Priming of Vigna mungo (Black Gram) Seeds with Magnetic Hematite Nano-Particle Ameliorates Salinity and Drought Stress via Modulation of Physiochemical Attributes
Vigna mungo (black gram) is a leguminous crop often cultivated in many areas of tropics, for protein-rich edible seeds. Drought and salinity are the major abiotic stress which hampers the growth, development and yield of crop plants including black gram. The present study aimed to investigate the efficacy of magnetic hematite nanoparticles on the stress tolerance potential of Vigna mungo against salinity and drought stress. We found that the treatment of magnetic hematite nanoparticles (Fe2O3) helped to improve black gram tolerance to salt and drought stress under in-vitro as well as in-vivo conditions. The treated plants showed better phenotypic performance in terms of biomass accumulation imposed stress. Moreover, the photosynthetic performance was also higher in treated plants which include total chlorophyll content, stomatal conductance, and maximum photochemical efficiency of PSII as compared to the NNP (non-nanoparticle control) group grown under invivo conditions. In agreement with the results of the increased antioxidant enzyme activity, the V. mungo plants treated with nanoparticles had much less membrane damage, electrolyte leakage, and a greater relative water content and proline content. Further experiments revealed that under imposed stress, the treated V. mungo plants had significantly lower Na+ and higher K+ content which correspond to the higher K+/Na+ ratio than the non-treated stressed control plants.
Introduction
The combined challenges of climate change and population growth underscore the need for integrated approaches that prioritize sustainable and resilient agricultural systems. Climate change has profound implications for agriculture, directly affecting crop growth and development, and ultimately overall yield. Mitigating these impacts necessitates a combination of strategies, including the development of climate-resilient crop varieties and, the adoption of ecologically sustainable farming practices to address the complex issues posed by climate change on agriculture.
Abiotic stresses are the most devastating factors to limit crop growth, development and yield. Abiotic stresses represent unfavourable growth conditions for plants, which include salinity drought, flood, nutrient deficiency, temperatures, high and low light intensity [1]. Salinity and drought are among the list of most prominent stresses that affect the growth and development of plants around the world. During salt and drought stress plants face numerous physical, biochemical and molecular changes [2], and it leads to inhibition of plant growth, via disruption of metabolic pathways, and a decrease in quantity and quality of yield [3]. For this reason, it is the need of the hour to develop strategies which can help the plant species cope with environmental stresses to meet the increasing food and nutritional demand around the globe. Vigna mungo is one of the most valuable and important crops in the world, possessing high nutrition due to its rich source of protein, vitamins and minerals [4]. Additionally, V. mungo has a short vegetative period, and the capacity to fix nitrogen, strengthen the soil, and prevent soil erosion [5]. Screening of contemporary research findings suggested that the yield of black gram decreases due to salt and drought stress [6, 7]. The salt and drought stress limit the yield of black gram through negative impact on the growth and physiology. The salt stress leads to a reduction in seed germination, root and shoot lengths, fresh weight, and seedling viability which ultimately affect yield [8].
Nanotechnology has the potential to bring answers to agricultural difficulties, and technological advancements in the 21st century have been achieved in agriculture to address the challenges of food and nutritional security with ecological sustainability [9]. One of the 16 elements necessary for plant growth and reproduction is iron (Fe). For higher growth and output, annual crops need 1.0–1.5 lbs of iron (Fe) per acre as opposed to 80–200 lbs of nitrogen (N) [10]. Due to their potential to convert to the Fe3+ state, which is not present in the soil, plants only provide little support [11]. The use of metal nano-fertilizers is gaining attention as a solution to this problem to increase the effectiveness of nutrient uptake and decrease soil toxicity brought on by the excessive use of synthetic fertilizers [12]. Hematite is a rich supply of iron oxide nanoparticles, which plants use to release iron in a palatable and relatively stable form of oxide [13]. Hematite nanoparticles serve as an effective nano-fertilizer that boosts Fe availability and biomass production at the same time because of their tiny size and large specific surface area. The current work was carried out to evaluate the effectiveness of magnetic hematite nanoparticles towards the development of stress resilience in V. mungo against salt and drought stress in light of the afore mentioned scientific findings.
Experimental Details
Procurement of Vigna Mungo Seeds and Experimentation Site: The seeds of Vigna mungo (Black gram) (var. IPU 13-
1) were procured from the Indian Council of Agriculture Research- Indian Institute of Pulse Research (ICAR-IIPR), Kanpur, Uttar Pradesh, India and the experimentation was carried out at Centre for Tissue culture Technology, Jacob Institute of Biotechnology and Bioengineering, Sam Higginbottom University of Agriculture Technology and Sciences, Prayagraj, Uttar Pradesh, India in year 2023. Chemical Synthesis of Nanoparticle: According to the procedure outlined by Schwertmann U [14], the synthesis of hematite nanoparticles (Fe3O4) was carried out by co- precipitation from an autocatalytic reaction of ferrous (FeSO4.7H2O) and ferric salts (FeCl3) in an alkaline medium. 50 mL of FeSO4.7H2O (0.3M) and 50 mL of FeCl3.6H2O (0.6 M) were thoroughly mixed. Further, NaOH solution (2M) was added slowly to the mixture and vigorous stirring was carried out at 80℃ for precipitation. The pH of the mixture was maintained at 10.5 during the reaction process. The precipitate was then heated at 80°C for 60 minutes, rinsed five times with distilled water and ethanol, and then centrifuged for 10 minutes at 5000 rpm. The precipitate was then washed three times with distilled water and dried in a vacuum oven at 75°C after the supernatant was discarded. Determination of Lethal Dose (LD50) for Nacl and Nano- Particle Under In-vitro Condition: Seeds of Vigna mungo were washed under running tap water in the presence of tween-20 detergent and rinsed with distilled water to remove the trace of detergent, the rinsed seeds were soaked for 24h at different concentrations of NaCl (100 mM, 150 mM, 200 mM, 250 mM, and 300 mM) and Mannitol (100 mM, 150 mM, 200 mM, 250 mM and 300 mM). Simultaneously, to determine the LD50 of magnetic hematite nanoparticles the rinsed seeds were soaked for 1h in the solution containing different concentrations (50 mgL-1, 100 mgL-1, 200 mgL-1, and 300 mgL-1) of magnetic hematite nanoparticles. In both conditions 3 replicates, each of 10 seeds was placed under dark conditions in petri dishes to observe and record the data. The sprouting of seeds was observed regularly and the data of germination percentage, length of radical and plumule were recorded after 10 days of treatments.
Imposition of Salt and Drought under In-vivo Conditions: A 4:1 mixture of soil and manure was used to prepare the planting medium to impose salt and drought stress under in-vivo conditions. An additional 3.0 kg of sowing mix was placed in 8.0-inch-tall earthen pots. After that, the sowing medium was irrigated with 250 mL of NaCl (150 mM) and Mannitol (200 mM) separately to mimic salt and drought stress under in-vivo conditions and placed in a polyhouse for 72 h. After 72 h, the pre-soaked seeds of V. mungo in magnetic hematite nanoparticle (200 ppm) for 12 h were sown in salt and drought-treated sowing medium and placed back in polyhouse for further analysis.
Measurement of In-vitro Growth Parameters
To investigate the impact of stress (salt and drought) and nanoparticle treatment under in-vitro conditions germination percentage, days to sprouting, length of radical and plumule were recorded manually. The length of radical and plumule of Vigna mungo plants was measured using a scale of 30 cm. The days to sprouting were observed daily and counted after 7 days after treatment (DaT). Moreover, the germination percentage was calculated according to the formula given below;
$$\text{Germination}(\%) = \frac{\text{Number of seedlings}}{\text{Total number of seeds sown}} \times 100$$
**Measurement of Plant’s Behaviour Growing under Imposed Stress Condition**
To study the impact of magnetic hematite nanoparticles on the response of plants under imposed stress (salt and drought) under in-vivo conditions different morphological and physico-chemical attributes were measured. The germination percentage, plant height, root length, and shoot length were recorded. Moreover, the physiochemical status of the V. mungo plants was measured in terms of relative water content, malonyldialdehyde, electrolyte leakage, proline content, enzyme activity, total chlorophyll content, Fv/Fm, and transpiration rate.
Morphological Assessment of In-vivo Growing V. Mungo Plants Under Salt and Drought Stress
The height of the V. mungo plants, root length and shoot length was measured from the base of the stem to the apex of the stem after 30 days of treatment. All the parameters were measured manually by using a centimetre scale.
**Physiochemical Analysis of In-vivo Growing V. mungo Plants**
Physiochemical evaluation, which includes measurement of relative water content (RWC), proline content, electrolyte leakage (EL), and malonyldialdehyde (MDA) content in leaf samples taken from all treatments, was performed on well-developed in-vivo growing plants of all treatments. RWC was determined according to Garcia Mata C, et al. [15] by using the following equation:
$$\text{RWC}(\%) = \frac{\text{FW-DW}}{\text{TW-DW}} \times 100$$
Where
FW (fresh weight) was measured at the end of stress imposition.
DW (dry weight) was measured after drying the samples at 75°C for at least 24 h.
TW (turgid weight) was determined after rehydration of leaves for 2 h.
According to Bates LS, et al. [16], the proline content was approximated. Fresh leaf tissue weighing one gram from 45-day-old V. mungo plants was homogenized in 20 mL of 3% (w/v) sulfosalicylic acid, heated at 100°C for 15 minutes, and then cooled at room temperature. After centrifuging the mixture for two minutes at 10,000 g, 2.0 mL of the supernatant was recovered. Additional 2.0 mL of the acid-ninhydrin reagent, which is a 1:1 mixture of glacial acetic acid and supernatant, were heated at 100°C for 60 minutes before the reaction was stopped with an ice bath for 15 minutes. 2.0 mL of toluene was vigorously shaken to extract the proline, and the amount of toluene’s color was determined by recording the absorbance at 520 nm. The concentration of free proline was calculated from the standard curve made by using purified l-proline (Sigma, MO, USA) in $\mu$mol$^{-1}$.
The electrolyte leakage was assessed using the technique described by Sairam RK, et al. [17]. The 45-day-old plants from each treatment had their leaves collected, which were then washed in deionized water for 10 minutes at room temperature. The sample was incubated at 55°C for 30 minutes to measure EC2 after the EC1 of the solution was determined. In addition, the samples were heated in a water bath for one hour while the total conductivity of the solution (EC3) was measured. The EC was calculated and depicted in percentage according to the formula given below;
$$\text{Electrolyte Leakage} = \left[ \frac{\text{EC2} - \text{EC1}}{\text{EC3}} \right] \times 100$$
By measuring the lipid peroxidation by-product malonyldialdehyde, the membrane stability index was examined. It was determined using a fresh weight basis and represented in terms of nmol basis using the thiobarbituric acid reactive substance (TBARS) method as per [18]. The MDA content was calculated by using the equation mentioned below:
$$\text{MDA} \, \text{g}^{-1} \text{FW}(\text{nmol}) = 6.45(\text{OD532} - \text{OD600}) - (\text{OD450}) \times 1000$$
(4)
Determination of Activities of Antioxidant Enzymes
The antioxidant enzyme activity in the 45-day-old leaves of all treated $V. \text{mungo}$ plants was measured. Fresh leaves (100 mg) from all treatments were homogenized in 200 L of extraction buffer (10 % glycerol, 62.5 mM Tris-HCl, pH 7.5, 0.05% -mercaptoethanol, and 1 mM EDTA) to extract the total crude protein. Protein was then quantified using the Bradford method [19] and a protein dye reagent (Bio-Rad). The extracted protein was utilized in assays for antioxidant enzymes. The reduction in $A_{290}$ caused by ascorbate consumption in 1 mL of the assay mixture (10 mM $H_2O_2$ and 0.5 mM ascorbate) with an extinction coefficient at 290 nm of 2800 M$^{-1}$ Cm$^{-1}$ owing to 50 µl of the crude protein extract in buffer was used to determine the activity of the APX enzyme [20]. One unit of APX activity was defined as the quantity of enzyme needed to decrease 1.0 mol of $H_2O_2$ in one minute under the test conditions, and the enzyme activity was expressed as a unit mgL$^{-1}$ of protein. The method proposed was used to measure SOD activity by inhibiting the photochemical reduction of nitro blue tetrazolium (NBT) spectrophotometrically at 560 nm [21]. The experiment was conducted in 5.0 mL of a reaction mixture containing 100 mL of enzyme extract, 13 mM methionine, 63 mM NBT, 1.3 mM riboflavin, and 50 mM phosphate buffer (pH 7.8). The GR’s activity was assessed by using the method described by Smith IK, et al. [22]. The assay mixture (5 mL) contains 50 µl of enzyme extract at 25 °C, 1 mM oxidized glutathione, and 2 mM NADPH. Monitoring the decline in absorbance at 340 nm allowed researchers to analyze the oxidation of NADPH in more detail. The simultaneous CAT activity was assessed by the $H_2O_2$ intake in accordance with [23]. 100 µl of the protein extract, 50 mM potassium phosphate buffer (pH 7.0), 15 mM $H_2O_2$, and 2.5 mL of the reaction mixture were used to assay the CAT activity. The molar extinction coefficient of $H_2O_2$ at 240 nm, which was measured to be 36 M L$^{-1}$Cm$^{-1}$, was used to estimate the CAT activity. The CAT activity was displayed as 1 mg of protein per CAT unit minute.
**Measurement of Transpiration Rate and Photochemical Yield (Fv/Fm)**
The seventh leaf from the bottom of 45-day-old plants of all treatments of $V. \text{mungo}$ was selected to measure transpiration rate and photochemical yield. An open gas exchange system with a fluorescence chamber head (Li-Cor 6400XT; Li-Cor Inc., Lincoln, NE, USA) was used to measure the stomatal conductance and maximal photochemical efficiency of PS-II.
**Measurement of the Concentrations of $Na^+$ And $K^+$**
The measurement of $Na^+$ and $K^+$ concentrations was done in the leaf tissue of 45-day-old plants of all treatments of $V. \text{mungo}$ by the method described by Shekhar S [24]. In 2.0 mL vessels, 100 mg of dried material was weighed. Then, 1.0 mg of ultrapure water (Fluka Analytical, Sigma-Aldrich, USA) was added, and the tubes were heated for 60 minutes at 80°C. The boiled sample was centrifuged at 3000 g for 3 min to filter it through a 96-well filter plate from Thermo Scientific in Rochester, New York, USA. 5.0 l of the filtrate was diluted with 5.0 mL of ultrapure water (Fluka Analytical, Sigma-Aldrich) to determine the concentrations of $Na^+$ and $K^+$. An atomic absorption spectrometer (AAS) (Analyst 200; PerkinElmer) was used to detect the amounts of $Na^+$ and $K^+$. Sodium and potassium atomic spectroscopy standard concentration (Fluka Analytic, Sigma-Aldrich) was used to calibrate the AAS.
**Experiment Design and Statistical Analysis**
All the treatments were carried out in three replicates and each replicate possessed five plants. The data were subjected to analysis of variance (ANOVA). Mean differences were compared by the least significant difference (LSD) test using SPSS 22.0. Differences at p ≤0.05 were considered significant.
**Results and Discussion**
The germination percentage progressively diminished with increasing salinity and drought induced by NaCl and Mannitol respectively as compared to the unstressed control seeds. Under unstressed control conditions, 91.0% of seeds germinated after 5 days of sowing. However, the gradual increase of NaCl and Mannitol concentrations significantly reduced the germination percentage and number of days to sprouting of $V. \text{mungo}$ seeds (Figures 1a & 1b). The lowest percentage of germination was recorded as 34 % and 23 % in the seeds treated with 300 mM NaCl and 300 mM Mannitol respectively (Figures 1a & 1b). Similarly, the days to sprouting at the same concentration of NaCl and Mannitol were recorded as 16 days and 20 days respectively (Figures 1a & 1b). Our results illustrated the reduction of seed germination percentage and number of days to seed sprouting with gradual increment in the NaCl and Mannitol concentration from 100mM to 300mM. Due to osmotic stress and ion-specific effects, salt and drought stress had an impact on the seed germination process, which reduced the germination rate with prolonged germination time [25]. These two stressors enhanced external osmotic potential and caused a reduction in water uptake during imbibitions. Moreover, salinity stress also affects the seed germination due to the toxicity of excessive Na+ and Cl- ions which ultimately affects embryo viability [26, 27]. These toxic effects include disruption of protein structure, respiration, photosynthesis, protein synthesis and damage to cell organelles [28]. The seed germination follows three steps under normal conditions; in the first step the seed germination begins with the imbibitions followed by a plateau step and the third step is reactivation of cellular metabolisms. Based on these three steps of seed germination, the reduction in germination frequency and delaying of sprouting time under salinity and drought stress might be ascribed to osmotic stress during imbibitions, and plateau steps. The osmotic stress and ionic stress interact together and inhibit/ delay the seed germination processes during the step of cellular metabolism [29]. In the present investigation, the reduction of germination percentage and delayed seed sprouting might be associated with the disruption of ionic homeostasis and osmolarity which disrupts other metabolic pathways.
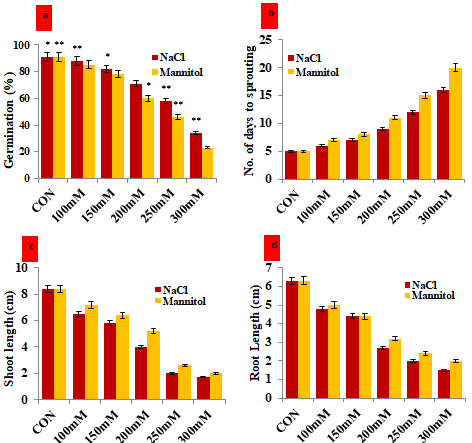
Figure 1: Assessment of the impact of magnetic hematite nanoparticle on in-vitro growth and development attributes of Vigna mungo under different concentration of NaCl and Mannitol stress. a) Germination percentage of V. mungo seeds soaked in 100-300 mM NaCl and Mannitol solution for 24 h; b) Number of days to sprouting of V. mungo seeds; c) Shoot length (cm) of V. mungo seeds sown in petri plates; d) Root length (cm) of V. mungo seeds sown in petri plates. (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non- treated control plant exposed to the same treatment).
Moreover, to investigate the effect of hematite nanoparticles on the vegetative growth of V. mungo seeds grown under in-vitro imposed stress conditions, we measured the length of radical and plumule. It was recorded that the gradual increment in the NaCl and Mannitol concentration influenced the length of plumule and radical (Figures 1c & 1d). The non-treated control seeds of V. mungo possessed the highest length of plumule and radical as compared to the NaCl and Mannitol-treated seedlings. Simultaneously, a negative correlation with an inverse linear relationship was evident between imposed stress level (salt and drought) and the length of plumule and radical. Under the non-stressed control condition, the highest length of plumule and radical was measured as 8.4 cm and 6.3 cm respectively (Figures 1c & 1d). However, the minimum length of plumule was measured in the seeds treated with 300 mM of NaCl (1.7)
and 300 mM Mannitol (2.0 cm). Similarly, the length of the radical also follows the same pattern as the plumule. The minimum length of the radical was measured as 1.5 cm under 300 mM of NaCl, however, under 300 mM of Mannitol the radical length was measured as 2.0 cm (Figures 1c & 1d). The reason might be due to the negative effect of NaCl and Mannitol on the activity of phytohormones, growth regulators which are the important players of cell division and elongation. Deferral in the differentiation of plumule and radical and their respective elongation under saline and drought conditions might be associated with disturbance of the osmotic potential of the cell. Higher concentration of salt restricts the water intake by seeds during seed germination by obstructing film and it leads to the inactivation of cytosolic catalysts and plant growth regulators, which are important player in cell differentiation, elongation and division. El Baz FK, et al. [30, 31, 32], reported that the presence of NaCl even at a low concentration enters into the plant and break the osmotic potential of sprouting structures, which ultimately reduce/inhibit the elongation of plumule and radical.
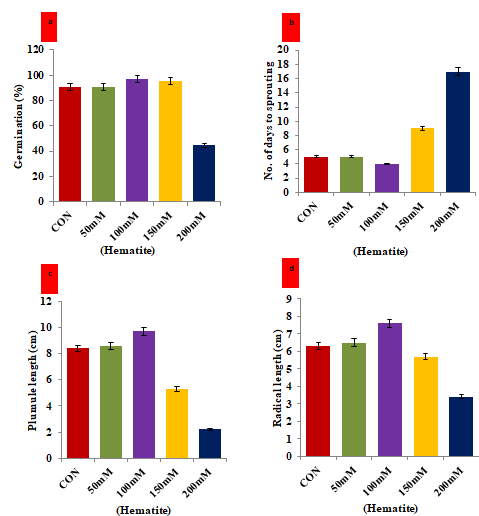
Figure 2: Hormesis effect of magnetic hematite nanoparticle on the attributes of V. mungo seeds a) Histogram showed the germination percentage of V. mungo seeds soaked in hematite nanoparticle (50mM to 200mM) for 1 h; b) Number of days to sprouting of V. mungo seeds; c) Shoot length (cm) of V. mungo seeds sown in petri plates; d) Root length (cm) of V. mungo seeds sown in petri plates. (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non-treated control plant exposed to the same treatment).
The seeds of V. mungo were treated with four different concentrations (50, 100, 150, and 200µM) of hematite nanoparticles to determine their impact on the seed germination characteristics. Our results given in Figures 2a, 2d showed a differential impact of treatment under in- vitro conditions. The highest percentage of germination was evident under treatment of 100 µM hematite treatment followed by 150 µM concentration of hematite. Moreover, the treatment of 200µM hematite exerted a deleterious impact on germination percentage as compared to other treatments and untreated control seeds (Figure 2a). The number of days to sprouting was lowest in the seeds treated with 100 µM of hematite nanoparticles, and it was recorded as 4 days. However, the treatment with 200 µM of hematite enhanced the days to sprouting and it was recorded as 17 days. Simultaneously, we measured the length of plumule and radical under different treatments of hematite. Our results emphasized that the longest plumule and radical was evident under the treatment of 100 µM of hematite nanoparticle among all the four treatments and untreated control seeds. The outcome of the hematite treatment revealed that 100 µM concentrations was best suited and optimized to enhance the germination percentage by 12.0%, plumule length by 15.4% and radical length by 20.6% as compared to untreated stressed control seedlings. Iron is one of the important elements required for plant growth, development and reproduction via regulation of different physiological and metabolic processes in the plants. The black gram seedlings grown under different salt and drought stress differ in morphology in terms of plumule and radical length. Schiefelbein, et al. [33] reported that roots increases the area for the absorption of mineral uptake and the root hairs polarized towards the outgrowth of root epidermal cells, which lead to the elongation of radical and plumule. The elongation of plumule and radical is also regulated by the plant hormones as well as environmental factors [34].
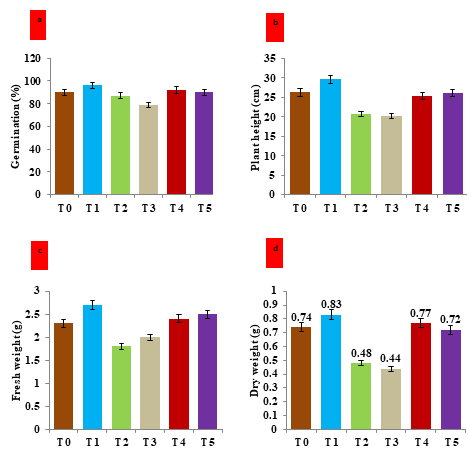
Figure 3: Effect of hematite nanoparticles on the morphological attributes of V. mungo plants grown under salt and drought stress under in-vivo condition. a) Histogram showing the germination percentage of V. mungo seeds under different treatment; b) Histogram showing the plant height of V. mungo grown under different treatments; c) Histogram showing the Fresh weight (g) of V. mungo plants grown under in-vivo condition; d) Histogram showing the dry weight (g) of V. mungo plants grown under in-vivo condition. T0: Control (non-stressed, non-treated) plants of V. mungo; T1: Non-stressed plants, of V. mungo treated with hematite nanoparticle; T2: Salt (NaCl) stressed plants of V. mungo; T3: Drought (Mannitol) stressed plants of V. mungo; T4: Salt (NaCl) stressed plants of V. mungo treated with hematite nanoparticle; T5: Drought (Mannitol) stressed plants of V. mungo treated with hematite nanoparticle (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non-treated control plant exposed to the same treatment).
The bioavailability of Fe in nano form might show efficient sequestration as compared with the ionic forms of Fe available in rhizosphere. Hence, magnetic hematite causes an enhancement in the length of plumule and radical. Results revealed that application of hematite nanoparticles enhances the vegetative growth attributes under imposed stress such as; germination percentage, plant height, fresh weight and dry weight of V. mungo plants grown under in-vivo conditions. When treated with 100mM of hematite nanoparticles, the germination percentage of seedlings cultivated under salt and drought stress was increased by 5.7% and 7.7%, respectively, in comparison to the corresponding non-treated stressed counterparts. The results shown in Figure 3b emphasized that the treatment of hematite positively regulates the plant height. In the comparison of mean plant height, it was evident that treatment of hematite nanoparticles increases the plant height by 22.2% and 28.7% under salt and drought stress respectively in comparison to their corresponding counterparts which were grown under untreated stressed conditions (Figure 3b). The fresh weight and dry weight possessed a positive correlation with plant height (Figures 3c & 3d). The percentage increase of fresh weight was highest in the plants grown from seeds treated with 100 mM of hematite nanoparticles and sown under salt and drought stress (Figure 3c). The fresh weight of hematite-treated plants was recorded as 33.3% higher than the untreated salt- stressed (T2) plants. Simultaneously, the percentage increase of fresh weight was recorded as 25.0 % in nanoparticle- treated and stressed plants as compared to non-treated salt-stressed counterparts (Figure 3c). Moreover, the dry weight also follows the same pattern as fresh weight. The highest dry weight was recorded in the plants grown from the seeds treated with 100mM hematite nanoparticles and grown under imposed stress of salt and drought (Figure 3d). The percentage increment in dry weight was recorded as 60.4% and 63.3% in the plants grown under salt and drought stress respectively after treatment with 100mM of hematite nanoparticle as compared to their corresponding non- treated and stressed counterparts (Figure 3d). In line to these findings, [35] assessed the impact of iron nanoparticles on muskmelon and showed that a positive correlation between NPs treatment and plant growth attributes. Another study conducted by Tombuloglu H, et al. [36] reported that the use of iron nanoparticles enhanced the fresh and dry weight of barley plants. Our results also showed the synonymous findings [37, 38].
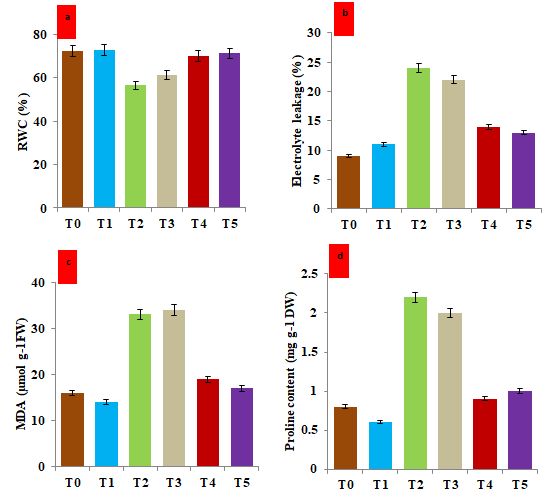
Figure 4: Effect of hematite nanoparticle on the biochemical stress markers in the leaf tissue of V. mungo grown under salt and drought stress under in-vivo condition after 30 days. a) Histogram showing the percentage of relative water content (RWC) under different treatments; b) Histogram showing the electrolyte leakage under different treatments; c) Histogram showing the malonyldialdehyde (MDA) content; d) Histogram showing the proline content. T0: Control (non-stressed, non-treated) plants of V. mungo; T1: Non-stressed plants, of V. mungo treated with hematite nanoparticle; T2: Salt (NaCl) stressed plants of V. mungo; T3: Drought (Mannitol) stressed plants of V. mungo; T4: Salt (NaCl) stressed plants of V. mungo treated with hematite nanoparticle; T5: Drought (Mannitol) stressed plants of V. mungo treated with hematite nanoparticle. (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non-treated control plant exposed to the same treatment).
Biochemical responses of salt and drought stress and treatment of hematite nanoparticles under salt and drought stress were investigated. We measured; relative water content (RWC), Electrolyte leakage (EL), malonylaldehyde (MDA), and proline. Our results emphasized that salinity and drought stress significantly lowered the RWC (Figure 4a). Under salt and drought stress a reduction of 22.0% and 15.8% in RWC was recorded as compared to the unstressed untreated control plants. However, treatment of hematite increased the RWC by 23.9% and 16.3% under salt and drought stress conditions, respectively. In the present study, EL, MDA, and proline content were increased due to salt and drought stress. The comparison in the electrolyte leakage (EL) between stressed and unstressed plants showed that EL increased by 166.2% due to salt stress and 144.4% due to drought stress (Figure 4b). The seed treatment of hematite nanoparticle reduced the EL by 78.6% and 82.6% under imposed stress of salt and drought stress, respectively (Figure 4b). A significant effect of salt and drought stress on the MDA content was recorded. The level of MDA in untreated and unstressed control plants was recorded as 160.65 µmolg-1 of fresh weight in V. mungo plants. However, under salt and drought treatment, the MDA content increased by 106.0 % and 112.5%, respectively. The treatment of hematite nanoparticles significantly reduced the MDA content by 42.4% and 50.0% in the salt and drought-stressed plants (Figure 4c). Similarly, the incremental percentage of the proline content in salt and drought stress was recorded as 175% and 150%, respectively. Moreover, the treatment of hematite nanoparticles has reduced the proline by 59.0% and 50.0% in salt and drought-treated plants, respectively (Figure 4d). Some previous studies reported the role of iron NP in the mitigation of the deleterious effects of abiotic stress in plants [39, 40, 41]. Fe NPs induce protection in Vitis vinifera under drought stress through the reduction in MDA with enhancement of non-enzymatic antioxidants and the activity of enzymatic antioxidants [42]. Lipid peroxidation of the cell membrane is one of the crucial metabolic processes which defines the extent of cell damage and ionic leakage in plants during stress. Under stress conditions, the selective permeability of the membrane is distorted due to lipid peroxidation which allows higher permeability and electrolyte leakage [43]. The results of the present study are synonymous with the findings of aforesaid scientific facts that treatment of hematite nanoparticles resulted in the lowering of MDA, EL and proline content and enhanced the RWC which led to enhanced the stress tolerance potential of the V. mungo plants.
Imposition of salt and drought stress significantly affects total chlorophyll content, Fv/Fm, and stomatal conductance (Figure 5a, 5c). In the present study, we found that salt and drought stress reduced the chlorophyll content by 54.27% and 58.8% respectively. However, the treatment of hematite nanoparticles under salt and drought stress increased the chlorophyll content by 78.7% and 82.4%, respectively as compared to their stressed counterparts (Figure 5a). Our results revealed that the maximum quantum yield of PSII (Fv/Fm) under salt and drought stress decreased in the plants of V. mungo (Figure 5b). Under salt stress Fv/Fm was reduced by 23.3%, however under drought stress the Fv/Fm was reduced by 28.5%. However, the treatment with hematite nanoparticles significantly enhances the photochemical yield of PSII. The treatment of hematite enhanced the Fv/Fm by 13.5% and 14.5% under imposed stress of salt and drought stress in contrast to their stressed counterparts (Figure 5b). The stomatal conductance was also decreased under salinity and drought stress; however treatment of hematite nano- particle under salt and drought stress significantly minimize the reduction in the stomatal conductance as compared to stressed plants. Under salt and drought stress the stomatal conductance decreased by 33.2% and 36.6%, respectively (Figure 5c). However, the percentage of increase in stomatal conductance by 63.0% and 61.0% due to the treatment of hematite nanoparticles under imposed stress of salt and drought respectively (Figure 5c).
The Na+ and K+ contents and K+/Na+ ratio in the leaves under salt and drought stress were measured to examine the effect of the hematite NPs on the plant tolerance potential (Figures 5d & 5f). The Na+ contents in the leaves of unstressed untreated control plants of V. mungo were measured as 3.7 mg g-1 dry weight. However, under imposed stress of salt and drought, the Na+ content was increased by 213.5% and 232.4%, respectively as compared to control plants. However the K+ content was measured as 19.3 mg g-1 dry weight in unstressed untreated control plants of V. mungo (Figure 5e). The K+ content decreased by 38.8% and 35.7% under the salt and drought stresses, respectively. In comparison to control counterparts, treated hematite NPs reduced Na+ by 42.3% and 44.8% under salt and drought stress, respectively. However, K+ concentrations increased by 33.8% and 32.2% under the same conditions. Under normal conditions, the K+/Na+ ratio was 5.2; however, when salt and drought stress were applied, the ratio decreased by 80.4% and 82.7%, respectively. Whereas the hematite treatment enhanced the ratio of K+/Na+ by 213.7% and 190.0% under imposed salt and drought stress (Figure 5f). As a measure of the photosynthetic energy conversion in plants, the Fv/Fm ratio captures the light reemission by chlorophyll molecules when light returns from the 350 nm excited state to the ground state [44]. In our study treatment with hematite NP enhanced the ratio of Fv/Fm and it could be corroborated with the maximum conversion of photosynthetic energy in V. Mungo plants under salt and drought stress. Sharma, et al. [45] also reported that the treatment of iron nanoparticles enhances the stress tolerance potential of Faba bean against salinity stress through the maintenance of ionic homeostasis.
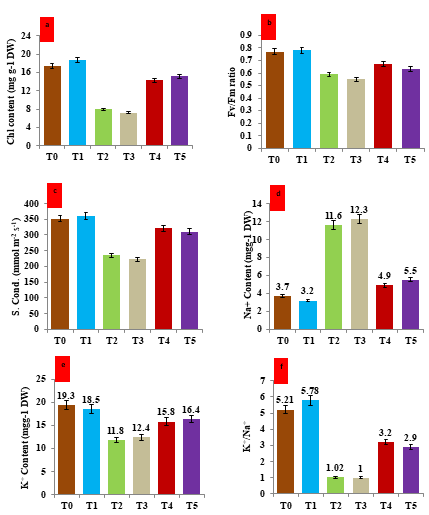
Figure 5: Impact of hematite nanoparticle treatment on the physiological attributes of V. mungo leaves grown under salt and drought stress- a) Total chlorophyll content (mg) of per gram dry weight of the leaf tissue of V. mungo; b) Histogram showing the photosynthetic rate of photosystem II under the influence of hematite treatment; c) Histogram showing the stomatal conductance of V. mungo plants grown under salt and drought stress; d) Sodium (Na+) content in the V. mungo plants grown under salt and drought stress; e) Pottasium (K+) content in the V. mungo plants grown under salt and drought stress; f) Ratio of potassium/sodium (K+/Na+) in the V. mungo plants grown under salt and drought stress. T0: Control (non-stressesd, non- treated) plants of V. mungo; T1: Non-stressed plants, of V. mungo treated with hematite nanoparticle; T2: Salt (NaCl) stressed plants of V. mungo; T3: Drought (Mannitol) stressed plants of V. mungo; T4: Salt (NaCl) stressed plants of V. mungo treated with hematite nanoparticle; T5: Drought (Mannitol) stressed plants of V. mungo treated with hematite nanoparticle. (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non-treated control plant exposed to the same treatment).
The analysis of antioxidant enzyme activity revealed that salt and drought stress significantly increase the activity of four antioxidant enzymes in the V. mungo plants. In comparison to their untreated, unstressed control counterparts, the activity of SOD, APX, CAT, and GR rose by 282.2%, 187.3%, 120.0%, and 441.6% under induced salt stress. Similar to SOD, APX, CAT, and GR, under imposed drought stress, activity rose sharply by 247.8%, 210.3%,
100.0%, and 424.2%, respectively, in comparison to matching control equivalents (Figures 6a-6d). The maximum increases in all four enzyme activities were observed in the hematite NP-treated plants of V. mungo in comparison with stressed plants. Under hematite treatment under salt stress, the increase was 80.0 %, 78.5 %, 55.5 % and 33.3 % in SOD, APX, CAT and GR activity, respectively as compared to the stressed plants. Hematite treatment raised the simultaneous activity of SOD, APX, CAT, DHAR, and GR enzymes under drought stress in Kufri Badshah by 71.6, 60.5, 79.8, 82.5, and 99%, respectively, compared to stressed plants. Chen, et al. [46], demonstrated under salt stress, iron NP treatment in grapes dramatically improved the activity of antioxidant enzymes. Our findings are consistent with those of Mozafari, et al. [44]. Application of iron NP reduced SOD and POX activity, while 1.5 mg/l iron NP increased CAT activity in a high-drought environment. PPO activity was triggered by two concentrations of iron NP at low levels of dryness. In our study, the CAT enzyme was more impacted by iron NP than the SOD enzyme. This is in line with our findings relating to H2O2 concentration, which showed a considerable decrease after being exposed to iron NP. Rezayian, et al. [47] according to reports, Fe plays a key role in plant metabolism by activating antioxidant enzymes. When iron is employed as nanoparticles, it can help V. mungo plants to rapidly absorb the needed quantity of iron under stressful circumstances. The antioxidant enzymes involved in detoxifying reactive oxygen species created in stressed plants by the NPs may function more effectively. The resilience of V. mungo plants to salt and drought stress associated with hematite NP may be connected to these changes in antioxidant enzyme activity. Our findings demonstrated a strong relationship between iron nanoparticles (NP) and antioxidant enzymes in plant cells.
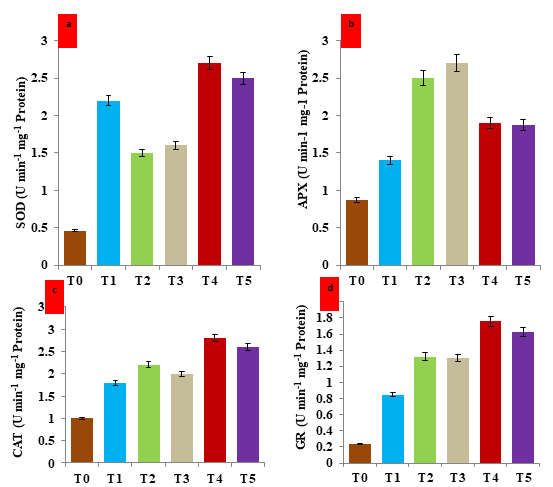
Figure 6: Effect of hematite nanoparticle on the antioxidant enzyme activity of protein extracted from the leaves of V. mungo plants grown under salt and drought stress-a) Histogram showing the enzyme activity of superoxide dismutase (SOD); b) Histogram showing the enzyme activity of ascorbate peroxidase (APX); c) Histogram showing the enzyme activity of catalase (CAT); d) Histogram showing the enzyme activity of glutathione reductase (GR). T0: Control (non-stressed, non-treated) plants of V. mungo; T1: Non-stressed plants, of V. mungo treated with hematite nanoparticle; T2: Salt (NaCl) stressed plants of V. mungo; T3: Drought (Mannitol) stressed plants of V. mungo; T4: Salt (NaCl) stressed plants of V. mungo treated with hematite nanoparticle; T5: Drought (Mannitol) stressed plants of V. mungo treated with hematite nanoparticle. (Data represent the mean of three replicates (n=3), with error bars representing standard deviation of the mean. (p≤ 0.05) between treated lines and non-treated control plant exposed to the same treatment).
Conclusion
This study showed that the magnetic hematite nanoparticle (NP) increased the potential of V. mungo salt and drought tolerance and offered a new method for increasing plant tolerance to abiotic stress. Our findings from this experiment demonstrated the effectiveness of magnetic hematite nanoparticles (NP) in preventing salt and drought-induced disturbance in the Halliwell-Asada Pathway of V. mungo plants by increasing ROS detoxification and ionic homeostasis maintenance. This work also highlighted the possible function of hematite nanoparticles in controlling photosynthetic behaviour by maintaining increased chlorophyll content, stomatal conductance, and PSII’s maximal photochemical efficiency, which led to the restoration of cellular homeostasis under salt and drought stress. However, our findings indicate that hematite NPs play a crucial role in a broad spectrum of functions.
References
-
Esmaeili N, Cai Y, Tang F, Zhu X, Smith J, et al. (2021) Towards doubling fibre yield for cotton in the semiarid agricultural area by increasing tolerance to drought, heat and salinity simultaneously. Plant Biotechnology Journal 19(3): 462-476.
-
Munns R, Tester M (2008) Mechanisms of Salinity Tolerance. Annual Review of Plant Biology 59: 651-681.
-
Seleiman MF, Al Suhaibani N, Ali N, Akmal M, Alotaibi M, et al. (2021) Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants (Basel) 10(2): 259.
-
Jhanji S, Dhingra M (2020) Ameliorative effect of thiourea priming on germination characteristics of mungbean (Vigna radiata L.) under water and salinity stress. Legume Research 43(3): 353-358.
-
Allahmoradi PM, Ghobadi S, Taherabadiand S (2011) Physiological Aspects of Mungbean in Response to Drought Stress. Food Eng Biotechnology IACSIT Press, Singapoore.
-
dos Santos TB, Ribas AF, de Souza SGH, Budzinski IGF, Domingues DS (2022) Physiological Responses to Drought IGF, Salinity, and Heat Stress in Plants: A Review. Stresses 2(1): 113-135.
-
Mariyappillai A Kulanthaivel V (2022) Effect of Nacl Salt Stress on Germination and Seedling Growth of Black Gram (Vigna mungo). Indian Journal of Agricultural Research.
-
Saha P, Chatterjee P, Biswas AK (2010) NaCl pretreatment alleviates salt stress by enhancement of antioxidant defensesystem and osmolyte accumulation in mungbean (Vigna radiata L. Wilczek). Indian Journal of Experimental Biology 48(6): 593-600.
-
Kumera N, Ayman N, Siraj U, Yetenayet B (2021) Application of nanotechnology in agriculture, postharvest loss reduction and food processing: food security implication and challenges. Heliyon 7(12): e08539.
-
Hochmuth G (2011) Iron (Fe) nutrition of plants. SL353/ SS555, 8/2011. EDIS 2011(8).
-
Morrissey J, Guerinot ML (2009) Iron uptake and transport in plants: the good, the bad, and the ionome. Chem Rev 109(10): 4553-4567.
-
Babu S, Singh R, Yadav D, Rathore SS, Raj R, et al. (2022) Nanofertilizers for agricultural and environmental sustainability. Chemosphere 292: 133451.
-
Bisht S, Sharma V, Kumari N (2022) Biosynthesized hematite nanoparticles mitigate drought stress by regulating nitrogen metabolism and biological nitrogen fixation in Trigonella foenum-graecum. Plant Stress 6: 100112.
-
Schwertmann U, Cornell RM (2000) Iron Oxides in Laboratory: Preparation and Characterization, second ed. WILEY-VCH, Weinheim, Germany.
-
Garcia Mata C, Lamattina L (2003) Abscisic acid, nitric oxide and stomatal closure-is nitrate reductase one of the missing links. Trends in Plant Science 8(1): 20-26.
-
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water stress studies. Plant Soil 39: 205-207.
-
Sairam RK, Srivastava GC (2002) Changes in antioxidant activity in subcellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Science 162: 897-904.
-
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125(1): 189-198.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
-
Hiner AN, Rodrıguez Lopez JN, Arnao MB, Lloyd Raven E, Garcıa Canovas F, et al. (2000) Kinetic study of the inactivation of ascorbate peroxidase by hydrogen peroxide. Biochem J 348(2): 321-328.
-
Beyer WF, Fridovich I (1987) Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem 161(2): 559-566.
-
Smith IK, Vierheller TL, Thorne CA (1989) Properties and functions of glutathione reductase in plants. Physiol Plant 77(3): 449-456.
-
Aebi H (1984) Catalase in vitro. Methods Enzymol 105: 121-126.
-
Shekhar S, Mahajan A, Pandey P, Raina M, Rustagi A, et al. (2023) Salicylic Acid and Methyl Jasmonate Synergistically Ameliorate Salinity Induced Damage by Maintaining Redox Balance and Stomatal Movement in Potato. Journal of Plant Growth Regulation 42: 4652- 4672.
-
Munns R (2002) Comparative physiology of salt and water stress. Plant Cell and Environment 25(2): 239- 250.
-
Jahromi F, Aroca R, Porcel R, Ruiz Lozano JM (2008) Influence of salinity on the in vitro development of Glomus intraradices and on the in vivo physiological and molecular responses of mycorrhizal lettuce plants. Microbial Ecology 55(1): 45-53.
-
Daszkowska Golec A (2011) Arabidopsis seed germination under abiotic stress as a concert of action of phytohormones. OMICS 15(11): 763-774.
-
Uçarlı C (2020) Effects of Salinity on Seed Germination and Early Seedling Stage. In: Shah Fahad, Shah Saud, et al. (Eds.), Abiotic Stress in Plants.
-
de la Reguera E, Veatch J, Gedan K, Tully KL (2020) The effects of saltwater intrusion on germination success of standard and alternative crops. Environmental and Experimental Botany 180: 104254.
-
El Baz FK, Mohamad AA, Aly AA (2003) Development of biochemical markers for salt stress tolerance in cucumber plants. Pakistan Journal of Biological Sciences 6(1): 16-22.
-
Horoun SA (2002) Fenugreek growth and metabolism in response to gibberallic acid and sea water. Bulletin of the Faculty of Science Assiut University 31: 11-21.
-
Huang WL, Liu FL (2002) Carbohydrate metabolism in rice during callus induction and shoot regeneration induced by osmotic stress. Botanical Bulletin of Academia Sinica 43: 107-113.
-
Schiefelbein JW (2000) Constructing a plant cell. The genetic control of root hair development. Plant Physiology 124 (4): 1525-1531.
-
Schmidt W, Schikora A (2001) Different pathways are involved in phosphate and iron stress‐induced alterations of root epidermal cell development. Plant Physiology 125(4): 2078-2084.
-
Wang Y, Wang S, Xu M, Xiao L, Dai Z, et al. (2019) The impacts of γ-Fe2O3 and Fe3O4 nanoparticles on the physiology and fruit quality of muskmelon (Cucumis melo) plants. Environmental Pollution 249: 1011-1018.
-
Tombuloglu H, Albenayyan N, Slimani Y, Akhtar S, Tombuloglu G, et al. (2022) Fate and impact of maghemite (γ-Fe2O3) and magnetite (Fe3O4) nanoparticles in barley (Hordeum vulgare L.). Environ Sci Pollut Res Int 29(3): 4710-4721.
-
Adrees M, Khan ZS, Ali S, Hafeez M, Khalid S, et al. (2020) Simultaneous mitigation of cadmium and drought stress in wheat by soil application of iron nanoparticles. Chemosphere 238: 124681.
-
Dola DB, Mannan MdA, Sarker U, Al Mamun MdA, Islam T, et al. (2022) Nano-iron oxide accelerates growth, yield, and quality of Glycine max seed in water deficits. Frontier in Plant Science 13: 992535.
-
Waqas M, Ishtiaq M, Maqbool M, Akram R, Shahid A, et al. (2022) Seed priming with iron oxide nanoparticles raises biomass production and agronomic profile of water-stressed flax plants. Agronomy 12(5): 982.
-
Bidabadi SS, Sabbatini P, Vander Weide J (2023) Iron oxide (Fe2O3) nanoparticles alleviate PEG-simulated drought stress in grape (Vitis vinifera L.) plants by regulating leaf antioxidants. Scientia Horticulture 312(15): 111847.
-
Saed Moucheshi A, Hasheminasab H, Khaledian Z, Pessarakli M (2015) Exploring morpho-physiological relationships among drought resistance related traits in wheat genotypes using multivariate techniques. J Plant Nutr 38: 2077-2095.
-
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. Journal of Experimental Botany 51(345): 659-668.
-
Torabian S, Zahedi M, Khoshgoftar AH (2017) Effects of Foliar Spray of Nanoparticles of FeSO4 on the Growth and Ion Content of Sunflower under Saline Condition. Journal of Plant Nutrition 40: 615-623.
-
Mozafari A, Ghadakchi A, Ghaderi N (2018) Grape response to salinity stress and role of iron nanoparticle and potassium silicate to mitigate salt induced damage under in vitro conditions. Physiol Mol Biol Plants 24(1): 25-35.
-
Sharma YK, León J, Raskin I, Davis KR (1996) Ozone- induced responses in Arabidopsis thaliana: The role of salicylic acid in the accumulation of defense-related transcripts and induced resistance. Proc Natl Acad Sci 93(10): 5099-5104.
-
Chen J, Liu X, Wang C, Yin SS, Li LX, et al. (2015) Nitric oxide ameliorates zinc oxide nanoparticles-induced phytotoxicity in rice seedlings. J Hazard Mater 297: 173- 182.
-
Rezayian M, Niknam V, Arabloo M (2023) Iron nanoparticle regulates succinate dehydrogenase activity in canola plants under drought stress. Sci Rep 13: 9628.
- Solution-Processed Chiral Perovskites for Biomedical Applications
- Nanotechnology in Health Chemistry and Medicine: Current Challenges and Future Directions
- Human Exposure to Micro- and Nanoplastics: Pathways, Toxicity, and Intervention Strategies
- Exosome Nanomedicine for Cancer Therapy
- Micro and Nanoplastics–Plastisphere, Biotoxicity, Impact on Human Health, and Mitigation Strategies
- Process Validation of Cefixime Powder for Suspension Dosage Form, 50 mL