The Effects of Moderate-Intensity Inhomogeneous Static Magnetic Fields on Neuromuscular Transmission
The present study focuses on the effects of moderate-intensity inhomogeneous static magnetic fields (SMF) up to 0.73 T on action potentials in Wistar rats. Thirty three animals were divided into three groups: a sham-exposed control group and two exposed groups with exposure to maximum flux densities (Bmax) of 0.24 and 0.73 T SMF. The amplitude of the electrically evoked compound nerve action potentials (CNAP) and compound muscle action potentials (CMAP) were measured for up to 2 h. In the CNAP, the excitation of Aδ fibers was significantly enhanced by both 0.24 T and 0.73 T SMF for 2 h, relative to the sham-exposed control. Furthermore, the CMAP decrement was significantly enhanced by 0.73 T SMF for 1 to 2 h, but not by 0.24 T SMF for up to 2 h, compared with the control. These results suggest that SMF (0.24 and 0.73 T for 2 h) enhances pain perception because the Aδ fibers are responsible for pain transmission. In addition, SMF (0.73 T for 1 to 2 h) may modulate neuromuscular transmission. Thus, the magnetic force produced by SMF could affect the behavior of some types of ion channels associated with Aδ fibers, probably due to SMF-induced modulation of ion/ligand binding and ion transport.
Introduction
Considering the widespread and rapid increase in the static magnetic fields (SMF) used in medical diagnosis and applications, various studies have investigated the effects of acute or chronic exposure to SMF on humans and animals over the past 30-40 years. A number of neuro- and electrophysiological studies on SMF effects, including or focusing on less than 1 T, have been well-reviewed elsewhere [1, 2, 3]. Several studies have reported on significant neurophysiological effects of moderate- intensity SMF ranging from 1 mT to 1 T with exposure duration from 100 sec to 40 days [4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44]. However, only a few studies have reported on the effects of moderate- intensity SMF on action potentials in vivo, probably due to SMF-induced modulation of ion/ligand binding and ion transport [4, 8, 22]. For example, a spatially homogeneous SMF of 0.65 T suppressed the temporal decrease of the muscle tension induced by electrical stimulation of the sartorius muscle in the neuromuscular preparation of the bullfrog [22]. Four possible biochemical mechanisms for the SMF effect on muscle tension were proposed: (i) change of acetylcholine release from presynaptic terminals; (ii) sensitivity change of acetylcholine receptors at the endplate; (iii) change of Ca2+ dynamics; and (iv) change of large molecules and enzymes, e.g. calmodulin molecules, myosin light chain kinase, and ATP molecules [22]. These studies, however, mainly focused on isolated skeletal muscles and nerve fibers in vitro, and almost none on neuromuscular transmission in vivo. Therefore, this study focuses on the in vivo effects of moderate- intensity SMF up to 0.73 T on the rat sciatic nerve fibers of the sartorius muscle.
Materials and Methods
Animals and Groups
Thirty three adult male Wistar rats (body weight 250– 300 g) were used in this study. Wistar rats were purchased from Charles River Laboratories Japan. The animals were housed individually in the same room, with a 12-h light/dark cycle (lights on: 7 a.m.) at a temperature of 25±0.5°C, and a relative humidity of 50±5%. Animals were fed standard rodent chow and tap water ad libitum. All experimental procedures performed on the animals were in strict accordance with the Guide for the Care and Use of Laboratory Animals (7th ed. 1996, US National Academy Press, Washington, DC, USA) and approved by the Animal Ethics Committee in Chiba University in Japan. The animals were divided into three groups: I) sham- exposed control group (control group); II) SMF-exposed group with inhomogeneous exposure to maximum flux densities (_B_max) of 0.24 T (0.24 T-exposed group); and III) SMF-exposed group with _B_max of 0.73 T (0.73 T-exposed group), which were further subdivided into six subgroups for measuring either kind of action potentials (each subgroup had 5-6 animals): Ia) control group for measuring compound nerve action potentials (CNAP); IIa) 0.24 T-exposed group for measuring CNAP; IIIa) 0.73 T- exposed group for measuring CNAP; Ib) control group for measuring compound muscle action potentials (CMAP); IIb) 0.24 T-exposed group for measuring CMAP; IIIb) 0.73 T-exposed group for measuring CMAP. The electrical stimulation and recording of both action potentials was performed for approximately 5 minutes at 1-h and 2-h exposure time in the SMF-exposed group under SMF exposure and in the sham-exposed group under non-SMF exposure (except for the Earth's magnetic field).
Magnetic Field
The SMF was generated by two neodymium–iron– boron (NdFeB) magnet assemblies (TDK, Japan), detailed specifications have been described elsewhere [42]. One magnet assembly was constructed of a pair of rectangular NdFeB magnets (magnet dimension L ×W × D = 15 × 15 × 2.4 cm; B_max 0.26 T on the surface of the magnet). The upper and lower magnetic plates, with oppositepolarities vertically attracting each other through a 5 cm air gap, were fixed parallel to both sides of the stainless frame. In the other magnet assembly, a pair of stronger NdFeB magnetic plates (L ×W × D = 15 × 15 × 6 cm; _B_max 0.75 T) was used with the same air gap of 5 cm. The magnet assemblies in the sham-exposed control group consisted of the same material but were not magnetized. The rear part of the body of the anesthetized animal, which was placed on an animal bed holder (L ×W × D = 30 cm × 10 cm × 0.5 mm), was located on the lower magnetic plate (North seeking pole) inside either type of a magnet assembly (Figure 1). The front part of the body, which was placed on a supporting box, was located outside the magnet assembly. The direction of long axis of the body (their body length of about 30 cm) was oriented orthogonal to the attracting force of magnetic plates. The spatial distribution of the SMF was measured along the _y- axis at the animal site using a magnetic field meter (model 4048 with a Hall probe sensor A-4048-002, Bell Technologies, USA). The whole body was exposed to spatially inhomogeneous SMF (Figure 1a,b). The distance between the magnet surface and the body surface was 0.5 mm due to the depth of the animal bed holder. The B_max _applied to the body was 0.24 T (Figure 1a) or 0.73 T (Figure 1b). The gradient in the whole body was calculated on the basis of measured field strengths (B) as:
$$ G = \partial B / \partial y \tag {1} $$
The force product (FP) was defined as:
FP = BG (2) In the magnet assembly of B_max = 0.24 T, the maximum values of magnetic gradient (_G_max), and force product (_FP_max) were 12.53 T m-1 and 0.97 T2m-1, respectively (Figure 1a). In the magnet assembly of _B_max = 0.73 T, the _G_max, and _FP_max values were 26.73 T m-1 and 7.23T_2_m-1, respectively (Figure 1b). In both magnet assemblies the _G_max, and _FP_max values were located near the edge of the magnet. The _anesthesia was maintained during the entire experimental period for at least 2 h and the animal was restrained in the magnetic assembly. The ambient temperature in the magnet was maintained at 25±0.5°C with the relative humidity of 50± 5%.
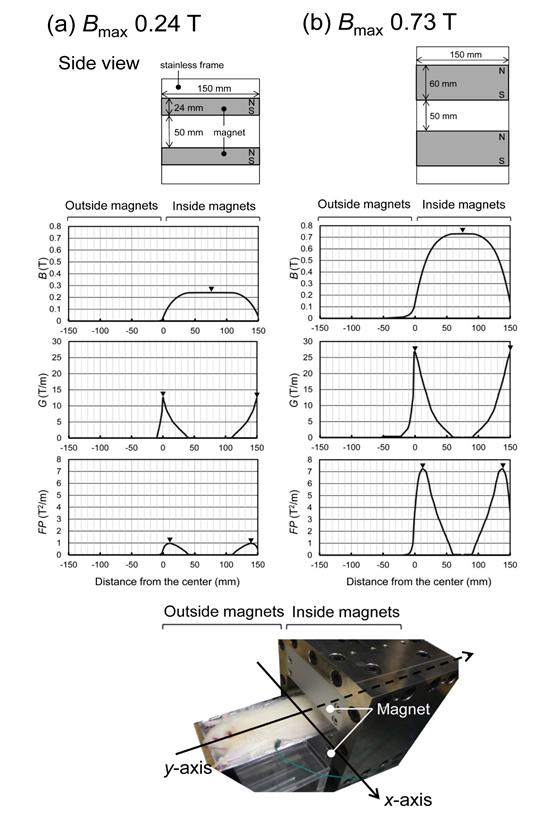
Figure 1: Schematic view of exposure of a ratto a magnet assembly with a vertically oriented and spatial distribution of SMF in two types of magnet assemblies along the y-axis. The animals were exposed to a spatially inhomogeneous SMF of B_max 0.24 T (a) or 0.73 T (b) at the mid-pole field along the _y-axis. Upper row, magnetic flux density (B) values; middle row, magnetic gradient (G) values; lower row, force product (FP) values; and ▼, respective peak values.
Compound Nerve Action Potentials (CNAP)
Wistar rats were anesthetized with urethane (1.0 g kg-1, i.p.; ethyl carbamate; Wako Pure Chemical, Japan). The rats were placed in prone position, a lateral skin incision was made on the dorsal side of the right hind limb, and then a sciatic nerve and a gastrocnemius nerve were exposed. One of a bipolar electrode was placed on the sciatic nerve for applying repetitive electrical nerve stimulation and the other bipolar electrode was attached to the medial gastrocnemius nerve for measuring compound nerve action potentials (CNAP). The distance between the stimulating and recording electrodes was approximately 3 cm. A reference ground electrode was attached to the body of each animal. The reference ground electrode was connected to the earth terminal of the power source to eliminate stray current interference. All electrodes were made of pure platinum, which is inert and non-ferromagnetic material (molar magnetic susceptibility χ_mol (cgs) = 193 ×10-6cm3mol-1 at 295 K) [46] and, therefore, is apparently neither attracted nor repelled by a magnet. After attaching the electrodes to the nerves, the nerves were covered with liquid paraffin, and were sealed with Para film (Bemis Flexible Packaging, USA) to prevent desiccation. In each animal, the four limbs and the lead wires were tightly fixed to an animal bed holder using strings in order to avoid motions associated with muscle contractions during the repetitive electrical nerve stimulation. The anesthesia lasted for at least 2 h during the measuring experiment. The sciatic nerve was electrically stimulated during exposure to a non-homogeneous SMF of _B_max 0.24 T or 0.73 T. Except for a strong SMF (2-8 T) generated by a superconducting magnet, a similar procedure has been described elsewhere [47]. Briefly, a measurement instrument (Neuropack MEB-2200, Nihon Kohden, Japan) was used to stimulate the nerve bundle and to record the CNAP. Repetitive electrical nerve stimulation was applied with intensities ranging from 1, 3, 5 and 7 mA, with a pulse width of 0.1 ms and a pulse repetition rate of 3 Hz. The signal-to-noise ratio (SNR) was improved by averaging 20 repetitive recordings. Data were collected from the medial gastrocnemius nerve in the right side of 15 animals for approximately 5 minutes at 1-h and 2-h exposure time, in which stability was sufficient to allow detailed analysis. The animals were randomly divided into the above mentioned three subgroups: Ia) control group (_n = 5); IIa) 0.24 T-exposed
group (n = 5); and IIIa) 0.73 T-exposed group (n = 5) for up to 2 h. The values of CNAP were measured for these three groups.
Compound Muscle Action Potentials (CMAP)
The animals used in CMAP recordings were different individuals from those used in CNAP recordings. With the anesthetized rat in a prone position, the left sciatic nerve was exposed through a lateral skin incision on the dorsal side of the left mid-thigh. A measurement instrument (Neuropack MEB-2200, Nihon Kohden, Japan) was also used to stimulate the sciatic nerve and to record the compound muscle action potentials (CMAP). For stimulation, a bipolar electrode was attached to the sciatic nerve. The sciatic nerve was electrically stimulated during exposure to a non-homogeneous SMF of _B_max 0.24 T or 0.73 T, using 10 repetitive pulses of 8 mA, 0.5 ms and 3 Hz, as a supramaximal stimulus. The CMAP was recorded from the left gastrocnemius muscle using a platinum concentric needle electrode (NM-030T, Nihon Kohden).
The distance between the stimulating and recording electrodes was approximately 6 cm. The SNR was improved by averaging 10 repetitive recordings. All other procedures were almost the same as for the experiment of CNAP. We calculated the values of “the relative average amplitude” and “the decrement rate” of CMAP. The values of the relative average amplitude were determined by signal averaging the successive responses elicited by repetitive ten pulses and were calculated relative to the corresponding pre-exposure values (pre-exposure baseline value = 100%). The CMAP decrement rate (%) was simply calculated as follows: Decrement rate %) = 1 − 𝑃min 𝑃1 ) × 100 (3) Where _P_1 is the first pulse stimulus-induced amplitude and _P_min is the minimum amplitude induced by the following pulse stimulus (Figure 2).
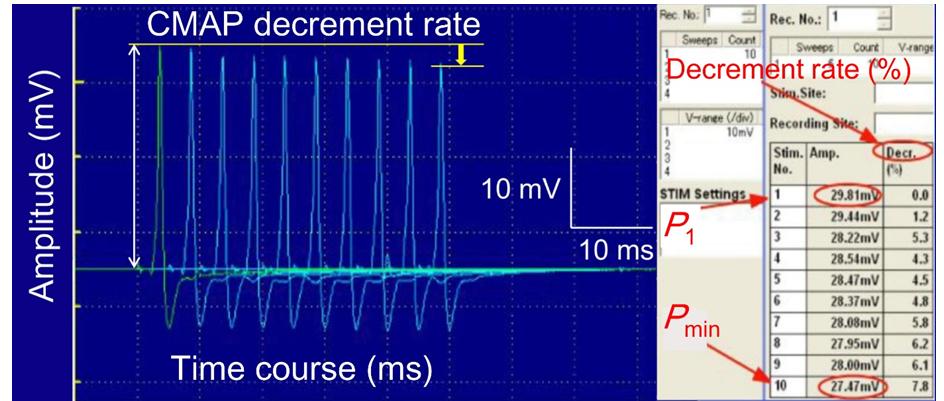
up to 2 h. The values of CMAP were measured for these three groups.
Data Processing
The results are expressed as mean ± standard error of the mean (SEM). Comparisons between groups over time were made using two-way analysis of variance with repeated measures, followed by the Wilcoxon rank sum test for pairwise comparisons. Significance was set at P< 0.05.
Results
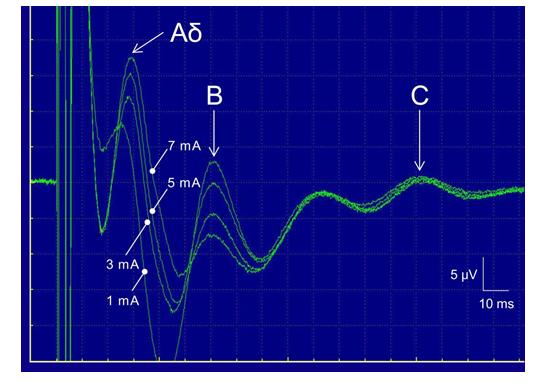
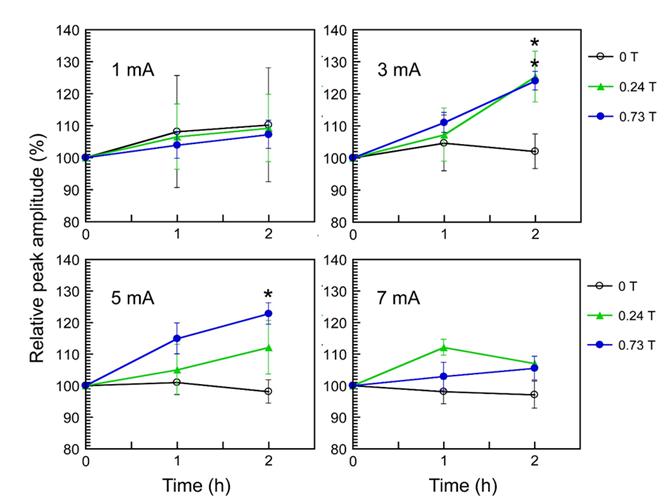
Multiple peaks were observed in the recorded CNAP (Figure 3). Nerve fibers were classified into several types (Aα, Aβ, Aγ, Aδ, B, and C) according to their conduction velocities and functions. We identified three peaks corresponding to the Aδ, B, and C fibers Figure 3). The respective conduction velocities of the Aδ, B, and C fibers were 13.6, 3.8 and 0.5 ms-1. To investigate the SMF effects on pain-related nerve fibers, we focused on two kinds of sensory nerve fibers, that is, Aδ and C fibers, and analyzed their relative peak amplitude. Myelinated Aδ fibers respond to stimuli such as cold and pressure, and as a nociceptor, Aδ fibers convey fast pain information [48]. Slowly-conducting, unmyelinated C fibers, by contrast, carry slow pain [48]. The values of “the relative peak amplitude” of Aδ and C fibers were calculated as the relative change in the peak amplitude, relative to the corresponding peak pre-exposure values (pre-exposure baseline value = 100%). Significant results of SMF effects on the relative peak amplitude of Aδ fibers were obtained in the range of moderate-intensity electric stimulation applied (3 and 5 mA). At 2-h exposure, both _B_max 0.24 T and 0.73 T SMF caused significant increase in the relative peak amplitude of Aδ fibers at 3 mA stimulation Figure 4). In addition, at 2-h exposure, _B_max 0.73 T SMF significantly elevated the relative peak amplitude of Aδ fibers at 5 mA stimulation (Figure 4). Neither _B_max 0.24 T nor _B_max 0.73 T SMF exposure for up to 2 h induced any significant changes in the relative peak amplitude of C fibers in response to any electrical stimulation used in this study (Table 1).
| Electrical stimulus intensity | Exposure duration | |||
|---|---|---|---|---|
| 0 h | 1 h | 2h | ||
| 0 T (Control) | 1 mA | 100.0 ± 0.0 | 104.8 ± 13.8 | 109.5 ± 13.8 |
| 0 T (Control) | 3mA | 100.0 ± 0.0 | 129.2 ± 18.6 | 137.5 ± 20.5 |
| 0 T (Control) | 5mA | 100.0 ± 0.0 | 117.0± 11.8 | 128.3 ± 13.5 |
| 0 T (Control) | 7 mA | 100.0 ± 0.0 | 125.8 ± 3.4 | 134.9 ± 3.4 |
| 0.24 T | 1 mA | 100.0 ± 0.0 | 110.3 ± 19.5 | 97.7 ± 17.7 |
| 0.24 T | 3mA | 100.0 ± 0.0 | 120.4 ± 10.1 | 134.7 ± 22.8 |
| 0.24 T | 5mA | 100.0 ± 0.0 | 123.1 ± 9.6 | 120.0 ± 8.9 |
| 0.24 T | 7 mA | 100.0 ± 0.0 | 117.1 ± 8.7 | 112.2 ± 8.8 |
| 0.73 T | 1 mA | 100.0 ± 0.0 | 108.7 ± 16.5 | 110.9 ± 17.5 |
| 0.73 T | 3 mA | 100.0 ± 0.0 | 131.5 ± 12.4 | 135.2 ± 12.4 |
| 0.73 T | 5 mA | 100.0 ± 0.0 | 130.8 ± 8.3 | 132.3 ± 8.3 |
| 0.73 T | 7 mA | 100.0 ± 0.0 | 131.3 ± 2.3 | 129.9 ± 8.1 |
| Exposure | Electrical stimulus intensity | Exposure duration | ||
| Exposure | Electrical stimulus intensity | 0 h | 1 h | 2h |
| 0 T (Control) | 1 mA | 17.9 ± 0.1 | 17.8 ± 0.1 | 17.8± 0.2 |
| 0 T (Control) | 3mA | 17.8 ± 0.1 | 17.7 ± 0.1 | 17.8 ± 0.1 |
| 0 T (Control) | 5 mA | 17.8 ± 0.2 | 17.9 ± 0.2 | 17.8± 0.2 |
| 0 T (Control) | 7 mA | 17.7 ± 0.2 | 17.8 ± 0.1 | 17.7 ± 0.2 |
| 0.24 T | 1 mA | 17.8 ± 0.1 | 17.8 ± 0.1 | 17.8 ± 0.1 |
| 0.24 T | 3mA | 17.8 ± 0.2 | 17.7 ± 0.2 | 17.8± 0.1 |
| 0.24 T | 5 mA | 17.7 ± 0.1 | 17.8 ± 0.1 | 17.7 ± 0.1 |
| 0.24 T | 7 mA | 17.8 ± 0.2 | 17.9 ± 0.2 | 17.8± 0.1 |
| 0.73 T | 1 mA | 17.8 ± 0.1 | 17.8 ± 0.1 | 17.9 ± 0.2 |
| 0.73 T | 3mA | 17.9 ± 0.2 | 17.8 ± 0.2 | 17.8± 0.1 |
| 0.73 T | 5 mA | 17.8 ± 0.1 | 17.9 ± 0.1 | 17.7 ± 0.1 |
| 0.73 T | 7 mA | 17.8 ± 0.1 | 17.7 ± 0.1 | 17.8± 0.2 |
Table 1: Changes in the relative peak amplitude (%) of C fibers in response to SMF exposure Table 1: Changes in the relative peak
The values of the response latency from stimulus to the peak amplitude of Aδ fibers Table 2) and C fibers Table 3) were also analyzed. No significant results of SMF effects on the latency of Aδ and C fibers were obtained in any case (Tables 2 & 3).
The CMAP decrement was significantly enhanced by _B_max 0.73 T SMF during 1- to 2-h exposure period, but not by _B_max 0.24 T SMF during the entire exposure period of 2 h, compared with the unexposed control (Figure 5). The relative average amplitude of CMAP was not affected by both SMF exposures (Table 4).
| Exposure | Electrical stimulus intensity | Exposure duration | ||
| Exposure | Electrical stimulus intensity | 0 h | 1 h | 2h |
| 0 T (Control) | 1 mA | 122.2±0.2 | 122.1±0.2 | 122.2±0.2 |
| 0 T (Control) | 3mA | 122.1±0.2 | 122.2±0.2 | 122.2±0.1 |
| 0 T (Control) | 5 mA | 122.2±0.2 | 122.2±0.2 | 122.3±0.2 |
| 0 T (Control) | 7 mA | 122.3±0.1 | 122.2±0.2 | 122.2±0.1 |
| 0.24T | 1 mA | 122.2±0.2 | 122.3±0.2 | 122.2±0.2 |
| 0.24T | 3mA | 122.1±0.2 | 122.2±0.1 | 122.2±0.2 |
| 0.24T | 5 mA | 122.2±0.2 | 122.1±0.2 | 122.3±0.2 |
| 0.24T | 7 mA | 122.1±0.2 | 122.2±0.2 | 122.2±0.1 |
| 0.73 T | 1 mA | 122.3±0.2 | 122.2±0.2 | 122.1±0.2 |
| 0.73 T | 3mA | 122.1±0.2 | 122.1±0.2 | 122.2±0.2 |
| 0.73 T | 5 mA | 122.2±0.1 | 122.2±0.2 | 122.1±0.2 |
| 0.73 T | 7 mA | 122.3±0.2 | 122.2±0.2 | 122.1±0.2 |
Table 2: Changes in the latency (ms) of C fibers in response to SMF exposure.
| Exposure duration | |||
|---|---|---|---|
| Exposure | 0 h | 1 h | 2h |
| 0 T (Control) | 100.0±0.0 | 112.7±14.9 | 108.2±17.7 |
| 0.24 T | 100.0±0.0 | 108.9±15.9 | 106.9±12.7 |
| 0.73 T | 100.0±0.0 | 115.2±12.3 | 113.7±12.1 |
Table 3: Changes in the relative amplitude (%) of CMAP in response to SMF exposure.
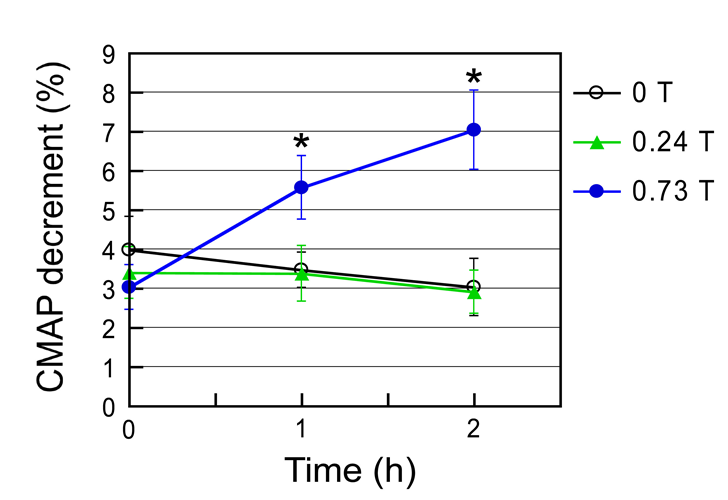
The major finding of this study is that excitation of Aδ fibers was significantly enhanced by both 0.24 T and 0.73 T SMF for 2 h, relative to the sham-exposed control. These results in vivo implies that exposure to moderate- intensity inhomogeneous SMF enhances pain perception, through the reduction of pain threshold, because Aδ fibers are responsible for pain transmission. In contrast, our previous results in vitro (excised frog sciatic nerve preparation) suggested that _B_max 0.70 T SMF could increase pain threshold by decreasing nerve conduction velocity of C fibers alone, but not of Aδ fibers [42]. In the current study we used urethane as an anesthetic agent because urethane is widely used to investigate neurovascular coupling due to its balanced effect on neurotransmitters [49], although the effects on neurotransmitter-gated ion channels, including nicotinic acetylcholine receptors, were reported [50]. Ideally, if possible, the experiment should be performed in conscious animals without the interference of anesthetics.
Anesthesia by itself can modulate not only pain perception but also probably excitation of Aδ fibers. However, the finding that SMF increases nerve fibers excitability and modulate neuromuscular transmission has more important potential implication than other pain perception processes. Another study in vitro (excised adult guinea pig spinal cords) showed that exposure to a homogeneous SMF of 0.5 T induced a decrease in the amplitude of CNAP without a change in the response latency during SMF exposure [23]. A maximum effect was evident 1–2 min after the SMF was applied, with a return to baseline within 1 min after the SMF was removed. The results were explained by a conduction block in the small axon subpopulation owing to the SMF effect on voltage- activated Na+ channels. The relative selectivity of the SMF was believed to occur because of the relatively greater density of Na+ channels present in smaller axons. These results in vitro are consistent with our previous results on the SMF effect on nerve conduction in vitro [42], but are inconsistent with our present results on the SMF effect on neuromuscular transmission in vivo. It has been reviewed that in vivo nerve conduction studies have failed to establish a link between in vitro effects and the analgesic responses observed in pain studies [51]. It seems likely that the SMF effects on pain perception in vitro are often different from those in vivo, depending on the physiological conditions of the nerve membrane excitation. Although the mechanistic reasons for this difference in the threshold have yet to be clarified, SMF could affect the behavior of some types of ion channels associated with Aδ and C fibers. The other significant finding is that the CMAP decrement was significantly enhanced by 0.73 T SMF for 1 to 2 h, compared with the control. From the viewpoint of the electrophysiological diagnosis, the enhanced CMAP decrement in repetitive electrical nerve stimulation is related to dysfunction of neuromuscular transmission [52]. This implies that SMF may cause dysfunction of neuromuscular transmission. More specifically here, the CMAP changes roughly indicate the ratio of “the number of acetylcholine (ACh) receptor responded to Ach release from presynaptic terminals” to “the amount of ACh release” [53]. Therefore, the SMF-enhanced CMAP decrement implicates that SMF may increase the ACh release from presynaptic terminals and/or decrease the sensitivity of ACh receptor because the ACh release from nerve terminals is the cause of muscle contraction. As we mentioned in the introduction, the effects of a moderate- intensity homogeneous SMF on muscle tension were found using in vitro neuromuscular preparation [22]. In considering the biochemical mechanisms, our results might be in accordance with the mechanism(s) that modulation of muscle tension appears to be a pathway(s) through which (i) ACh release from presynaptic terminals and/or (ii) sensitivity change of ACh receptors at the endplate can influence CMAP. Additional experiments with the pharmacological agents (for instance, ACh release inhibitors) and muscle tension measurements are needed to make it clear. Despite the pain-related response induced by moderate-intensity heterogeneous SMF [16, 17, 28, 30, 31, 37, 39, 42, 43], however, there has been little documented on the evident effects of SMF on neuromuscular transmission, except for the present study. Our findings indicate that exposure to moderate- intensity inhomogeneous SMF could transiently affect such aspects of the neurophysiological response as enhanced excitation of Aδ fibers and enhanced CMAP decrement, resulting in increased pain perception. Furthermore, SMF (0.73 T for 1 to 2 h) may modulate neuromuscular transmission. Thus, SMF could affect the behavior of some types of ion channels associated with Aδ fibers. Several studies have tested whether activation or inactivation of excitable tissues and cells by moderate- intensity SMF [5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45] and their theoretical models for the action of SMF on excitable tissues and cells have been proposed [14, 54, 55, 56, 57]. To date, however, there is insufficient direct experimental evidence pertaining to these theoretical models and the discrepancy of our present results with other studies cannot yet be explained by theoretical models. Further studies are required to better understand the underlying mechanisms of SMF effects on neuromuscular transmission and pain perception.
Conclusion
Both 0.24 T and 0.73 T SMF for 2 h significantly enhanced the excitation of Aδ fibers in the CNAP, compared with the sham-exposed control. Furthermore, 0.73 T SMF for 1 to 2 h significantly enhanced the CMAP decrement, but 0.24 T SMF for up to 2 h did not change it significantly, compared with the control. These results suggest that SMF (0.24 and 0.73 T for 2 h) enhances pain perception because the Aδ fibers are responsible for pain transmission. In addition, SMF (0.73 T for 1 to 2 h) may modulate neuromuscular transmission. Thus, SMF could affect the behavior of some types of ion channels associated with Aδ fibers, probably due to SMF-induced modulation of ion/ligand binding and ion transport.
Conflicts of Interest
The authors declare that we have no conflicts of interest on submission of manuscript for publication.
References
-
Ueno S, Shigemitsu T (2006) Biological effects of static magnetic fields. Handbook of Biological Effects of Electromagnetic Fields: Bioengineering and Biophysical Aspects of Electromagnetic Fields 3rd(edn), CRC Press, Boca Raton, pp: 203-259.
-
Health Protection Agency (2008) Static magnetic fields.The Report from the Independent Advisory Group on Non-ionising Radiation, Documents of the Health Protection Agency.Radiation, Chemical and Environmental Hazards, Oxfordshire, pp: 78-83.
-
Ueno S, Okano H (2011) Static, low-frequency, and pulsed magnetic fields in biological systems. Electromagnetic Fields in Biological Systems, CRC Press, Boca Raton, pp: 115-196.
-
Tamaki T, Yoshioka T, Nakano S (1987) Effect of magnetic field on the contractility and glycogen content in neuromuscular preparation. Tokai J Exp Clin Med 12(1): 55-59.
-
Rosen AD, Lubowsky J (1987) Magnetic field influence on central nervous system function. Exp Neurol 95(3): 679-687.
-
Rosen AD, Lubowsky J (1990) Modification of spontaneous unit discharge in the lateral geniculate body by a magnetic field. Experimental Neurology 108(3): 261-265.
-
Azanza MJ (1989) Steady magnetic fields mimic the effect of caffeine on neurons. Brain Res 489(1): 195- 198.
-
Rosen AD (1992) Magnetic field influence on acetylcholine release at the neuromuscular junction. Am J Physiol 262(6): 1418-1422.
-
Rosen AD (1993) Membrane response to static magnetic fields: effect of exposure duration. Biochim Biophys Acta 1148(2): 317-320.
-
Rosen AD (1993) A proposed mechanism for the action of strong static magnetic fields on biomembranes. Int J Neurosci 73(1-2): 115-119.
-
Rosen AD (1994) Threshold and limits of magnetic field action at the presynaptic membrane. Biochim Biophys Acta 1193(1): 62-66.
-
Rosen AD (1996) Inhibition of calcium channel activation in GH3 cells by static magnetic fields. Biochim Biophys Acta 1282(1): 149-155.
-
Rosen AD (2003) Effect of a 125 mT static magnetic field on the kinetics of voltage activated Na+ channels in GH3 cells. Bioelectromagnetics 24(7): 517-523.
-
Rosen AD (2003) Mechanism of action of moderate- intensity static magnetic fields on biological systems. Cell Biochem Biophys 39(2): 163-173.
-
Ayrapetyan SN, Grigorian KV, Avanesian AS, Stamboltsian KV (1994) Magnetic fields alter electrical properties of solutions and their physiological effects. Bioelectromagnetics 15(2): 133- 142.
-
Cavopol AV, Wamil AW, Holcomb RR, McLean MJ (1995) Measurement and analysis of static magnetic fields that block action potentials in cultured neurons. Bioelectromagnetics 16(3): 197-206.
-
McLean MJ, Holcomb RR, Wamil AW, Pickett JD, Cavopol AV (1995) Blockade of sensory neuron action potentials by a static magnetic field in the 10 mT range. Bioelectromagnetics 16(1): 20-32.
-
Trabulsi R, Pawlowski B, Wieraszko A (1996) The influence of steady magnetic fields on the mouse hippocampal evoked potentials in vitro. Brain Res 728(1): 135-139.
-
Pacini S, Vannelli GB, Barni T, Ruggiero M, Sardi I, et al. (1999) Effect of 0.2 T static magnetic field on human neurons: remodeling and inhibition of signal transduction without genome instability. NeurosciLett 267(3): 185-188.
-
Sonnier H, Kolomytkin OV, Marino AA (2000) Resting potential of excitable neuroblastoma cells in weak magnetic fields. Cell Mol Life Sci 57(3): 514-520.
-
Wieraszko A (2000) Dantrolene modulates the influence of steady magnetic fields on hippocampal evoked potentials in vitro. Bioelectromagnetics 21(3): 175-182.
-
Satow Y, Matsunami K, Kawashima T, Satake H, Huda K (2001) A strong constant magnetic field affects muscle tension development in bullfrog neuromuscular preparations. Bioelectromagnetics 22(1): 53-59.
-
Coots A, Shi R, Rosen AD (2004) Effect of a 0.5-T static magnetic field on conduction in guinea pig spinal cord.J NeurolSci 222(1-2): 55-57.
-
Hirai T, Yoneda Y (2004) Functional alterations in immature cultured rat hippocampal neurons after sustained exposure to static magnetic fields. J Neurosci Res 75(2): 230-240.
-
Hirai T, Taniura H, Goto Y, Tamaki K, Oikawa H, et al. (2005) Counteraction by repetitive daily exposure to static magnetism against sustained blockade of N- methyl-D-aspartate receptor channels in cultured rat hippocampal neurons. J Neurosci Res 80(4): 491-500.
-
Hirai T, Taniura H, Goto Y, Ogura M, Sng JC, et al. (2006) Stimulation of ubiquitin-proteasome pathway through the expression of amidohydrolase for N- terminal asparagine (Ntan1) in cultured rat hippocampal neurons exposed to static magnetism. J Neurochem 96(6): 1519-1530.
-
Prina-Mello A, Farrell E, Prendergast PJ, Campbell V, Coey JM (2006) Influence of strong static magnetic fields on primary cortical neurons. Bioelectromagnetics 27(1): 35-42.
-
Sándor K, Helyes Z, Gyires K, Szolcsányi J, László J (2007) Static magnetic field-induced anti-nociceptive effect and the involvement of capsaicin-sensitive sensory nerves in this mechanism. Life Sci 81(2): 97- 102.
-
Shen JF, Chao YL, Du L (2007) Effects of static magnetic fields on the voltage-gated potassium channel currents in trigeminal root ganglion neurons. NeurosciLett 415(2):164-168.
-
Gyires K, Zádori ZS, Rácz B, László J (2008) Pharmacological analysis of inhomogeneous static magnetic field-induced antinociceptive action in the mouse. Bioelectromagnetics 29(6): 456-462.
-
László J, Reiczigel J, Székely L, Gasparics A, Bogár I, et al. (2007) Optimization of static magnetic field parameters improves analgesic effect in mice. Bioelectromagnetics 28(8): 615-627.
-
Todorović D, Kalauzi A, Prolić Z, Jović M, Mutavdžić D (2007) A method for detecting the effect of magnetic field on activity changes of neuronal populations of _Morimusfunereus_(Coleoptera, Cerambycidae). Bioelectromagnetics 28(3): 238-241.
-
Nikolić L, Kartelija G, Nedeljković M 2008) Effect of static magnetic fields on bioelectric properties of the Br and N1 neurons of snail Helix pomatia. Comp Biochem Physiol A MolIntegr Physiol 151(4): 657- 663.
-
Nikolić L, Todorović N, Zakrzewska J, Stanić M, Rauš S, et al. (2012) Involvement of Na+/K+ pump in fine modulation of bursting activity of the snail Br neuron by 10 mT static magnetic field. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 198(7): 525- 540
-
Nikolić L, Bataveljić D, Andjus PR, Nedeljković M, Todorović D, Janać B 2013) Changes in the expression and current of the Na+/K+ pump in the snail nervous system after exposure to a static magnetic field. J Exp Biol 216(18): 3531-3541.
-
Yeh SR, Yang JW, Lee YT, Tsai LY (2008) Static magnetic field expose enhances neurotransmission in crayfish nervous system. Int J Radiat Biol 84(7): 561- 567.
-
Antal M, László J (2009) Exposure to inhomogeneous static magnetic field ceases mechanical allodynia in neuropathic pain in mice. Bioelectromagnetics 30(6): 438-445.
-
Gmitrov J (2010) Static magnetic field blood pressure buffering, baroreflex vs. vascular blood pressure control mechanism. Int J Radiat Biol 86(2): 89-101.
-
Kovács-Bálint Z, Csathó A, László JF, Juhász P, Hernádi I (2011) Exposure to an inhomogeneous static magnetic field increases thermal pain threshold in healthy human volunteers. Bioelectromagnetics 32(2): 131-139.
-
Oliviero A, Mordillo-Mateos L, Arias P, Panyavin I, Foffani G, et al. (2011) Transcranial static magnetic field stimulation of the human motor cortex. J Physiol 589(20): 4949-4958.
-
Spasić S, NikolićL, Mutavdžić D 2011) Effect of a static magnetic field on the fractal complexity of bursting activity of the Br neuron in the snail detected by factor analysis. Arch Biol Sci Belgr 63: 177-183.
-
Okano H, Ino H, Osawa Y, Osuga T, Tatsuoka H (2012) The effects of moderate-intensity gradient static magnetic fields on nerve conduction. Bioelectromagnetics 33(6): 518-526.
-
Kiss B, Gyires K, Kellermayer M, László JF (2013) Lateral gradients significantly enhance static magnetic field-induced inhibition of pain responses in mice-a double blind experimental study. Bioelectromagnetics 34(5): 385-396.
-
Narinyan LY, Ayrapetyan GS, Ayrapetyan SN (2013) Age-dependent magnetosensitivity of heart muscle ouabain receptors. Bioelectro magnetics 34(4): 312- 322.
-
Lu XW, Du L, Kou L, Song N, Zhang YJ, et al. (2015) Effects of moderate static magnetic fields on the voltage-gated sodium and calcium channel currents in trigeminal ganglion neurons. ElectromagnBiol Med 34(4): 285-292.
-
Martienssen W, Warlimont H (2005) The elements: Springer Handbook of Condensed Matter and Materials Data, Springer-Verlag, New York, pp: 141.
-
Sekino M, Tatsuoka H, Yamaguchi S, Eguchi Y, Ueno S (2006) Effects of strong static magnetic fields on nerve excitation. IEEE Trans Magn 42(10): 3584- 3586.
-
Basbaum AI, Bautista DM, Scherrer G, Julius D (2009) Cellular and molecular mechanisms of pain. Cell 139(2): 267-284.
-
Masamoto K, KannoI (2012) Anesthesia and the quantitative evaluation of neurovascular coupling. J Cereb Blood Flow Metab 32(7): 1233-1247.
-
Hara K, Harris RA (2002) The anesthetic mechanism of urethane: the effects on neurotransmitter-gated ion channels. Anesth Analg 94(2): 313-318.
-
Laakso L, Lutter F, Young C (2009) Static magnets: what are they and what do they do? Rev Bras Fisioter 13(1): 10-23.
-
Rich MM (2006) The control of neuromuscular transmission in health and disease. Neuroscientist 12(2): 134-142.
-
Elmqvist D, Mattsson C, Heilbronn E, Lundh H, Libelius R (1977) Acetylcholine receptor protein,Neuromuscular transmission in immunized rabbits. Arch Neurol 34(1): 7-11.
-
Malmivuo J, Plonsey R (1995) Application of Maxwell's equations in bioelectromagnetism: Bioelectromagnetism: Principles and Applications of Bioelectric and Biomagnetic Fields, Oxford University Press, New York, pp: 557-564.
-
St Pierre TG, Dobson J (2000) Theoretical evaluation of cell membrane ion channel activation by applied magnetic fields. Eur Biophys J 29(6): 455-456.
-
Hinch R, Lindsay KA, Noble D, Rosenberg JR (2005) The effects of static magnetic field on action potential propagation and excitation recovery in nerve. Prog Biophys Mol Biol 87(2-3): 321-328.
-
Pilla AA (2007) Mechanisms and therapeutic applications of time-varying and static magnetic fields: Handbook of Biological Effects of Electromagnetic Fields, 3rd edition, CRC Press, Boca Raton, pp: 351-411.
- Solution-Processed Chiral Perovskites for Biomedical Applications
- Nanotechnology in Health Chemistry and Medicine: Current Challenges and Future Directions
- Human Exposure to Micro- and Nanoplastics: Pathways, Toxicity, and Intervention Strategies
- Exosome Nanomedicine for Cancer Therapy
- Micro and Nanoplastics–Plastisphere, Biotoxicity, Impact on Human Health, and Mitigation Strategies
- Process Validation of Cefixime Powder for Suspension Dosage Form, 50 mL