3D Oral Mucosal Models for Studying Host-Pathogen Interactions in Periodontal Disease
The host immune response to the oral biofilm plays a central role in the aetiology of periodontitis. Our understanding of hostpathogen interactions has historically been reliant on monolayer culture systems stimulated by microorganisms. However, 2D culture systems possess major shortcomings as these do not adequately mimic the complexities of the native tissue and therefore fail to provide meaningful information on the pathogenesis of periodontal disease. To mimic the in vivo condition, various types of 3D oral mucosal equivalents (OME) have been employed, including reconstituted models and 3D organotypic culture models. The rationale for OME is to serve as a relevant in vitro tool to examine the interactions of human epithelial cells with bacterial biofilms, to understand the process of epithelial layer damage, molecular mechanisms underpinning the development of periodontitis and models for testing novel therapeutics. Furthermore, to gain a deeper understanding of periodontal dysbiosis, it is important to appreciate the molecular mechanisms underlying host epithelial cell production of cytokines and chemokines in response to bacterial stimulation; and signalling pathways which underpin host-pathogen interactions such as mitogen-activated protein kinases (MAPK) and nuclear factor kappa light chain enhancer of activated B cells (NFkB). Thus, the aim of this narrative review is to summarise the evidence regarding the application of 3D oral mucosal models in host-pathogen infection studies relating to periodontal disease, review the molecular mechanisms underlying host response and provide recommendations for future studies in this field.
Introduction
Physiologically relevant 3D mucosal models offer the ideal application for studying the molecular mechanisms of periodontal disease development and testing of novel therapeutics. The major goal of these oral mucosal equivalents (OME) is to be a valuable in vitro tool for studying (a) the interactions of human epithelial cells with bacterial biofilms, (b) the process of epithelial layer damage, (c) the molecular mechanisms underlying periodontitis development, and (d) models for testing novel therapeutics [1]. However, until late 1980’s, our understanding and studies of host-pathogen interactions was only limited to monolayer culture systems stimulated by microorganisms as they were able to allow the cells to receive a consistent supply of nutrients and growth factors [2]. However, these 2D (monolayer) culture systems were observed to have significant shortcomings: they do not adequately mimic the complexity of native tissue and thus fail to provide reliable information on morphological changes [2]. Importantly, they also lack the multilayering of epithelium, which, acts as a barrier against pathogen invasion, is responsible for the production of cytokines in response to oral microbiota and is involved in keratinocyte- fibroblast interactions [3]. Studies utilising 3D OMEs, including reconstituted models and 3D organotypic culture models, were being employed to mimic the in vivo condition [2, 4, 5]. These 3D models were considered physiologically relevant for studying the mechanisms of periodontal disease development, and testing novel therapeutics, even though most of the studies were restricted to <4h time frame due to the toxicity of microbes to epithelial cells [1]. However, as biofilm dysbiosis underlies the pathogenesis of periodontal disease [6] it is essential that an in vitro model be developed which would mimic the in vivo condition to the fullest extent, thus enabling the translational research to be carried out.
A variety of different 3D organotypic cultures have been suggested over the years that respond to infection with microbial pathogens in ways that reflect the natural infection process [7, 8, 9] and commercially or engineered models of the oral mucosa have also been developed for studying periodontal disease. We have also previously, proposed a plausible in vitro 3D organotypic culture model of the dentogingival junction which simulated the physiology of the tissues in vivo and respond to secreted biomolecules from the underlying fibroblasts [10, 11]. In all these models the standard procedure for engineering oral mucosa models involves: (a) isolation and expansion of epithelial and fibroblast cells from original tissues or commercial cell lines (b) seeding and culture of fibroblasts in a suitable substrate, and (c) finally, seeding of epithelial cells onto the engineered connective tissue layer [12, 13]. One of the major advantages of using tissue- engineered models in microbiology is the reduction of animal experiments [4] which reinforces the principles of the 3Rs, replacement, reduction, and refinement [14].
Inflammatory responses to oral microbiota differ depending on whether the epithelial cells are stimulated by planktonic or biofilms. These responses include altered cytokine levels, altered gene expression and changes in signalling pathways [3, 15]. While most infection studies have employed planktonic bacteria, biofilm dysbiosis is the basis of periodontal disease, as discussed earlier [6]. Therefore, this underlies the importance of employing multi-species biofilms and oral mucosal models in studying mechanisms of disease pathology. Currently, there is a lack of human physiologically relevant in vitro models for studying mechanisms of disease development. Therefore, the aim of this narrative review is to summarise the evidence regarding the application of 3D oral mucosal models in host-pathogen infection studies relating to periodontal disease, review the molecular mechanisms underpinning the host-pathogen interactions and provide recommendations for future studies in this field.
3D Organotypic Cultures
Epithelial-mesenchymal interactions may be more appropriately studied via the construction of 3D organotypic cultures. This technique allows the specific interaction between two populations to be studied in vitro under controlled conditions [4, 16, 17, 18, 19, 20]. The main advantage of employing the use of 3D cultures is that it is able to recapitulate the 3D organisation of cells and ECM within tissues and organs [2]. These culture systems allow for the in vitro growth of complex biological tissues in a way that models the 3D space which cells normally reside and function in vivo. Therefore, organotypic cultures systems models complex cellular interactions more effectively than monolayer cultures [21, 22, 23] by allowing complex interactions cell-cell and cell-ECM interactions to be modelled in such a way that replicates part of their normal physiology and function [2, 24].
The 3D culture models have proven to be more realistic when it comes to translating study findings for in vivo applications. The decrease in the gap between the cell culture system and the cellular physiology is a significant advantage of the 3D approach over the 2D approach [21, 25]. Some of the important areas for which 3D cell culture systems are excellent models include disease pathogenesis studies, drug discovery studies, drug response studies, and other applications such as pharmacological studies, understanding cell physiology, gene and protein expressions, cancer research, and tissue engineering [24, 26].
3D organotypic cultures used to model the oral mucosa typically consisted of epithelia supported by a collagen bed enriched with fibroblasts. Fibroblasts were typically embedded into collagen gel, which was allowed to contract before being seeded with keratinocytes and raised to the air- liquid interface for further growth and maturation [27].
Collagen matrix, composed of native collagen type I, is a commonly used hydrogel scaffold in 3D culture systems [28]. Type I collagen is the predominant structural protein with the ECM. The ease of handling, processing and relatively low cost are some of its many advantages. In addition, the structural properties of the collagen gel may be altered to suit the requirements of the experiment. For example, the pore size, ligand density and stiffness can be varied by changing the concentration of collagen or introducing chemical cross-linking compounds [29]. However, one drawback with collagen-based scaffolds is its poor biostability, with a relatively high biodegradation rate [5, 13] Cross-linked collagen with glutaraldehyde appears to provide an effective method to alter the biodegrading rate of collagen-based scaffolds without compromising on its biocompatibility [5, 12].
Reconstituted Epithelial Models
Advancements in tissue engineering, leading to the development of tissue-scaffold constructs to facilitate cell growth and differentiation, could be manipulated to express surface markers indicative of phenotypic traits, thereby producing immune-modulating molecules. Four commercially available tissue-engineered oral mucosa models are available [30]:
- Skin Ethic TM Human Oral Epithelium (HOE)
- Skin Ethic Human Gingival Epithelium (HGE) constructs by EPISKIN (Lyon, France)
- Epi Oral TM
- Epi Gingival TM tissues from MatTek Corporation (Ashland, MA) In the Skin Ethic models, epithelial cells are seeded onto inert filter substrates and raised to the air-liquid interface within a humidified incubator [30]. The models were cultured in a defined nutrient medium provided via the filter substratum. This model demonstrates the expression of natural major markers of epithelial differentiation and responds to pharmacologically active treatments similar to human epithelium in vivo [30]. Similarly, the Human Gingival Epithelium (HGE) model involves culturing human gingival cells on an inert polycarbonate filter at the air-liquid interface using a chemically defined medium [30]. In certain studies, the HGE model was co-cultured with immunological cells [31]. The HGE models feature proliferating basal cells (Ki- 67 positive) that extend to deeper layers, an advantageous characteristic for studying infections, as basal layer cell proliferation is typically restricted in normal mucosal cells [32]. The Epioral and EpiGingival models are cultivated in serum-free medium at the air-liquid interface and are equipped with a collagen matrix containing buccal and gingival fibroblasts [33, 34]. The epithelial tissue’s barrier integrity is assessed via the non-invasive, quantitative method of transepithelial electrical resistance (TEER) [35]. Notably, cornified EpiGingival tissue exhibits higher barrier lipid content and a more robust barrier compared to cornified EpiOral tissue. Both tissues manifest active cell division, as evidenced by Ki-67 staining in the basal layer, demonstrating similar Ki-67 expression to native tissue [32, 36]. MatTek has also introduced dendritic cells into the full thickness EpiOral tissue, while the functionality of dendritic cells within the tissue requires further investigation, the EpiOral model has proven useful in exploring disease progression and pathogenic mechanisms [36].
Bacterial Infection And Invasion Of Oral Mucosal Equivalents
Oral mucosal models could be infected by the addition of the desired concentration of microorganisms or bacterial biofilm (Figure 1). Most host- microbiome interaction studies generally employ the use of planktonic bacterial culture (non-biofilm design) and indeed our early understanding of pathogenesis of periodontal disease were based on monolayer cultures stimulated by planktonic bacterial culture [3].
However, oral microbiota tends to form multispecies biofilms, and their behaviour when embedded in a biofilm matrix is distinct from their planktonic form [37]. Indeed, an in vitro periodontal multi-species biofilm model developed to investigate the host modulatory effects of bioactive molecules on biofilm has demonstrated encouraging results which were found to be reproducible and stable over time [38]. Therefore, it was proposed that this particular in vitro periodontal biofilm model be applied for testing the efficacy of bioactive molecules for the treatment periodontitis prior to moving to clinical trials [38].
Long-term bacterial infection of the oral mucosa model is challenging due to their varying requirements for culture media. The time of infection of the oral mucosa with pathogenic microorganisms differs between studies, usually ranging from 1.5 to 48 hours, though in one study oral mucosal models were exposed to bacteria for 72 hours [1]. Prolonged incubation of oral mucosa model in anaerobic atmosphere compromises the structural integrity and epithelial cell viability after 24 hours [39]. Additionally, prolonged infection by periopathogens such as P. gingivalis and F. nucleatum reduces its viability as these organisms require anaerobic incubation to thrive [15].
The invasion of epithelium and connective tissue by periopathogens to elicit specific immune responses and the subsequent upregulation of signalling pathways are key events in the development of periodontitis [6]. Scratch assays performed to the epithelial layer to induce damage to the epithelial layer will provide a pathway for microbial invasion [1]. The penetration of F. nucleatum into the collagen matrix is strain- dependent and is more pronounced in the biofilm form than in planktonic form [37]. P. gingivalis is known to penetrate connective tissues [40] with the severity of its invasiveness being attributed to the presence of gingipains [41].
On the other hand, commensal oral bacteria seem to act as antagonists against potential pathogens and facilitate the maintenance of oral mucosal health [3]. A recent study showed that exposure to a biofilm of multispecies commensal microorganisms from healthy human saliva for 7 days increased epithelial thickness and keratinocyte proliferation in oral mucosa models [42] suggesting that commensal microbiota improved oral epithelial barrier function.
Lastly, the relative thickness of oral epithelium appears to impact on bacterial invasion. For example, submerged 3D organotypic constructs with thin epithelium allows penetration of P. gingivalis into the connective tissues, while airlifted constructs with thicker epithelium prohibits its penetration into the lamina propria. The viability of P. gingivalis in 3D constructs also decreases with time [15].
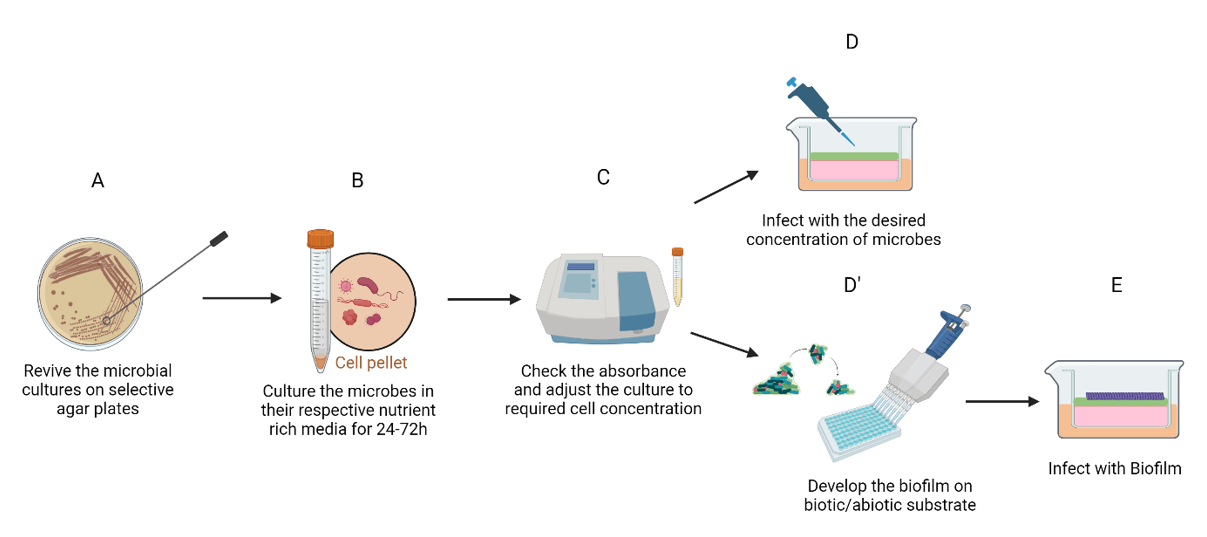
Figure 1: Protocol for bacterial infection of oral mucosal models. Following revival of microorganisms (A), they are cultured in respective nutrient rich media for 24-48 hours (B). Once the absorbance has been verified (C), the microorganisms could be directly infected on the mucosal model at the desired concentration (D). Alternatively, a biofilm could be developed on a substrate (D’) prior to infection (E).
Co-Infection Of Host Cells With P. Gingivalis And F. Nucleatum
In vitro epithelial cell co- cultures have been developed to model the complexities of periodontal bacteria–epithelium interactions, consisting of primary gingival epithelial cells, or epithelial cell lines either in monolayered or multilayered culture [31, 43]. Primary gingival epithelial cells, obtained from gingival explants, were known to express specific keratin and differentiation markers which characterise gingival epithelial cells [44, 45] . Transformed cell lines derived from gingival epithelial cells were known to be poorly differentiated and respond similarly to oral bacteria [46, 47].
Porphyromonas gingivalis, a Gram-negative anaerobic bacterium with black pigmentation, has been identified as a significant aetiological factor in the initiation and progression of periodontitis [16]. Fusobacterium nucleatum, an anaerobic Gram-negative bacterium, adheres to early colonisers and enhances the attachment of other periodontopathic bacteria such as P. gingivalis. Co-infection with F. nucleatum has been observed to heighten the ability of P. gingivalis to penetrate host cells [37]. However, the mechanisms underlying F. nucleatum’s interactions with P. gingivalis to influence the invasion of P. gingivalis into host cells remain unknown [48]. Brown, et al. have introduced an innovative model involving immune cells, gingival tissue, and biofilms to replicate cellular interactions at the gingival margin, while capturing the local inflammatory response [31].
The synergy between P. gingivalis and F. nucleatum in polymicrobial infections was observed to enhance the invasion of host cells by P. gingivalis, resulting in a notable increase in invasion efficiencies [49]. In particular, F. nucleatum, through its interactions with host cells, could synergistically enhance the invasion of P. gingivalis [50]. Additionally, P. gingivalis was found to hinder the expression of adhesion-related proteins (FadA and FomA) of F. nucleatum through proteases, potentially preventing F. nucleatum’s intracellular degradation and promoting proliferation and amplifying its overall infectivity [49]. Additionally, P. gingivalis and F. nucleatum were found to synergistically facilitate the invasiveness of other organisms. For example, P. gingivalis enhances the invasion of T. forsythia into epithelial cells [51] and F. nucleatum can transport non- invasive Streptococcus cristatus into epithelial cells through the formation of co-adhered dual species consortia [52]. These observations underscore the involvement of multi- species bacteria in the pathogenesis of periodontitis. In the next section we explore host epithelial cell responses to periopathogenic microorganisms.
Molecular Mechanisms Underpinning Pathogenic Invasion Of Host Cells
The oral epithelium, in collaboration with cells from the innate immune system, detects pathogen-associated molecular patterns (PAMPs) produced by microbes utilising a set of pattern recognition receptors (PRRs) encoded in germline cells [53]. There are, indeed, four principal classes of PRRs recognized: (1) Toll-like receptors (TLRs), (2) C-type lectin receptors (CLRs), (3) Nod-like receptors (NLRs), and (4) retinoic-acid-inducible gene 1 receptors (RIGs) [54]. The distribution of these PRRs varies based on cell type, with each type recognising specific PAMPs. Among these four classes, TLRs and NLRs predominantly participate in the identification of bacteria [55]. Activation of these PRRs triggers intracellular signalling pathways and specific transcription factors, ultimately resulting in the release of inflammatory mediators. The nuclear factor kappa light chain enhancer of activated B cells (NF-KB) and mitogen- activated protein kinases (MAPKs) signalling pathways play pivotal roles in regulating inflammation, immunological responses, as well as cellular growth, differentiation, and survival [56, 57].
The receptor activator of NF-KB (RANK) and its ligand RANKL, are both vital to osteoclast development and activation. RANKL, via the mitogen-activated protein kinase (MAPK) and the I kappaB kinase (IKK), triggers the activation of transcription factors AP-1 and NF-KB. The MAPK pathway comprises three arms: p38, c-Jun N-terminal kinase (JNK), and extracellular signal-regulated kinase 1/2 (ERK1/2). This signalling cascade engages downstream transcriptional events, including the activator protein-1 (AP-1) heterodimer [57]. Both the NF-KB and MAPK pathways significantly govern cytokine gene expression. These signalling pathways form an intricate network with overlapping processes that ultimately impact nuclear substrates responsible for regulating gene expression and protein synthesis [58].
TLR1 to TLR10 are recognized members within the TLR family. Notably, TLR1, 2, 4, 5, 6, and 9 participate in bacterial recognition [59]. Upon binding to ligands, the cytoplasmic Toll/IL-1Β receptor (TIR) domain of TLR interacts with various adaptor molecules, setting off a distinct MyD88- dependent pathway, primarily leading to the generation of inflammatory cytokines [60, 61]. Essentially, MyD88 engages with TNF receptor-associated factor 6 (TRAF6), which activates MAPK kinase (MAPKKK) TAK1. The latter instigates I kappa B (IKB) and NFkB, ultimately resulting in the production of pro-inflammatory cytokines. (Figure 2).
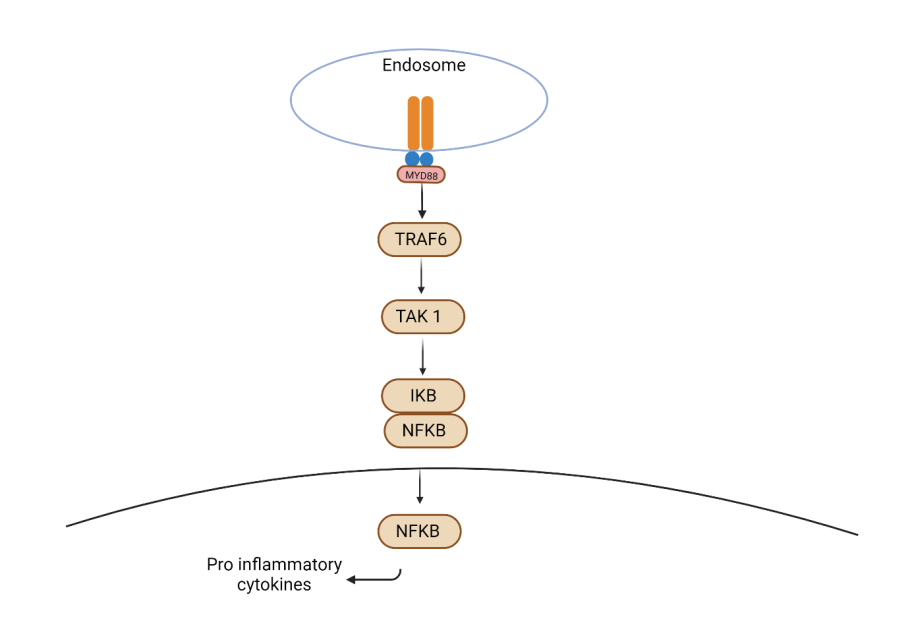
Figure 2: MyD88 dependent signalling pathway involved in bacterial recognition. MyD88 engages with TNF receptor- associated factor 6 (TRAF6), which activates MAPK kinase (MAPKKK) TAK1. The latter instigates I kappa B (IKB) and nuclear factor kappa light chain enhancer of activated B cells (NF-kB), ultimately resulting in the production of pro- inflammatory cytokines.
Upon invasion of host epithelial cells by pathogenic bacteria, several of the molecular signalling pathways are upregulated. For example, following the interaction of oral epithelial cells with either heat-killed or live F. nucleatum intensifies the activation of numerous signalling pathways, namely, triggering of the p38 and JNK MAP kinase pathways [62]. Upon exposure to live F. nucleatum, oral epithelial cells exhibit heightened expression of IL-8 mRNA, under the regulation of NF-KB, p38, and ERK1/2 activation. Specifically, live F. nucleatum triggers TLR and MAPK signalling pathways, leading to activation of IL-6 and IL-8 [61]. In a separate investigation focused on gingival epithelial cell multilayers, F. nucleatum stimulation was linked to the induction of IL-1β and TNF-⍺ [3]. Furthermore, investigations into F. nucleatum infections have illuminated the pivotal roles of NF-KB, p38, and JNK MAPK pathways in governing the production of IL- 1⍺, IL-1β, and IL-8 genes [62].
Upon exposure to heat-killed F. nucleatum, both TLR4 and TLR9 activation ensue, accompanied by the activation of NF-KB (p65 transcription factor). This prompts the induction of various factors including IL-1β, IL-8, GM-CSF, TNF-⍺, CCL2, CCL20, as well as S100A8 and S100A9-genes which are pivotal for cellular protection, neutrophil recruitment, and activation [63, 64].
Live P. gingivalis induces dose-dependent downregulation of ERK1/2, with minimal activation of the JNK and p38 MAPK pathways [65]. Conversely, P. gingivalis infections were associated with decreased ERK and IL-8 mRNA levels, confirming the role of the MAPK pathway [66]. In the presence of live P. gingivalis, IL-1β production is triggered, whereas IL-6 and IL-8 production in oral epithelial cells are inhibited. Strikingly, heat-killed P. gingivalis elicits a contrasting response, generating substantial amounts of IL-6 and IL-8 while not inducing IL-1β [67]. Notably, live, and heat-killed P. gingivalis elicit different reactions, with heat- killed P. gingivalis provoking a pro-inflammatory response, compared to the live strain, exhibiting a more subversive influence [68].
Heat-killed P. gingivalis activates TLR4, induces NF-KB activation, and stimulates the production of IL-1β, IL-8, GM- CSF, TNF-⍺, CCL2, and CCL20 in oral epithelial cells [69]. Studies indicate that P. gingivalis expresses pro-inflammatory factors, and its LPS has been found to induce the secretion of IL-1β, IL-6, IL-1Β0, TNF-⍺, IFN-, IP-10, and MIP-1 proteins in oral epithelial cells via TLR2 [69]. Moreover, P. gingivalis LPS activates the NF-KB signaling pathway, including the p65 transcription factor, along with the p38 and JNK MAPK pathways [70]. Recently, the haemoglobin receptor (HbR) protein of P. gingivalis was found to stimulate IL-8 secretion via p38 and ERK1/2 activation, subsequently activating ATF-2, CREB, and NF-KB transcription factors [71]. Despite the pro-inflammatory profile associated with heat-killed P. gingivalis and the anti-inflammatory profile of live P. gingivalis, investigations using the live strain have demonstrated both phenotypes, underscoring the bacterium’s capability to manipulate host responses [72].
Discussion
3D oral mucosal models are critical in gaining a comprehensive understanding of the molecular basis underlying periodontal dysbiosis. The combination of epithelial cells with fibroblasts in culture and the resultant molecular crosstalk between these two different cell types can be utilised in microbial infection of co- cultures to elicit clinically relevant host responses [1, 37, 38, 73]. The technological advances in the development of OME has the capacity to histologically examine the infectious process, visualise live epithelial tissue invasion and microorganism- induced damage, and analyse the in situ expression of specific fungal and host proteins or mRNA. This setup facilitates the exploration of interactions between a pathogen’s virulence attributes and host defence mechanisms.
The oral microbiota exists as complex multispecies biofilms. This is important since a multi-species biofilm can form a microbial community with metabolic benefits which can better withstand environmental stresses and higher resistance to antibacterial agents [37, 38, 74, 75].
Most of the bacterial infections applied to oral mucosal models have used planktonic bacterial cells instead of biofilm grown bacteria. Periodontal disease, however, is characterised by subgingival biofilm dysbiosis, inflammation and tissue destruction [6, 76]. Additionally, due to the complexity of the biofilm interactions that occur in a microbial community, biofilms behave very differently compared to their single species counterparts [74] such as the generation of distinct cytokine profile, signalling events and cellular differentiation [73]. Therefore, to effectively understand the mechanisms of host-pathogen interactions in periodontal disease, appropriate in vitro oral mucosal models, stimulated by oral multi-species biofilms are needed to replicate and reflect the intricacies of the in vivo host -biofilm interactions.
Although 3D mucosal models are more relevant and informative than epithelial cell monolayer cultures, there are still important limitations to be considered. For example, the models lack constant desquamation, an absence of saliva consisting of mucin, a deficiency in the number of present bacteria and immune responses and most of all, a static environment [3, 15, 38]. Most co-culture models (conventional submerged keratinocytes and fibroblasts) are limited to 48- 72 hours bacterial exposure, even though the interactions in vivo are in a lasting dynamic status [1, 15, 77, 78]. These limitations make it challenging to extrapolate data from in vitro experiments to the clinical situation. Lastly, the 3D oral mucosal models lack some of the other cell types present in normal human oral mucosa and therefore are unable to completely replicate the complexities of cellular interactions and its relationship with other anatomical structures in vivo [13]. Therefore, future studies employing vascularized OMM or those that contain immune cells would be indicated for studying host-pathogen interactions in periodontal disease.
In summary, 3D oral mucosal models offer the ideal application for studying the mechanisms of periodontal disease development and testing of novel therapeutics. Understanding the molecular mechanisms underlying host epithelial cell production of cytokines and chemokines in response to bacterial stimulation and invasion will be critical for the discovery of novel therapeutics for the management of periodontal disease. Thus, OMEs provides a robust in vitro methodology which would facilitate the exploration of interactions between a pathogen’s virulence attributes and host defence mechanisms.
References
-
De Ryck T, Grootaert C, Jaspaert L, Kerckhof FM, Van Gele M, et al. (2014) Development of an oral mucosa model to study host-microbiome interactions during wound healing. Appl Microbiol Biotechnol 98(15): 6831-6846.
-
Shamir ER, Ewald AJ (2014) Three-dimensional organotypic culture: Experimental models of mammalian biology and disease. Vol. 15, Nature Reviews Molecular Cell Biology. Nature Publishing Group pp: 647-664.
-
Dickinson BC, Moffatt CE, Hagerty D, Whitmore SE, Brown TA, et al. (2011) Interaction of oral bacteria with gingival epithelial cell multilayers. Mol Oral Microbiol 26(3): 210-220.
-
Elena Costea D, Lako Loro L, Anne Okumo Dimba E, Karsten Vintermyr O, Christine Johannessen A (2003) Crucial Effects of Fibroblasts and Keratinocyte Growth Factor on Morphogenesis of Reconstituted Human Oral Epithelium. J Invest Dermatol 121(6): 1479-1486.
-
Moharamzadeh K, Franklin KL, Brook IM, van Noort R (2009) Biologic Assessment of Antiseptic Mouthwashes Using a Three-Dimensional Human Oral Mucosal Model. J Periodontol 80(5): 769-775.
-
Hajishengallis G, Chavakis T, Lambris JD (2020) Current understanding of periodontal disease pathogenesis and targets for host-modulation therapy. Periodontology 2000 84(1): 14-34.
-
Carterson AJ, Höner Zu Bentrup K, Ott CM, Clarke MS, Pierson DL, et al. (2005) A549 lung epithelial cells grown as three-dimensional aggregates: Alternative tissue culture model for Pseudomonas aeruginosa pathogenesis. Infect Immun 73(2): 1129-1140.
-
LaMarca HL, Ott CM, Höner zu Bentrup K, LeBlanc CL, Pierson DL, et al. (2005) Three-dimensional growth of extravillous cytotrophoblasts promotes differentiation and invasion. Placenta 26(10): 709-720.
-
Nickerson CA, Goodwin TJ, Terlonge J, Ott CM, Buchanan KL, et al. (2001) Three-dimensional tissue assemblies: Novel models for the study of Salmonella enterica serovar typhimurium pathogenesis. Infect Immun 69(11): 7106-7120.
-
Lu EMC, Hobbs C, Dyer C, Ghuman M, Hughes FJ (2020) Differential regulation of epithelial growth by gingival and periodontal fibroblasts in vitro. J Periodontal Res 55(6): 859-867.
-
Lu EMC, Hobbs C, Ghuman M, Hughes FJ (2021) Development of an in vitro model of the dentogingival junction using 3D organotypic constructs. J Periodontal Res 56(1): 147-153.
-
Igarashi M, Irwin CR, Locke M, Mackenzie IC (2003) Construction of large area organotypical cultures of oral mucosa and skin. J Oral Pathol Med 32(7): 422-430.
-
Moharamzadeh K, Brook IM, Van Noort R, Scutt AM, Smith KG, et al. (2008) Development, optimization and characterization of a full-thickness tissue engineered human oral mucosal model for biological assessment of dental biomaterials. J Mater Sci Mater Med 19(4): 1793- 1801.
-
Graham ML, Prescott MJ (2015) The multifactorial role of the 3Rs in shifting the harm-benefit analysis in animal models of disease. Eur J Pharmacol 759: 19-29.
-
Pinnock A, Murdoch C, Moharamzadeh K, Whawell S, Douglas CWI (2014) Characterisation and optimisation of organotypic oral mucosal models to study Porphyromonas gingivalis invasion. Microbes Infect 16(4): 310-319.
-
Locke M, Hyland PL, Irwin CR, Mackenzie IC (2008) Modulation of gingival epithelial phenotypes by interactions with regionally defined populations of fibroblasts. J Periodontal Res 43(3): 279-289.
-
Chinnathambi S, Tomanek-Chalkley A, Ludwig N, King E, DeWaard R, et al. (2003) Recapitulation of oral mucosal tissues in long-term organotypic culture. Anatomical Record 270(2): 162-174.
-
Okazaki M, Yoshimura K, Suzuki Y, Harii K (2003) Effects of subepithelial fibroblasts on epithelial differentiation in human skin and oral mucosa: Heterotypically recombined organotypic culture model. Plast Reconstr Surg 112(3): 784-792.
-
Gibbs S, Ponec M (2000) Intrinsic regulation of differentiation markers in human epidermis, hard palate and buccal mucosa. Arch Oral Biol 45(2): 149-158.
-
Kautsky MB, Fleckman P, Dale BA (1995) Retinoic acid regulates oral epithelium differentiation by two mechanisms. J Invest Dermatol 104(4): 546-553.
-
Ravi M, Paramesh V, Kaviya SR, Anuradha E, Paul Solomon FD (2015) 3D cell culture systems: Advantages and applications. J Cell Physiol 230(1): 16-26.
-
Mazzoleni G, Di Lorenzo D, Steimberg N (2009) Modelling tissues in 3D: the next future of pharmaco-toxicology and food research?. Genes Nutr 4(1): 13-22.
-
Sun T, Jackson S, Haycock JW, MacNeil S (2006) Culture of skin cells in 3D rather than 2D improves their ability to survive exposure to cytotoxic agents. J Biotechnol 122(3): 372-381.
-
Vinci M, Gowan S, Boxall F, Patterson L, Zimmermann M, et al. (2012) Advances in establishment and analysis of three-dimensional tumor spheroid-based functional assays for target validation and drug evaluation. BMC Biol 10.
-
Oh JW, Hsi TC, Fernando C, Ramos R, Plikus MV (2013) Organotypic skin culture. J Invest Dermatol 133(11): e.14.
-
Kato H, Marcelo CL, Washington JB, Bingham EL, Feinberg SE (2015) Fabrication of Large Size Ex Vivo-Produced Oral Mucosal Equivalents for Clinical Application. Tissue Eng Part C Methods 21(9): 872-880.
-
Dongari-Bagtzoglou A, Kashleva H (2006) Development of a highly reproducible three-dimensional organotypic model of the oral mucosa. Nat Protoc 1(4): 2012-2018.
-
Loh QL, Choong C (2013) Three-dimensional scaffolds for tissue engineering applications: Role of porosity and pore size. Tissue Eng Part B Rev 19(6): 485-502.
-
Harjanto D, Zaman MH (2013) Modeling Extracellular Matrix Reorganization in 3D Environments. PLoS One 8(1): e52509.
-
Klausner M, Handa Y, Aizawa S (2021) In vitro three- dimensional organotypic culture models of the oral mucosa. In Vitro Cell Dev Biol Anim 57(2): 148-159.
-
Brown JL, Johnston W, Delaney C, Rajendran R, Butcher J, et al. (2019) Biofilm-stimulated epithelium modulates the inflammatory responses in co-cultured immune cells. Sci Rep 9(1).
-
Yadev NP, Murdoch C, Saville SP, Thornhill MH (2011) Evaluation of tissue engineered models of the oral mucosa to investigate oral candidiasis. Microb Pathog 50(6): 278-285.
-
Klausner M, Ayehunie S, Breyfogle BA, Wertz PW, Bacca L, et al. (2007) Organotypic human oral tissue models for toxicological studies. Toxicol In Vitro 21(5):938-949.
-
Morse DJ, Wilson MJ, Wei X, Lewis MAO, Bradshaw DJ, et al. (2018) Denture-associated biofilm infection in three- dimensional oral mucosal tissue models. J Med Microbiol 67(3): 364-375.
-
Benson K, Cramer S, Galla HJ (2013) Impedance-based cell monitoring: barrier properties and beyond. Fluids and Barriers of the CNS 10(1): 1-1.
-
Schlage WK, Iskandar AR, Kostadinova R, Xiang Y, Sewer A, et al. (2014) In vitro systems toxicology approach to investigate the effects of repeated cigarette smoke exposure on human buccal and gingival organotypic epithelial tissue cultures. Toxicol Mech Methods 24(7): 470-487.
-
Gursoy UK, Pöllänen M, Könönen E, Uitto VJ (2010) Biofilm Formation Enhances the Oxygen Tolerance and Invasiveness of Fusobacterium nucleatum in an Oral Mucosa Culture Model. J Periodontol 81(7): 1084-1091.
-
Millhouse E, Jose A, Sherry L, Lappin DF, Patel N, et al. (2014) Development of an in vitro periodontal biofilm model for assessing antimicrobial and host modulatory effects of bioactive molecules. BMC Oral Health 14: 80.
-
Nagaraj NS, Vigneswaran N, Zacharias W (2004) Hypoxia- mediated apoptosis in oral carcinoma cells occurs via two independent pathways. Mol Cancer 3(1): 38.
-
Wayakanon K, Thornhill MH, Douglas CWI, Lewis AL, Warren NJ, et al. (2013) Polymersome- mediated intracellular delivery of antibiotics to treat Porphyromonas gingivalis-infected oral epithelial cells. FASEB J 27(11): 4455-4465.
-
Andrian E, Grenier D, Rouabhia M (2004) In vitro models of tissue penetration and destruction by Porphyromonas gingivalis. Infect Immun 72(8): 4689-4698.
-
Shang L, Deng D, Buskermolen JK, Janus MM, Krom BP, et al. (2018) Multi-species oral biofilm promotes reconstructed human gingiva epithelial barrier function. Sci Rep 8(1): 16061.
-
Tribble GD, Lamont RJ (2010) Bacterial invasion of epithelial cells and spreading in periodontal tissue. Periodontol 2000 52(1): 68-83.
-
Oda D, Dale BA, Bourekis G (1990) Human oral epithelial cell culture II. Keratin expression in fetal and adult gingival cells. In Vitro Cell Dev Biol 26(6): 596-603.
-
Oda D, Watson E (1990) Human oral epithelial cell culture I. Improved conditions for reproducible culture in serum-free medium. In Vitro Cell Dev Biol 26(6): 589- 595.
-
Handfield M, Mans JJ, Zheng G, Lopez MC, Mao S, et al. (2005) Distinct transcriptional profiles characterize oral epithelium-microbiota interactions. Cell Microbiol 7(6): 811-823.
-
Oda D, Bigler L, Lee P, Blanton R (1996) HPV immortalization of human oral epithelial cells: a model for carcinogenesis. Exp Cell Res 226(1): 164-169.
-
Saito A, Kokubu E, Inagaki S, Imamura K, Kita D, et al. (2012) Porphyromonas gingivalis entry into gingival epithelial cells modulated by Fusobacterium nucleatum is dependent on lipid rafts. Microb Pathog 53(5-6): 234- 242.
-
Zhang Z, Liu S, Zhang S, Li Y, Shi X, et al. (2022) Porphyromonas gingivalis outer membrane vesicles inhibit the invasion of Fusobacterium nucleatum into oral epithelial cells by downregulating FadA and FomA. J Periodontol 93(4): 515-525.
-
Saito A, Inagaki S, Kimizuka R, Okuda K, Hosaka Y, et al. (2008) Fusobacterium nucleatum enhances invasion of human gingival epithelial and aortic endothelial cells by Porphyromonas gingivalis. FEMS Immunol Med Microbiol 54(3): 349-355.
-
Inagaki S, Onishi S, Kuramitsu HK, Sharma A (2006) Porphyromonas gingivalis vesicles enhance attachment, and the leucine-rich repeat BspA protein is required for invasion of epithelial cells by “Tannerella forsythia.” Infect Immun 74(9): 5023-5028.
-
Edwards AM, Grossman TJ, Rudney JD (2006) Fusobacterium nucleatum transports noninvasive Streptococcus cristatus into human epithelial cells. Infect Immun 74(1): 654-662.
-
Suresh R, Mosser DM (2013) Pattern recognition receptors in innate immunity, host defense, and immunopathology. Adv Physiol Educ 37(4): 284-291.
-
Takeuchi O, Akira S (2010) Pattern Recognition Receptors and Inflammation. Cell 140(6): 805-820.
-
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124(4): 783-801.
-
Hayden MS, Ghosh S (2012) NF-κB, the first quarter- century: Remarkable progress and outstanding questions. Genes Dev 26(3): 203-234.
-
Yang SH, Sharrocks AD, Whitmarsh AJ (2013) MAP kinase signalling cascades and transcriptional regulation. Gene 513(1): 1-13.
-
Krogsgaard M, Li QJ, Sumen C, Huppa JB, Huse M, et al. (2005) Agonist/endogenous peptide-MHC heterodimers drive T cell activation and sensitivity. Nature 434(7030): 238-243.
-
Iancu SI (2016) Epithelial mechanisms in the microbial pathogenesis of periodontal disease [Doctoral dissertation]. [London]: King’s College London.
-
De Nardo D (2015) Toll-like receptors: Activation, signalling and transcriptional modulation. Cytokine 74(2): 181-189.
-
Krisanaprakornkit S, Kimball JR, Dale BA (2002) Regulation of human β-defensin-2 in gingival epithelial cells: the involvement of mitogen-activated protein kinase pathways, but not the NF-κB transcription factor family. J Immunol 168(1): 316-324.
-
Hasegawa Y, Mans JJ, Mao S, Lopez MC, Baker HV, et al. (2007) Gingival epithelial cell transcriptional responses to commensal and opportunistic oral microbial species. Infect Immun 75(5): 2540-2547.
-
Zhang G, Chen R, Rudney JD (2011) Streptococcus cristatus modulates the Fusobacterium nucleatum- induced epithelial interleukin-8 response through the nuclear factor-kappa B pathway. J Periodontal Res 46(5): 558-567.
-
Milward MT, Chapple ITT, Wright HT, Millard JT, Matthews JT, et al. (2007) Differential activation of NF-κB and gene expression in oral epithelial cells by periodontal pathogens. Clin Exp Immunol 148(2): 307-324.
-
Milward MR, Chapple IL, Carter K, Matthews JB, Cooper PR (2013) Micronutrient modulation of NF-κB in oral keratinocytes exposed to periodontal bacteria. Innate Immun 19(2): 140-151.
-
Watanabe K, Yilmaz Ö, Nakhjiri SF, Belton CM, Lamont RJ (2001) Association of mitogen-activated protein kinase pathways with gingival epithelial cell responses to Porphyromonas gingivalis infection. Infect Immun 69(11): 6731-6737.
-
Huang GTJ, Zhang HB, Dang HN, Haake SK (2004) Differential regulation of cytokine genes in gingival epithelial cells challenged by Fusobacterium nucleatum and Porphyromonas gingivalis. Microb Pathog 37(6): 303-312.
-
Stathopoulou PG, Benakanakere MR, Galicia JC, Kinane DF (2010) Epithelial cell pro-inflammatory cytokine response differs across dental plaque bacterial species. J Clin PeriodontolJan 37(1): 24-29.
-
Chen WA, Dou Y, Fletcher HM, Boskovic DS (2023) Local and Systemic Effects of Porphyromonas gingivalis Infection. Microorganisms 11(2): 470.
-
Kocgozlu L, Elkaim R, Tenenbaum H, Werner S (2009) Variable cell responses to P. gingivalis lipopolysaccharide. J Dent Res 88(8): 741-745.
-
Ding PH, Wang CY, Darveau RP, Jin LJ (2013) Nuclear factor-κB and p38 mitogen-activated protein kinase signaling pathways are critically involved in Porphyromonas gingivalis lipopolysaccharide induction of lipopolysaccharide-binding protein expression in human oral keratinocytes. Mol Oral Microbiol 28(2): 129-141.
-
Fujita Y, Nakayama M, Naito M, Yamachika E, Inoue T, et al. (2014) Hemoglobin receptor protein from Porphyromonas gingivalis induces interleukin-8 production in human gingival epithelial cells through stimulation of the mitogen-activated protein kinase and NF-κB signal transduction pathways. Infect Immun 82(1): 202-211.
-
Flak MB, Colas RA, Muñoz-Atienza E, Curtis MA, Dalli J, et al. (2019) Inflammatory arthritis disrupts gut resolution mechanisms, promoting barrier breakdown by Porphyromonas gingivalis. JCI Insight 4(13): e125191.
-
Ramage G, Lappin DF, Millhouse E, Malcolm J, Jose A, et al. (2017) The epithelial cell response to health and disease associated oral biofilm models. J Periodontal Res 52(3): 325-333.
-
Peyyala R, Ebersole JL (2013) Multispecies biofilms and host responses: Discriminating the Trees from the Forest. Cytokine 61(1): 15-25.
-
Stewart PS, Costerton JW (2001) Antibiotic resistance of bacteria in biofilms. lancet 358(9276): 135-138.
-
Johnston W, Rosier BT, Artacho A, Paterson M, Piela K, et al. (2021)Mechanical biofilm disruption causes microbial and immunological shifts in periodontitis patients. Sci RepDec 11(1): 9796.
-
Dabija-Wolter G, Bakken V, Cimpan MR, Johannessen AC, Costea DE (2013) In vitro reconstruction of human junctional and sulcular epithelium. J Oral Pathol Med 42(5): 396-404.
- Diagnosis and Management of Mental Nerve Paresthesia Secondary to Apical Periodontitis of Mandibular Second Premolar: A CBCT Based Case Report
- A Randomized, Double Blinded Clinical Trial to Compare the Effect of Oral Premedication (Diclofenac Potassium or Dexamethasone) on Post-Operative Pain Following Pulpectomy
- Modified Lip Repositioning Technique for the Management of Excessive Gingival Display
- Integral Role of Non-Dental Providers and Fluoride Dissemination
- Root Canal Treatment Rate in Deciduous Teeth Among 6-Year- Olds in the Era of Discontinuing Water Fluoridation - Historical Cohort Study
- The Impact of the Notch1 on the Migratory Capacity and the Expression of E-Cadherin and CyclinD1 in Ameloblastoma Cells