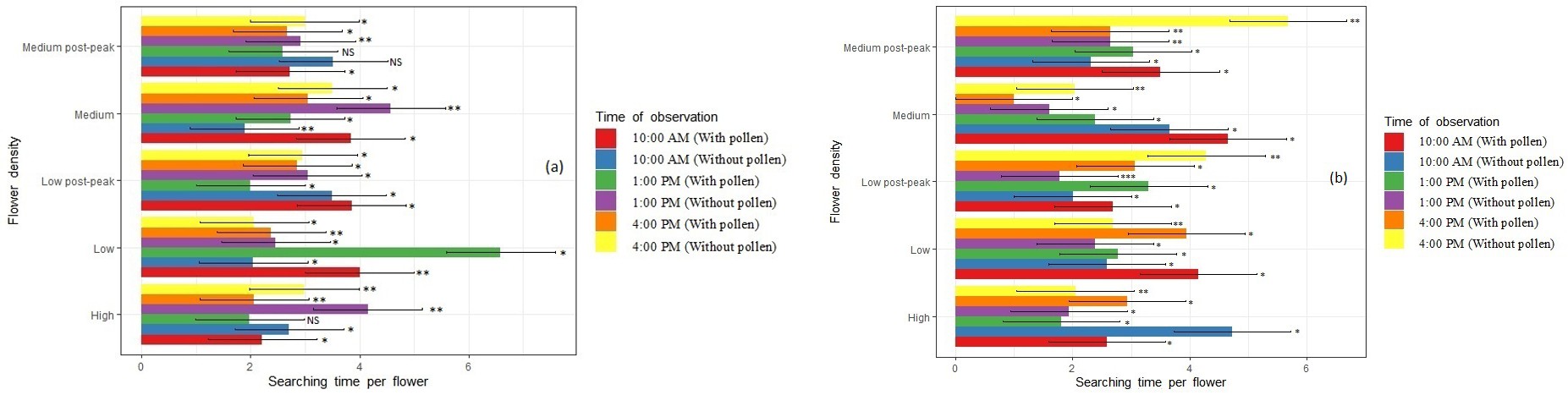
Figure 2: Time Spent for Searching Flowers by A) Apis Cerana Indica and B) A. Mellifera with and without Pollen, at the Time of Observation, while Foraging in Radish, Raphanus Sativus, throughout the Bloom. Period. Low = 20% Bloom, Medium = 50% Bloom, High = 100% Bloom, Medium (Post Peak) = 48% and Low (Post Peak) = 25%. *, , * are Significant at p=0.05, 0.01, 0.001.
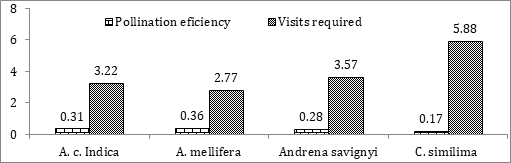
Pollination Efficiency of Bee Pollinators in Radish
The pollination efficiency of Indian honey bees on radish was calculated as 0.31 and a minimum of 3.22 bee visits are required per flower for optimum seed set. Pollination efficiency of European honey bee was calculated to be 0.36
Pollination Behaviour-Flower Searching Time
A. c. indica (Indian Honey Bee): Foragers with pollen spent more time searching for flowers at 10 AM (4.0±0.31 sec, P=0.006, F=72.5) and the least time at 4 PM (2.83±0.66 sec., P=0.005, F=54.67) when flower density was low (Figure 2).
Apis mellifera (European Honey Bee): When we compare different hours of the day foragers with pollen took statically different time to search for a radish flower at 10 AM (3.5±0.51 sec., P=0.03, F=1.59) and less time per flower at 1 PM (2.6±0.61 sec., P=0.04, F=5.69). Foragers without pollen spent more time searching for flowers at 4 PM, across all the flower densities (Figure 2).
Non-Apis Bees: During the low flower density, C. similima spent only a short time i.e., 2.0±0.62 sec (P=0.005, F=86.23). At peak flower density, A. savignyi spent statically different time to search flowers of radish 3.5±0.52 (P=0.006, F=74.3), 4.1±0.49 (P=0.03, F=16.34) and 4.2±0.37 (P=0.004, F=42.5) during 10 AM, 1 PM and 4 PM, respectively. At the end of bloom (25% bloom) Nomia sp., C. smaragdula, and Ceratina sp. were into low abundance to be in this analysis (Supplementary Figure 2).
and a minimum of 2.77 bee visits are required for optimum seed production. The pollination efficiency of A. savignyi and C. similima were calculated as 0.28 and 0.16 respectively requiring a minimum of 3.57 and 5.88 visits for optimum seed set in radish (Figure 3).
Interaction of Apis and Non-Apis Bees in Pollinating Radish
Results revealed that the total seeds obtained from flowers when bees were excluded from visiting radish flowers were just 2.26±0.62 seeds/silique and the flowers permitted for unrestricted pollinator’s visits produced 7.98±0.21 seeds per pod in radish. Flowers exposed to a single bee visit, either A. c. indica, A. mellifera, A. savignyi or C. similima produce statically different seed per pod with respect to each other (4.76±0.40, 5.00±0.61, 4.51±0.37 and
3.20±0.81 seeds per pod, P<0.001, F=156.5), respectively (Table 5). A. mellifera was the most efficient pollinator in radish in terms of number of seeds per pod while, C. similima was the least efficient.
In plots where A. c. indica and A. mellifera were visiting together, they performed statically better than either of the species in isolation, as the number of seeds per siliqua obtained were higher (i.e., 7.40±0.41, P<0.001, F=156.5) (Table 5).
| Treatment | No. of seeds/silique | Seed weight/10 silique (g) | 100 seed weight (g) | Length of Silique (cm) | Yield (g) per m² | Yield enhancement (percent) | |
|---|---|---|---|---|---|---|---|
| T1 | A. c. indica | 4.76±0.40 e | 0.525±0.05 d | 1.0561±0.56 d | 4.97±0.46 f | 43.9±5.9 | 148 |
| T2 | A. mellifera | 5.00 ±0.61d | 0.508±0.02 e | 1.1299 ±0.61c | 5.43±0.56 d | 46.7±6.7 | 163.8 |
| T3 | Andrena savignyi | 4.51±0.37 e | 0.344±0.11 i | 0.8667±0.11 i | 4.33±0.61 h | 41.8±4.9 | 136.1 |
| T4 | Ceratina similima | 3.20±0.89 f | 0.273±0.12 j | 0.9553±0.12 f | 4.07±0.42 i | 37.3±6.4 | 109 |
| T5 | A. c. indica + A. mellifera | 7.40±0.41 b | 0.607±0.03 b | 1.3094±0.30 b | 6.17±0.52 a | 52.8±5.4 | 198.3 |
| T6 | A. c. indica + Andrena savignyi | 5.56±0.55 c | 0.545±0.07 cd | 1.061±0.30 g | 5.42±0.32 d | 49.3±5.7 | 178.5 |
| T7 | A. c. indica + Ceratina similima | 4.92±0.31 e | 0.544±0.14 cd | 1.057±0.40 e | 5.59±0.71 c | 44.9±3.9 | 153.6 |
| T8 | A. mellifera + Andrena savignyi | 5.45±0.65 c | 0.562±0.08 c | 1.0030±0.12 e | 5.45±0.61 d | 51.7±5.1 | 192 |
| T9 | A. mellifera + Ceratina similima | 5.16±0.45 d | 0.521±0.10 d | 0.8850±0.41 h | 5.26±0.69 e | 48.6±6.4 | 174.5 |
| T10 | Andrena savignyi + Ceratina similima | 4.81±0.51 e | 0.357±0.10 h | 0.8534±0.07 i | 4.59±0.68 g | 43.2±6.8 | 144 |
| T11 | Control open | 7.98±0.21 a | 0.625±0.01 a | 1.3220±0.41 a | 6.07±0.39 b | 64.7±4.5 | 265.5 |
| T12 | Control close | 2.26±0.62 g | 0.211±0.20 k | 0.7299±0.31 j | 3.10±0.35 j | 17.7±6.3 | 100 |
| F-cal. | 156.5 | 14.27 | 193 | 129.5 | 289.9 | ||
| p-value | 0.0001 | 0.0005 | 0.0009 | 0.00001 | 0.001 |
Table 4: Yield Parameters in Radish Crop Pollinated by Apis and Non-Apis Bee Pollinators along with their Interaction.
In column and among particular rows, SE- followed by a common letter(s) are not significantly different by LSD (P=0.05) The data on yield enhancement show that plots receiving 10 foragers each of A. c. indica, A. mellifera, A. savignyi and C. similima recorded a yield enhancement of 148, 163 136 and 109 percent in comparison to closed control plots. The plots left open for unrestricted pollinators recorded 265.5 percent yield enhancement in comparison to plots receiving no pollinators (Control close treatment). It was recorded that plot receiving interaction of bee pollinators (5 foragers of each pollinators) recorded highest yield compared to plot receiving 10 foragers of individual bee pollinators (Table 5).
Discussion
Self-incompatibility is one of the major reasons why pollinators are the major determinant of crop yield [38, 39]. Pollinator diversity is high in radish and as many as 54 insect species were observed visiting radish flowers. Of all the flower visitors of radish, A. c. indica and A. mellifera accounted for 18.92% and 9.58%, respectively. Honey bees play an essential role in the pollination of cruciferous crops like radish [17], cabbage [34] and mustard [35]. Other non- Apis native bees (given in order of abundance) like Ceratina similima, Andrena savignyi, C. smaragdula, Ceratina sp. Megachile sp., M. bicolor, Apis florea, Bombus haemorrhoidalis, Xylocopa fenestrata were also noticed. Diverse groups of insect pollinators are helpful in ensuring the seed set by compensating and complementing with each other [40]. The syrphid population was relatively high which may also be attributed to the presence of aphids [41]. Syrphids are also considered as pollinators particularly of the Brassica crops [42, 43], although not efficient as honey bees.
Foragers of A. c. indica with pollen are more efficient pollinators than foragers without pollen in radish as they entered the flower from above, over the stigma and anthers. However, some of the foragers without pollen (~45%) were found to collect nectar from the base of flowers resulting usually in no/least pollination. This type of basal foraging for nectar collection is known among A. mellifera, A. cerana and Bombus in okra, cauliflower, radish, cabbage and mustard [34, 35, 44, 45, 46]. Increased recruitment of foragers without pollen in A. c. indica during peak flowering may not be a good sign in terms of pollination, as 45% of them are basal foragers. Pollen foragers are termed as effective pollinators in earlier studies also Davis AR, et al. [23].
Foraging behaviour is defined as the pattern by which bees collect pollen or nectar [47]. Foraging speed (i.e., time spent/flower) and foraging rate (i.e., flowers visited/min) are connected with the foraging behavior of the insects and floral structure for a particular crop, chiefly depth of the corolla as well as resource availability [48]. During the present investigation, the foraging activity was higher at 1 PM for both A. c. indica and A. mellifera. A similar result was reported in radish by Partap, et al. [18], showing the peak foraging by A. cerana between 1100 and 1400 hr. The peak foraging activity for A. mellifera, A. cerana and syrphids in Brassica campestris occurred between 1200to 1300 h [49].
It was reported that the chance of pollination increases significantly with increase in foraging frequency [50]. Foragers of A. c. indica with pollen speedily processed flowers during morning hours, irrespective of the flower densities. The foraging rate of pollen foragers of A. c. indica in radish is in line with the figures mentioned by Partap, et al. [18]. In case of A. mellifera, speed of pollination (time spent per flower) is not significantly different for different periods of observations (10 AM, I PM and 4 PM). Stanley J, et al. [34] reported similar results for A. cerana foragers in cabbage.
The basic technique to estimate the contribution of a pollinator in plant pollination is done by its visit frequency and duration of flower visitation [51, 52]. Still advanced techniques intended to estimate the transport of pollen grains [53, 54], pollen removed from the anthers [6], and its deposition on stigma [55]. All these experiments conclude in one common measure i.e., fruit set or seed yield. The technique used in the present research comprises both the plant and pollinator interaction i.e., pollinator visitation and the seed set as given by Spears EE [22]. Similar techniques with the above stated measurements are used to find out the pollinator effectiveness in Echium [23], coffee [56], Jatropha [57], cabbage [34] and mustard [35]. Apis mellifera was found to be the most efficient as measured by means of number of seed set per flower receiving a single visit. Our findings also showed that when the dominant pollinator i.e., Apis species are complemented by other bees like native wild bees the yield gets enhanced. This increased yield may be due to complementation in pollination behaviour in the presence of competitive pollinators. This study highlights the chiefly unexplored facilitative component of biological diversity along with its benefit to the ecosystem.
The interactive effect of non-Apis bees with that of honey bees is promising as the pollination efficiency gets enhanced in the presence of C. similima and A. savignyi and the honey bees, A. cerana and A. mellifera. Conserving biodiversity in agricultural ecosystems could bring unrecognized advantages, as more diverse pollination systems increase the long-term sustainable production of radish and other bee pollination dependent crops [21]. Species diversity is crucial for many ecosystem functions [58] and beyond ecosystem services [59]. No doubt honey bees are the most efficient and managed pollinators in many crops if not all. But the availability of honey bees as predicted is not proliferating at the same rate as required in the agricultural services [60]. Increasing the pollination effectiveness of honey bees and safeguarding native pollinators might assist to cause a surge in crop yields. Thus, synergistic pollination between A. cerana, A. mellifera and non-Apis native bee pollinators signifies a sustainable system to increase crop pollination, but the overview of such effects still require verification across multiple agricultural crops. Competition between honey bees and native bees are to be taken into consideration in each ecosystem. Though many reports establish the advantages of increased pollinator diversity [61], a few states the negative impact of introduced pollination on native bees particularly competition for floral resources between them [62]. In an experiment wherein pollinator density of honey bee introduced radish field and control fields were compared, we noticed a reduction in the density of A. savignyi in the honey bee introduced fields, which needs further investigation.
In conclusion, although radish flowers are visited by many insects, A. c. indica is found to be the most abundant pollinator in this location. However, A. mellifera were more efficient pollinators than Indian honey bees in pollinating
Disclosure
radish, in our study. The pollination efficiency in term of seed set per flower increases drastically when A. c. indica and A. mellifera were introduced in the cages. Similarly, when Apis species and non-Apis bee were introduced together, they showed enhanced pollination efficiency and synergistic effect in enhancing the yield. Our findings provide substantial evidence for a synergistic interaction across diverse pollinator communities. This is also in contradiction to the fear of negative interaction that might arise by adding a competing pollinator. The potential for such positive synergistic interaction should be examined for other cross- pollinated crops with a variety of pollinators.
Acknowledgements
The research was part of the SERB YSS 2015/000861 project entitled “Habitat management and conservation for non-Apis bee pollinator”. We acknowledge the humble support and supervision by the Director, and Heads, Crop Protection, ICAR-VPKAS, Almora, India. The authors are also thankful to the dept. of Zoology Kumaun University S.S.J campus, Almora for their suggestion during the course of research.
| Topic | Disclosures |
|---|---|
| Funding | The study has been funded by Indian Council of Agriculture Research, New Delhi |
| Conflict of Interest | All the authors have received the funding from ICAR, New Delhi and therefore have no conflict of interest for the submission and publication of the article to “Entomological Research”. |
| Data available within the article or its supplementary materials | The authors confirm that the data supporting the findings of this study are available within the article [and/or] its supplementary materials. |
| Code availability (software application or custom code) | The authors confirm that the software applications used in the present study are freely available in the public domain and no copyright was breached. |
Statement Ethical approval: This article does not contain any studies with human participants performed by any of the authors.
Ethical approval: This article does not contain any studies with animals performed by any of the authors.
Ethical approval: This article does not contain any studies with human participants or animals performed by any of the authors.
Author Contribution
• Sunaullah Bhat: Coordinated the work and carried out field observations.
• J. Stanley: Conceived the idea, wrote and was awarded the competitive grant of SERB, Govt of India.
• Sandeep Kumar & J.P. Gupta: Technical support and data analysis.
• A.R.N.S. Subbanna, Amit Umesh Paschapur & G
Preetha: Assisted with technical support, data analysis and manuscript preparation.
• Tasir Iqbal and Ashish Kumar Singh: Final review of the manuscript and English improvement.
References
-
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, et al. (2012) Biodiversity loss and its impact on humanity. Nature 486(7401): 59-67.
-
Hooper DU, Adair EC, Cardinale BJ, Byrnes JE, Hungate BA, et al. (2012) A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486(7401): 105-108.
-
Klein AM, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, et al. (2007) Importance of pollinators in changing landscapes for world crops. Proceedings of the Rroyal Ssociety B: Bbiological Ssciences 274(1608): 303-313.
-
Hoehn P, Tscharntke T, Tylianakis JM, Steffan-Dewenter I (2008) Functional group diversity of bee pollinators increases crop yield. Proceedings of the Royal Society B: Biological Sciences 275(1648): 2283-2291.
-
Klein AM, Brittain C, Hendrix SD, Thorp R, Williams N, et al. (2012) Wild pollination services to California almond rely on semi-natural habitat. Journal of Applied Ecology 49: 723-732.
-
Conner JK, Davis R, Rush S (1995) The effect of wild radish floral morphology on pollination efficiency by four taxa of pollinators. Oecologia 104(2): 234-245.
-
Zameer SU, Bilal M, Fazal MI, Sajjad A (2017) Foraging behavior of pollinators leads to effective pollination in radish _Raphanus sativus_ L. Asian Journal of Agriculture and Biology 5(4): 221-227.
-
Albrecht M, Schmid B, Hautier Y, Muller CB (2012) Diverse pollinator communities enhance plant reproductive success. Proceedings of the Royal Society B: Biological Sciences 279(1748): 4845-4852.
-
Brittain C, Kremen C, Klein AM (2013) Biodiversity buffers pollination from changes in environmental conditions. Global Change Biology 19(2): 540-547.
-
Foldesi R, Howlett BG, Grass I, Batáry P (2021) Larger pollinators deposit more pollen on stigmas across multiple plant species-A meta‐analysis. Journal of Applied Ecology 58(4): 699-707.
-
Greenleaf SS, Kremen C (2006) Wild bees enhance honey bees’ pollination of hybrid sunflower. Proceedings of the National Academy of Sciences 103(37): 13890-13895.
-
Inouye DW (1978) Resource partitioning in bumblebees: experimental studies of foraging behavior. Ecology 59(4): 672-678.
-
Gross CL, Mackay D (1998) Honeybees reduce fitness in the pioneer shrub Melastoma affine (Melastomataceae). Biological Conservation 86(2): 169-178.
-
Prendergast KS, Ollerton J (2021) Impacts of the introduced European honeybee on Australian bee‐ flower network properties in urban bushland remnants and residential gardens. Austral Ecology 47: 35-53.
-
Bateman AJ (1955) Self-incompatibility systems in angiosperms. III. Cruciferae. Heredity 9: 53-68.
-
Kercher S, Conner JK (1996) Patterns of genetic variability within and among populations of wild radish, Raphanus raphanistrum (Brassicaceae). American Journal of Botany 83: 1416-1421.
-
Priti RC, Mishra, Sihag RC (2001) Role of insect pollination in seed production of radish (_Raphanus sativus_ L.) Seed Research 29: 231-234.
-
Partap U, Verma LR (1994) Pollination of radish by Apis cerana. Journal of Apicultural Research 33(4): 237-241.
-
Evans LJ, Goodwin RM, Walker MK, Howlett BG (2011) Honey bee (_Apis mellifera_) distribution and behaviour on hybrid radish (_Raphanus sativus_ L) crops. New Zealand Plant Protection 64: 32-36.
-
Hladun KR, Parker DR, Tran KD, Trumble JT (2013) Effects of selenium accumulation on phytotoxicity, herbivory, and pollination ecology in radish (_Raphanus_ _sativus_ L.). Environmental Pollution 172: 70-75.
-
Garibaldi LA, Steffan-Dewenter I, Winfree R, Aizen MA, Bommarco R, et al. (2013) Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 339(6127): 1608-1611.
-
Spears EE (1983) A direct measure of pollinator effectiveness. Oecologia 57: 196.
-
Davis AR (1992) Evaluating honey bees as pollinators of virgin flowers of Echium plantagineum L.(Boraginaceae) by pollen tube fluorescence. Journal of Apicultural Research 31(2): 83-95.
-
Delaplane KS, Van Der Steen J, Guzman-Novoa E (2013) Standard methods for estimating strength parameters of Apis mellifera colonies. Journal of Apicultural Research 52(1): 1-12.
-
Vaissiere BE, Rodet G, Cousin M, Botella M, Torre- Grossa JP (1996) Pollination effectiveness of honey bees (Hymenoptera: Apidae) in a kiwifruit orchard. Journal of Economic Entomology 89: 453-461.
-
Choudhury B (2000) Radish. In: Vegetables. National Book Trust, New Delhi, India, pp: 133-140.
-
Ratti JT, Garton EO (1996) Research and experimental design. Di dalam: Bookhout TA editor. Research and Management Techniques for Wildlife and Habitats.
-
Batra SW (1977) Bees of India (Apoidea), their behaviour, management and a key to the genera. Oriental Insects 11(3): 289-324.
-
Seltmann KC (2004) Building web-based interactive keys to the hymenopteran families and superfamilies. Uknowledge, UK, pp: 1-74.
-
Karunaratne WAIP, Edirisinghe JP (2008) Keys for the identification of common bees of Sri Lanka. Journal of the National Science Foundation of Sri Lanka 36(1): 69- 89.
-
Buck M, Woodley NE, Borkent A, Wood DM, Pape T, et al. (2009) Key to Diptera families-adults. Manual of Central American Diptera 1: 95-156.
-
Marshall SA, Kirk Spriggs AH, Muller BS, Paiero SM, Yau T, et al. (2017) Key to Diptera families-adults. Suricata 4(1): 267-311.
-
Stanley J, Subbanna ARNS, Mishra KK, Pattanayak A (2018) Butterflies of Almora. Technical bulletin, ICARVPKAS, Almora, Uttarakhand, India, pp: 101.
-
Stanley J, Sah K, Subbanna ARNS, Preetha G, Gupta JP (2017) How efficient is Apis cerana (Hymenoptera: Apidae) in pollinating cabbage, Brassica oleracea var. capitata? Pollination behavior, pollinator effectiveness, pollinator requirement, and impact of pollination. Journal of Economic Entomology 110(3): 826-834.
-
Stanley J, Sah K, Subbanna ARNS (2017) How efficient is the Asian honey bee, Apis cerana in pollinating mustard, Brassica campestris var. toria? Pollination behavior, pollinator efficiency, pollinator requirements and impact of pollination. Journal of Apicultural Research 56(4): 439-451.
-
R Core Team (2020) R Core Team R: a language and environment for statistical computing. Foundation for Statistical Computing, Vienna, Austria.
-
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1).
-
Melathopoulos AP, Cutler GC, Tyedmers P (2015) Where is the value in valuing pollination ecosystem services to agriculture? Ecological Economics 109: 59-70.
-
Tamburini G, Bommarco R, Kleijn D, Van der Putten WH, Marini L (2019) Pollination contribution to crop yield is often context-dependent: a review of experimental evidence. Agriculture, Ecosystems and Environment 280: 16-23.
-
Kremen C (2018) The value of pollinator species diversity. Science 359(6377): 741-742.
-
Horn DJ (1981) Effect of weedy backgrounds on colonization of collards by green peach aphid, Myzus persicae, and its major predators. Environmental Entomology 10(3): 285-289.
-
Jauker F, Bondarenko B, Becker HC, Steffan‐Dewenter I (2012) Pollination efficiency of wild bees and hoverflies provided to oilseed rape. Agricultural and Forest Entomology 14(1): 81-87.
-
Orford KA, Vaughan IP, Memmott J (2015) The forgotten flies: the importance of non-syrphid Diptera as pollinators. Proceedings of Royal Society B 282(1805): 1-6.
-
Free JB, Williams IH (1973) The foraging behaviour of honeybees (_Apis mellifera_ L.) on Brussels sprout (Brassica oleracea L.). Journal of Applied Ecology 10(2): 489-499.
-
Mishra RC, Kumar J, Gupta JK (1987) Effect of mode of pollination on fruit characteristics of okra, _Abelmoschus_ _esculentus_ (L) Moench. Proceedings of Indian National Science Academy B 53: 157–160.
-
Kapoor KS, Dhaliwal HS (1989) Comparative foraging strategies of Apis cerana indica and Apis mellifera L. on cauliflower. Indian Bee Journal 51(3): 99-101.
-
Gary NE (1992) Activities and behavior of honeybees. The hive and the honey bee. Dadant and Sons, Hamilton, IL, USA, pp: 269-373.
-
Gilbert FS (1980) Flower visiting by hoverflies (Syrphidae). Journal of Biological Education 14(1): 70- 74.
-
Mishra RC, Kumar J, Gupta JK (1988) The effect of mode of pollination on yield and oil potential of _Brassica_ _campestris_ L. Var. Sarson with observations on insect pollinators. Journal of Apicultural Research 27(3): 186- 189.
-
Singh J, Agrawal OP, Mishra RC (2006) Foraging rates of different Apis species visiting different parental lines of _Brassica napus_ L. Zoos Print Journal 21(4): 2226-2227.
-
Galen C, Stanton ML (1989) Bumble bee pollination and floral morphology: factors influencing pollen dispersal in the alpine sky pilot, Polemonium viscosum (Polemoniaceae). American Journal of Botany 76(3): 419-426.
-
Herrera CM (1989) Pollinator abundance, morphology, and flower visitation rate: analysis of the “quantity” component in a plant-pollinator system. Oecologia 80(2): 241-248.
-
Mayfield MM, Waser NM, Price MV (2001) Exploring the ‘most effective pollinator principle’ with complex flowers: bumblebees and Ipomopsis aggregata. Annals of Botany 88(4): 591-596.
-
Ivey CT, Martinez P, Wyatt R (2003) Variation in pollinator effectiveness in swamp milkweed, _Asclepias_ _incarnata_ (Apocynaceae). American Journal of Botany 90(2): 214-225.
-
Muchhala N, Potts MD (2007) Character displacement among bat-pollinated flowers of the genus Burmeistera: analysis of mechanism, process and pattern. Proceedings of the Royal Society B: Biological Sciences 274(1626): 2731-2737.
-
Klein AM, Steffan-Dewenter I, Tscharntke T (2003) Fruit set of highland coffee increases with the diversity of pollinating bees. Proceedings of the Royal Society of London Series B 270(1518): 955-961.
-
Neves EL, Viana BF (2011) Pollination efficiency of Apis mellifera Linnaeus, 1758 (Hymenoptera, Apidae) on the monoecious plants Jatropha mollissima (Pohl) Baill. and Jatropha mutabilis (Pohl) Baill. (Euphorbiaceae) in a semi-arid Caatinga area, northeastern Brazil. Brazilian Journal of Biology 71(1): 107-113.
-
Coulin C, Aizen MA, Garibaldi LA (2019) Contrasting responses of plants and pollinators to woodland disturbance. Austral Ecology 44(6): 1040-1051.
-
Prendergast KS (2020) Beyond ecosystem services as justification for biodiversity conservation. Austral Ecology 45(2): 141-143.
-
Aizen MA, Harder LD (2009) The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Current Biology 19(11): 915-918.
-
Hass AL, Kormann UG, Tscharntke T, Clough Y, Baillod AB, et al. (2018) Landscape configurational heterogeneity by small-scale agriculture, not crop diversity, maintains pollinators and plant reproduction in western Europe. Proceedings of the Royal Society B: Biological Sciences 285(1872): 1-10.
-
Dohzono I, Yokoyama J (2010) Impacts of alien bees on native plant-pollinator relationships: A review with special emphasis on plant reproduction. Applied Entomology and Zoology 45(1): 37-47.
- California Red-Legged Frog and Non-Listed Amphibians Response to Non-Native Fish Removal
- Industrial Standardization of the Bio-OS: Algorithmic Codification of Resilience Engineering Guidelines and Version V8 Architecture
- Climate Variability and the Sustainability of Snail Farming in Nigeria: Past Trends, Present Challenges and Potential Outlook
- The Evaluation of the Surveillance System of Anthrax in Gilgit-Baltistan, Pakistan, 2018
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework