Utilization of Insight in Cumulus Cells Function in the Context of Oocytes Growth, Maturation, Ovulation and Fertilization for the Enhancement of Success Rates in ART by Better Quality Oocytes and Embryos Selection- A Systematic Review
Cumulus cells (CC) get initiated from the granulosa cells (GCs) that are undifferentiated, which differentiate into mural GCs (MGCs) as well as CCs at the time of antrum generation in the follicles by the allocation of residence. CCs represent cells that are supporting cells with regards to oocytes from the milieu, that aids in oocytes growth as well as maturation in the follicles. There is existence of bidirectional connections amongst oocytes with the idea of acquiring maturation as well as embryonic generational competence subsequent to fertilization. The surges of gonadotrophins (FSH as well as LH), result in generation of extracellular matrix (ECM) in CCs, with CCs going via cumulus expansion for aiding in meiosis getting resumed. Here we conducted a systematic review utilizing search engine pubmed, google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like; CC; oocyte maturation; GCs; mural granulosa cells (MGCs); cumulus oocyte complex (COC); zona pellucid(ZP); gap junctions; meiotic maturation of the oocyte; hyaluronic acid (HA); oocyte liberated factors (OLF); BMP15; GDF9; & their downstream targets-HAS2; PGST;GERM1; SMAD2/3; Bone morphogenetic protein receptor typeII (BMPR II); PIA3K/ AKT pathway; cAMP; cGMP; epidermal growth factor receptor(EGFR), ERK1/2;IGF; pentraxin3(PTX3); cumulus expansion; VCAN; connexin 37[Cx37&Cx43; Tsg6; inter-α inhibitor protein); ADAMTS1&MMP2; apoptotic genes (survivin); proapoptotic (caspase3&7) from 1990 to 2022 till date. We found a total of 150 articles out of which we selected 115 articles for this review. No meta-analysis was done. Thus the ultimate function of CCs is that they are implicated in the finishing of oocytes meiotic maturation along with ovulation, fertilization along with following embryonic generational competence besides pregnancy. Hence getting insight in the function of CCs at the time of follicular generation might aid in anticipation of oocytes quality followed by embryonic generational competence iin addition to pregnancy results along with field of assisted reproductive technology (ARTs) for the therapy of infertility.
Introduction
On the existence of great physical ambience in the context of intra follicular milieu cumulus cells (CC) that are surroundings the oocyte, aid in the capacity of oocyte maturation [1]. Acquisition of insight with regards to how the CC can influence the oocyte generation as well as confer protection to the oocyte from deleterious systemic diseases is key with regards to infertility management [2]. Furthermore, injury of the CCs in case of numerous pathogenesis possesses the potential of reduction in rates of fertilization in addition to pregnancy rates [3]. Concentration of more recent investigators has been on invasive technologies that implicated the evaluation of the somatic cells that surround the oocyte namely CCs for the estimation of the factors that are of relevance with regards to oocyte maturation, possess the capacity of anticipation of the oocyte quality in the context of achieving greater rates of oocyte maturation, fertilization, embryo generation along with pregnancy [1, 2, 3]. The aim of this review is the evaluation of CCs function in addition to its parameters which have been implicated in oocyte quality in the context of generation of the granulosa cells (GCs) along with oocyte, how they aid in fertilization in addition to early embryogenesis, besides anticipation of pregnancy in assisted reproductive technology (ARTs). With our endeavour of getting greater insight in advances in physiology of normal menstrual cycle, role of Bone morphogenetic protein BMP) and Growth Differentiation factors (GDF) with regards to their parts in the development of follicles in the human menstrual cycle with its clinical impact on ART success along with PCOS pathogenesis as well as therapy, improving success in ART with the utilization of artificial intelligence in addition to utilization of Extracellular Vesicles (ECV) in for escalaton of the success [4, 5, 6, 7]. Here we further tried to get an insight with regards to role of CCs function for enhancement of success of ART.
Methods
Here we conducted a systematic review utilizing search engine pubmed, google scholar; web of science; embase; Cochrane review library utilizing the MeSH terms like; CC; oocyte maturation; GCs; mural granulosa cells (MGCs); cumulus oocyte complex (COC); zona pellucid (ZP); gap junctions; meiotic maturation of the oocyte; hyaluronic acid (HA); oocyte liberated factors (OLF); BMP15; GDF9; &their downstream targets-HAS2; PGST; GERM1; SMAD2/3; Bone morphogenetic protein receptor type II (BMPR II); PI3K/ AKT pathway; cAMP; cGMP; epidermal growth factor receptor(EGFR), ERK1/2; IGF; pentraxin3(PTX3); cumulus expansion; VCAN; connexin 37[Cx37&Cx43; Tsg6; inter-α inhibitor protein); ADAMTS1&MMP2; apoptotic genes(survivin); proapoptotic (caspase 3&7) from 1990 to 2022 till date.
Results
We found a total of 150 articles out of which we selected 115 articles for this review. No meta-analysis was done.
Defining Cumulus Cells as well as the Differentiation of Granulosa Cells
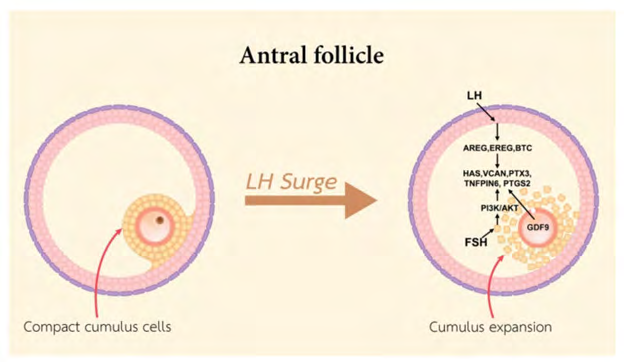
Basically CCs, represent the somatic cells that surround the oocyte. They play a significant part in the oocyte growth, meiotic maturation, ovulation in addition to subsequent fertilization in mammals [8]. The origination of CCs occurs from the undifferentiated granulosa cells (GCs) from primordial to preantral follicles. GCs are the primary cell kinds in the ovary which yield physical re inforcement / buttressing in addition to provision of milieu needed for the oocyte that is growing [9]. The preantral follicles GCs can get classified into two populations, namely i) mural granulosa cells (MGCs) along with CCs. The wall of the follicle gets lined by MGCs, whereas, CCs are correlated with the oocyte. The initial layer of CCs that surrounds the oocyte is known as corona radiata, with CCs generating a pseudostratified epithelium that is connecting with the oocyte by membrane extensions that surround the corona cells. At the time of early antral follicle generation, the structure of the cumulus oophorus gets generated by undifferentiated GCs. MGCs reside on the wall of the antral follicle, as well as CCs are the ones existent directly adjacent to the oocyte (figure1) (rev in ref [10]). CCs in direct connection with the oocyte generate an oocyte i. e cumulus oocyte complex (COC). This COC aids in crosstalk amongst the oocytes along with CCs by direct impact on gene expression as well as protein generation, resulting in differentiation in addition to cumulus expansion besides oocyte maturation. CCs communication with the cytoplasm takes place along with penetration of the zona pellucid (ZP) with gap junctions [11]. Subsequent to generation from the pre antral follicle to the CCs which go via proliferation along with the gap junctions get ultimately liberated from the ooplasm with meiotic maturation of the oocyte.
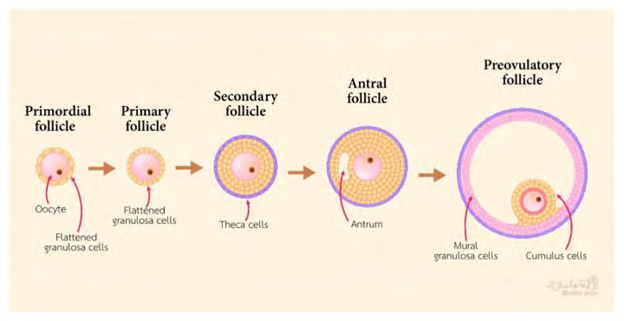
Besides the anatomical variations in the context of where they reside in the follicle, both CCs as well as MGCs are separate based on the functional aspects [12]. Under normal situations function of CCs is provision of support to the oocyte growth, generate hyaluronic acid (HA) along with once stimulation with FSH is there go through cumulus expansion, whereas MGCs perform endocrine functions, besides cooperation with the follicle generation [13]. Generation by CCs of HA occurs as well as they become expansive, whereas MGCs do not generate HA in the absence of expansion [14]. Subsequent to LH surge, the structure of the cumulus oophorus expands further by accrual of abundant HA in the extracellular matrix [ECM] that aids in ovulation in addition to following fertilization that takes place in the ampulla of the oviducts [15].
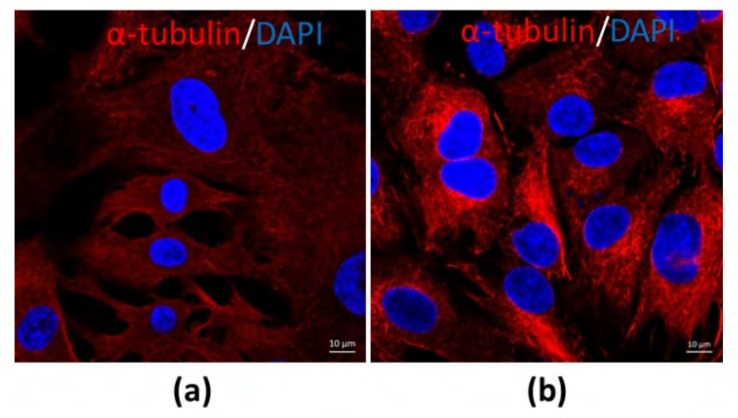
The morphological structure as well as function might be variable once the sizes of CCs as well as MGCs are from various sizes of follicles along with in various phases of menstrual cycle. COC’s from small antral follicles in the absence of stimulation with follicle stimulating hormone (FSH), Luteinizing hormone (LH) surge possess greater probability of displaying undifferentiated form. Nevertheless, COC’s from pre-ovulatory follicle/ dominant follicle in particular subsequent to an LH surge canonically illustrated a differentiated form. At this point of time, CCs along with MGCs are unique cells in the follicles. Morphologically MGCs are compact, possessing dark cytoplasm with the shape of an epithelial like cell (figure 2a). Conversely, CCs are loose, have clear cytoplasm, besides round in shape (figure 2b) on staining with alpha tubulin for observing [unpublished data).
The generation of antral follicles gets stimulated by gonadotrophins in addition to oocyte liberated factors (OLF). Preantral GCs are the usual precursors of CCs along with MGCs in addition to antral generation segregates the GCs into MGCs as well as CCs [12]. The generation of CCs gets controlled by OLF with in the follicles, while the generation of MGCs is controlled by the stimulation from the exterior follicle i. e. from the anterior pituitary gland [16]. FSH is a necessary hormone in the follicle with regards to the proliferation, growth along with differentiation of GCs by enhancement of vascularisation of the theca interna layer of cells that lie peripheral to the basal lamina, the generation of in the context of proliferation along with differentiation of GCs by the escalation of follicular fluid, in addition to the differentiation of MGCs along with CCs [17].
This differentiation of undifferentiated GCs into MGCs gets stimulated by FSH in an intricate crosstalk, with, insulin like growth factors (IGFs) [18, 19]. The FSH induction of differentiation of MGCs possesses key part in fertility besides resulting in the preparation of female reproductive organs in the context, of transportation of sperm, fertilization along with early embryonic generation [20]. FSH causes stimulation of the Luteinizing hormone receptor (LHR) mRNA expression in MGCs. LHR mRNA is a crucial marker of MGCs differentiation [21]. The amounts of the mural marker transcript LHR are greatest in the neighboring basal lamina, with least in the CCs [22]. In the form of MGC phenotype, FSH causes stimulation of the MGC transcripts implicated FSH in steroidogenesis (Cyp11a1), ovulation ((LHR) along with immune function (Cd34). In case of bovine GCs, IGFs has a part in follicle control which works with FSH to control GCs growth. There is significance of FSH IGFs as well as OLFs for the deciding of the MGCs besides CCs lineages, driving the differential expression of gene properties of MGCs or CCs phenotypes [23].
Conversely, the CCs in the preovulatory follicle escape from the differentiation actions of FSH along with IGFs by OLFs [16]. FSH results in repression of CCs transcripts like S1c38a3 along with the antimullerian hormone (AMH) mRNA amounts in COC, along with stimulation of mural transcripts, whereas the oocyte stimulated SMAD 2/3 signaling causes facilitation of escalated amounts of cumulus marker transcripts in CCs like via Growth Differentiation factor 9 (GDF9) along with Bone morphogenetic proteins15 (BMP15) liberated by the oocyte (Figure 3) [24]. Binding of GDF9 to TGF-beta takes place with type1 receptor kinase (ALK5) [25], as well as Bone morphogenetic protein receptor type II (BMPR II) [26], whereas BMP15 binding takes place to the receptor ALK6 as well as BMPR II. GDF9, along with BMP15 participate in crucial part in growth, differentiation along with function of GCs along with theca cells at the time of early follicles generation that has a key part in generation of oocyte, ovulation, fertilization along with, embryonic competence [27, 28]. In mice it was documented that CCs need OLFs via the crosstalk of GDF9 with its receptor, apart from activating extracellular signal –regulated kinase (ERK1/2) via epidermal growth factor receptor (EGFR), which in coordination caused facilitation of SMAD2/3 modulated gene [29] transcription. It is apparent that human CCs in a reaction to FSH need the IGF1R action with following, protein kinase B (AKT) activation, with this action of FSH was ameliorated, on hampering IGF1R action [23]. In case of the loss of FSH, the action of FSH might further end the CCs proliferation prior to ovulation.
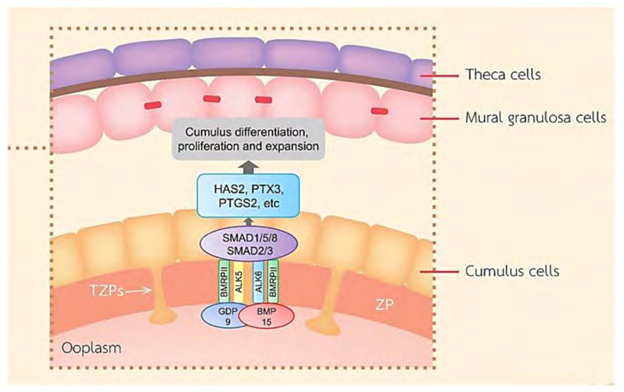
Figure 3: Courtesy ref no-10-A schematic diagram exhibiting roles of the main TGF-β superfamily member proteins in regulating ovarian function. BMP-15 and GDF-9 promote cumulus marker transcripts. GDF-9 and BMP-15 are important for the function of GCs, CC differentiation and proliferation, COC expansion, and hyaluronan production leading to ovulation.
The transcriptomes of CCs as well as MGCs in large mouse follicles, these CCs were rich in transcripts correlated with metabolism along with proliferation of cells, whereas MGCs were rich in transcripts implicated in cell signaling along with differentiation [30]. In case of humans the contrasting amongst CCs as well as MGCs illustrated important functional variations in these cell kinds. CCs are implicated, in steroidogenesis chamber, whereas MGCs are responsible in the angiogenesis chamber [31], while in mice, the expression of SLC38A3in CCs modulates the transportation of particular amino acids from the CCs to the oocyte for ensuring the growth of the oocytes [32]. The expression of androgens receptor (AR) in the follicles is needed for fertilization. Depletion of AR gene in mice resulted in infertility along with CCs aberrations in morphology as well as differentiation [33]. Prior studies demonstrated that the hampered SMAD 2/3 action resulted in blockade of the capacity of the oocytes to cause stimulation of proliferation of CCs [34], along with Smad 4 gene depletion in GCs illustrated abnormalities in the differentiation of CCs [35]. Hence for the differentiation of GCs to CCs requires particular factors along with causing repression of the stimulation of MGCs generation.
The Crosstalk of Oocytes as well as Cumulus Cells
In the case of oocytes growth, the part of CCs is to synergise the nuclear along with cytoplasmic maturation in addition to control the resuming of meiosis by yielding numerous factors to the oocytes like cyclic adenine monophosphate (cAMP) [36], cyclic guanosine monophosphate (cGMP) [37], along with controlling molecules [38, 39]. The second messengers namely cAMP as well as cGMP get generated by CCs that are implicated, in crosstalk, amongst CCs along with oocytes that gets modulated by gap junctions. Subsequent to birth, GCs ensure that oocyte s are arrested in the prophase stage of meiosis 1. Following, puberty GCs proliferation along with differentiation takes place towards CCs to provide nutrition in addition to result in the meiosis getting resumed [35]. The initial step for meiosis getting resumed occurs secondary to gonadotrophins, by activation of the GCs that are surrounding. As far as mammals are concerned, FSH along with LH effects on the meiosis of oocytes getting resumed modulates an escalation of cAMP generation through the stimulation of mitogen activated protein kinase (MAPK), [31, 36, 37], that works on the GCs along with facilitation of an escalation of cAMP amongst the granulosa cells chamber in addition to a reduction in cAMP as well as cGMP with in the oocytes by reduction in the permeability of the gap junctions amongst CCs along with oocyte, hence stimulating the meiosis getting resumed in addition to cumulus cells expansion for preparing for ovulation along with fertilization [37, 40].
In view of the incapacity of glucose generation by oocytes, the metabolism of glucose into pyruvate gets carried on by CCs that is followed by transportation of pyruvate as well as energy to the oocytes with regards to oocyte maturation [23]. The BMP 15 as well as GDF 9 that is generated by oocytes have been implicated in control of the glycolysis along with production of in view of GCs being the providers of nutrition to the oocytes with pyruvate, lactate along with the products that get obtained secondary to the production pathway of cholesterol through gap junctions [41]. Lipid droplets constitute the major structure that ensures sustenance in the context of mammalian oocyte maturation along with is followed by pre-implantation of embryonic production. In the case of bovine CCs metabolites, usually lipids were observed in CCs along with a reduction in lipid classes like phosphatidylcholine as well as phosphatidylethanolamine in CCs at the time of CCs maturation, that, pointed that a lipid metabolism coordination was existent amongst the CCs along with oocytes [42]. Perilipin 3 (PLIN3) is a factor that is implicated broadly for working for lipid production as well as control. Wide distribution of lipid droplets has been the observation amongst the CCs along with oocyte. Expression of greater quantities of PLIN3 has been seen in mature oocytes, with lipid droplets accrual at the time of oocyte maturation getting afflicted by PLIN3 cross interaction amongst CCs along with oocytes [43]. Interference with the crosstalk amongst CCs along with oocytes resulted in bad generational probability along with/or infertility [44].
The crosstalk amongst CCs along with oocytes implicates gap junctions, besides numerous signaling factors [45]. Gap junctions that are located amongst CCs along with oocytes are labeled as transzonal projec tions TZPs), thin cytoplasmic extrusions that aid in communication of CCs with the oocytes as well as are key for normal, oocyte generation [46]. The gap junctions unit located amongst CCs along with oocytes namely GJA4 (gap junction protein α4 alias connexin 37[Cx37]), that represents the main isoform of connexins. Gap junction associated protein α1 (GJA1 alias connexin 43[[Cx43]), represents the main isoform of connexins that is located amongst GCs); MGCs - MGCs along with MGCs- CCs. These gap junction cause transportation of nutrients as well as small molecules like ions, metabolites, amino acids in addition to intracellular signaling molecules from GCs to oocytes through CCs [23]. In case of GCs cGMP is responsible for sustenance of TZPs in the preovulatory phase. At the time of ovulation, binding of LH to LHR in the MGCs occurs, that results in the stimulation of liberation of EGFR ligands that subsequently cause the activation of ERK/MAPK, thus result in actin cytoskeleton getting reorganized along with retractions of TZPs into cell body of CCs [47]. In mice it has been documented that gap junctions are essential for the in vivo as well as in vitro maturation of oocytes, the existence of CCs at the time of insemination caused escalation of fertilization that was followed by the production of the blastocyst, with this escalation being controlled by gap junctions [48]. At the time of meiosis of oocytes getting resumed, a reduction in the amount cAMP in the oocytes was brought about by LH surge. This LHsurge causes the active CD44 that takes part in the phosphorylation of gap junction proteins, as well as the rupture of gap junctions amongst CCs along with oocyte resulting in a reduction of cAMP shift to the oocyte, hence causing stimulation of meiotic maturation [49].
Furthermore, oocytes aid in provision of OLFs with the idea of GCs proliferation, differentiation, apoptosis, metabolism, steroidogenesis along with expansion of CCs by BMP 15 as well as GDF 9[29, 50]. GDF 9 along with BMP 15 represent 2 members that belonged to the transforming growth factor beta (TGF-β) superfamily members whose expression occurs in oocytes [46]. Association amongst GDF 9 along with BMP 15 in the form of GDF 9 / BMP 15 heterodimers, labelled as cumulin, work in the form of controllers of GCs as well as CCs function besides result in escalation of oocyte quality [28]. At the time of oocyte maturation, GDF 9 along with BMP 15 work on CCs with the aim of induction of expansion stimulating factors. OLFs in addition to LH further result in avoidance along with reduction in apoptosis, in the CCs. In case of COC, binding of cumulin to its receptor BMPRII, for avoidance of CCs demise [51, 52]. Hence the collection network that is implicated in the COC control would have physiological relevance in the context of oocyte growth, oocyte maturation along with ovulation, along with following fertilization as well as embryonic production competences.
The Mode of Cumulus Expansion as well as Ovulation
The cumulus extracellular matrix [ECM], is basically the generation of HA, pentraxin3 (PTX3), Tumor necrosis factor (TNF) stimulated gene 6(TSG6 alias TNFAIP6) as well as heavy chains (HCs) of serum obtained inter-α inhibitor protein [53]. The intracellular matrix amongst CCs comprises of HA, bound to the cell surface receptors (CD44 as well as Rhamm). Stabilization in addition to organization of HA into a particular structure. Binding of each one of Versican (VCAN), Tsg6 along with inter-α inhibitor HC to HA takes place, while PTX3 cross talks with numerous Tsg6 molecules. VCAN represents an extracellular matrix proteoglycans that cross talks with integrins along with cell surface proteins via its C-terminal area along with anchoring the matrix of HA to the CCs. The protease ADAMTS1 (a dis-integrin as well as Metalloproteinase that possesses, the thrombospondin motif) causes cleavage of VCAN in the β glycosaminoglycans (GAG), i. e a chondroitin sulphate replacement midsection, a domain that causes production of HA binding fragments that finish with the neoepitope DPEAAE at the C-terminal area in this matrix. The Versican GI - DPEAAE alias versikine, works in the organization of the HA matrix [54]. Probably it modulates the COC matrix structure along with function besides its part in ovulation [55]. The cumulus expansion is based on the glycosaminoglycans production in the HA in ECM, where its part is in the form of a structural constituent of cumulus expansion along with a signaling molecule that controls the oocyte maturation. Generation of ECM molecules by CCs causes cumulus expansion, that is necessary for ovulation, as well as fertilization, besides being an anticipator of oocyte quality [56]. Enough CC layers, with sufficient generation of HA with subsequent cumulus expansion is essential for oocyte maturation [56].
At the time of transition from pre antral to antral follicles, oocyte correlated granulosa cells attain competence for undergoing cumulus expansion. OLFs cause facilitation of differentiation of the pre antral GCs to CCs like BMP 15 [50] along with GDF 9 [57]. In case of samples obtained of bovine origin, BMP 15 as well as GDF 9 work on un differentiated GCs for facilitation of production of antrum like structures. Absence of GDF 9 along with BMP 15 results in undifferentiated GCs in addition to antrum abnormalities [58]. The induction of different genes that is inclusive of HA- synthase 2(HAS2), cyclooxygenase 2[COX2], prostaglandin endoperoxidase synthase 2 (PTGS2), GREM1 along with of steroidogenic acute regulatory protein (StAR), as well as repression of LHR, that is significant for the follicle generation along with cumulus expansion [35, 57]. Hence the control of GDF 9 as well as its downstream factors in CCs might aid in anticipation of oocyte quality besides oocyte health. Intriguingly, the innate immune reaction along with cytokines generation CD34 antigen along with pathogen– recognition receptors are the members of toll like receptor (TLR) family which cause stimulation of cumulus expansion. PGE2 as well as AREG result in induction of expression of interleukin-6( IL-6) in mouse CCs, with IL-6 working as acontroller of cumulus expansion. In case of mice along with porcine cumulus expansion SMAD 2/3 signaling activation occured by EGFR as well as OLFs [59]. SMAD 2/3 activation results in stimulation of generation of HA in addition to proteins implicated in matrix expansion. FSH- EGFR results in activation of SMAD 2/3 signaling pathway. that is implicated in the control of cumulus expansion as well as steroidogenesis. FSH results in escalation of EGF stimulated tyrosine phosphorylation of EGFR along with results in stimulation of particular EGFR controlling proteins. Moreover, the production of HA along with progesterone get stimulated by FSH. Subsequently SMAD 2/3 activation by GDF 9 as well as TGF-beta influence gonadotrophins stimulated HA along with progesterone generation by porcine CCs [59].
Furthermore, OLFs are needed for ensuring escalation of HAS2, PTGS2, PTX3, along with Tnfaip6 transcripts at the time of cumulus expansion [60]. These transcripts are needed for the purpose of cumulus expansion, since the phenotype of null mutations in PTGS2, PTX3, or Tnfaip6 genes or the hampering of HAS2, results in robust interference with cumulus expansion [16, 44, 61]. Numerous studies have documented that HAS2 is essential for the differentiation in addition to expansion of CCs along with is associated with early embryogenesis. The generation of PTX3 gene is an ECM protein that crosstalks with the HA in the expanded cumulus matrix [62]. Both CCs along with GCs generate TNFAIP6 in the preovulatory follicle [58]. Inter -α trypsin inhibitor (IαI) as well as TNFAIP6 are implicated in ECM production of CCs at the time of cumulus expansion by stabilization of HA chains. The cleavage of IαI into heavy chains (HC) HC1 along with HC2 is necessary for the binding of HCs along with HA in addition to cumulus expansion stabilization [57]. The crosstalk of TNFAIP6 with HCs is necessary for greater stabilization of cumulus expansion. This binding is based on the crosstalk of TNFAIP6 with PTX3, whose up regulation is brought about by GDF9 along with generated at the time of cumulus expansion into ECM [63, 64]. At the time of the ovulatory phase, cumulus expansion as well as oocyte maturation takes place in the pre ovulatory follicles. The event of cumulus expansion needs the existence of factors that cause facilitation of cumulus expansion whose liberation takes place from the oocyte. Cumulus expansion implies the generation of HA –enriched ECM by HAS2, PTX3, Tnfaip6 along with VCAN [58]. Cyclic nucleotides (inclusive of cAMP, cGMP) are implicated in sustenance of oocyte arrest. Furthermore recently The extracellular matrix (ECM) is an essential structure with biological activities. It has been demonstrated that the ECM affects gene expression through cytoskeletal components as well as the gene expression is based upon cell crosstalk with molecules as well as hormones. The generation of ovarian follicles is a hormone based event. The surge in the luteinizing hormone triggers ovulatory changes in oocyte micromilieu. Furthermore, recently Nagyova, et al. [65] reviewed, how proteolytic cleavage impacts the generation of cumulus ECM subsequent to hormonal stimulation; specifically in, how the particular proteasome inhibitor MG132 influence gonadotropin- stimulated cytoskeletal structure, the organization of cumulus ECM, steroidogenesis, along with nuclear maturation. Their observation was that subsequent to the hampering of proteolytic cleavage, gonadotropin-stimulated oocyte–cumulus complexes (OCCs) were without any signs of cumulus expansion; they remained compact with preserved cytoskeletal F-actin-rich transzonal projections through the oocyte investments. Concomitantly, a significant reduction was seen in progesterone liberation along with in the expression of gonadotropin- induction of cumulus expansion–associated transcripts, like HAS2 as well as TNFAIP6. In agreement, the covalent binding amongst hyaluronan as well as the heavy chains of serum- obtained the inter-alpha-trypsin inhibitor, necessary for the organization of cumulus ECM, was missing [65]. (Figures 5 & 6) Sustenance of greater quantities of intracellular cAMP gets enabled through both the oocyte modulated production, in addition to the cAMP influx, that gets generated by the GCs that are surroundings via the communicating gap junctions [66]. At the time of LH surge EGFs, that are inclusive of the amphiregulin (AREG), epiregulin (EREG), BTC2 as well as PTGS2 get produced by the GCs along with hampering the generation of cAMP, as well as cGMP. Subsequently, a fall of cAMP in the oocytes along with resumption of meiosis propagation by protein kinaseA (PKA) dephosphorylation as well as stimulation of the mitosis promoting factor (MPF) along with meiotic maturation along with nuclear envelope break down (NEBD), in addition to oocyte maturation [67, 68]. The binding of LH to LHR in the MGCs causes induction of the expression of EGFs followed by the activation of EGFR, KRAS, along with ERK1/2 in MGCs as well as CCs. Furthermore, activated ERK1/2 results in stimulation of PTGS2, StAR, HAS2, as well as Tnfaip6 [69]. The LH surge results in cumulus expansion, whose modulation takes place by EGFs like peptides (EGFLPs) that is generated by MGCs along with causing the stimulation of expression of EGFLP [70]. AREG, EREG), BTC2 work directly on MGCs as well as which results in stimulation of resumption of meiosis, cumulus expansion followed by ovulation [71]. At the time of oocyte maturation, metabolomic alterations in bovine CCs have been documented that suggests that maximum increment of amino acids quantities in particular of serine. Probably serine is necessary for the Oxidative stress reduction through channeling the serine stores that get lost towards glutathione production in the oocyte. Utilization of serine can be done, for development of Single carbon units with the aim of nucleotide generation like glycine or α-ketoglutarate [42]. In the case of human CCs derived from mature oocytes long non coding RNA(lnc RNAS) that are inclusive of NEAT1, MALAT1, ANXA2P2, MEG3, IL6STP1 along with VIM-AS1 are implicated in apoptosis as well as ECM- associated functions that are necessary for growth of the oocytes (rev by us in ref [72]). Hence the lnc RNAS that are expressed in CCs might be implicated in control of necessary pathways that aid in the human oocyte maturation, fertilization in addition to embryonic generation, besides making provision for biomarkers for anticipation of oocyte quality, with the ideas of generation of noninvasive tests for Identifying the embryos that possess, the capacity of maximum generational probability [70].
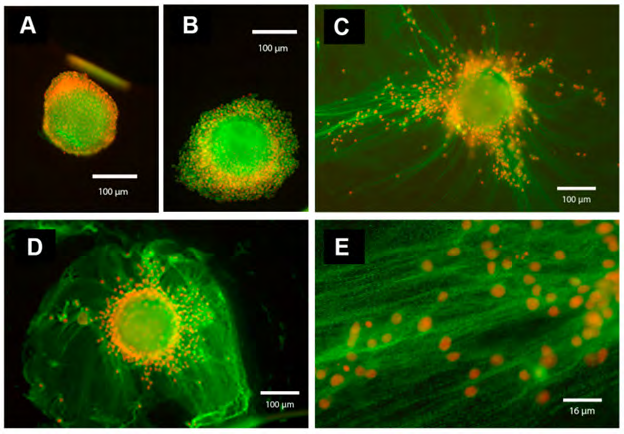
Oocyte-cumulus complexes (OCCs) were cultured for 20 h without (A) or with (B–E) FSH (100 ng/mL) in the absence (A, B; in PVP-supplemented medium) or in the presence of serum (5 % FBS) (C–E, magnification of ECM structure present in C). In the absence of serum (A,B), the cumulus cells remain close to each other and to the oocyte, and the ECM is absent. In contrast, in the presence of serum and FSH (C–E, magnification of the cumulus ECM structure present in C), cumulus cells synthesize and organize an ECM structure that contains components immune reactive for IαI and present as a network of cables. The immunofluorescence analysis was performed by using a rabbit anti-human IαI antibody (dilution 1:100; DAKO, Carpenteria, CA, USA) and Alexa Fluor 488 goat anti-rabbit IgG as a secondary antibody (dilution 1:500; Molecular Probes, green). Nuclei are in red.
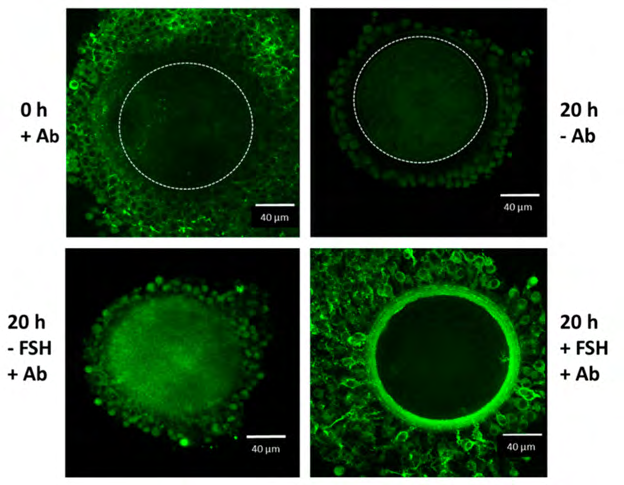
Figure 5: Courtesy ref no-67-Confocal microscope analysis of IαI in the oocyte-cumulus complexes. Oocyte–cumulus cells complexes were analyzed at the beginning (0 h) and at the end (20 h) of the culture carried out in the presence of 5 % FBS, with or without FSH (100 ng/mL) stimulation, as indicated. Immuno reactivity for IαI components was evident in the ECM structure organized around the cumulus cells after FSH stimulation by using a rabbit anti-human IαI antibody (Ab dilution 1:100; DAKO, Carpenteria, CA, USA) and Alexa Fluor 488 goat anti-rabbit IgG as a secondary antibody (dilution 1:500; Molecular Probes, green). The boundary of the oocyte is marked by a dashed line.
Additionally, FSH causes the induction of AREG, EREG gene expression along with stimulation of cumulus expansion in pigs [73], cattle [74], along with humans [75]. The mode by which FSH causes the induction of expansion takes place by an escalation of cAMP in CCs in addition to enhancement of HAS2 gene expression [76]. FSH causes the stimulation of cumulus expansion association to the accrual of GAG in the ECM. In case of pigs the effect of IGF1 which is acclaimed to activate PI3K / AKT signaling, results in FSH stimulated generation of HA within the expanded ECM by (PI3K) /v- AKT murine thymomoa viral oncogene homolog (AKT), besides mitogen activated kinase3 as well as 1 (MAPK3/1)- based modes [59]. The action of PI3K / AKT is necessary for gonadotrophin stimulated cumulus expansion in vitro. The control of expression of HAS2, as well as PTGS2 in CCs is controlled by the presence of an FSH stimulated along with a PKA independent pathway [77]. Any changes in cumulus expansion is deleterious for reproduction, directly or in the form of a reduction in functional along with structural qualities of the oocyte (figure 5), hence it might result in disturbance of the motion of the COC ovulation process.
Cumulus Cells as well as Fertilization
In view of the inclusion by CCs of gap junction generation, nutrients along with hormonal transportation to the oocyte, CCs are further associated with the sperm physiology by resulting in alterations in sperm, aiding in fertilization hence escalation of fertility [78]. Calcium (Ca2+) mobilization in CCs is implicated in the event of oocyte maturation as well as acrosome reaction. LH stimulated escalation of Ca2+ in CCs causes enhancement of cAMP concentration, besides stimulation of the MAPK pathway in CCs that is implicated in the expression of EGF like factors, besides impacting the oocyte maturation. At the time of fertilization, penetratation of the cumulus as well as ZP occurs by sperm. The selection of the motion of the sperm towards the oocyte gets carried out by CCs as a selective barrier function. Furthermore, liberation of progesterone by CCs which works as a stimulator of acrosome reaction, that gets stimulated by the cell surface receptor NYD-SP-8 stimulated Calcium (Ca2+) mobilization along with a part in fertilization by regulation of the binding of the sperms to the ZP [79]. Progesterone causes stimulation of the hyperactivation of the flagellar motion along with acrosome reaction [80]]. Prior studies demonstrated that in mice provision of factors by CCs influence sperm functions.
Prostaglandins, PGE1, PGE2, PGE2a were the observation in the incubation medium of COCs. On blocking the formation of Prostaglandins by indomethacin caused a reduction in fertilization rate [81].
Metalloproteinase represent a protease enzyme that possesses a catalytic mode. ADAMTS1, Matrix Metalloproteinase 2(MMP2) Tissue inhibitors of Matrix Metalloproteinases (TIMP1) are the Metalloproteinases implicated in fertilization [rev by us in ref [82]]. In case of mice, ADAMTS1, that is an extracellular metalloproteinase, results in the stimulation of ovarian follicles with the aid of ovulatory hormones as well as is necessary for fertility. A reduction in ovulation rate by 77%, in addition to fertilization rate by 63%occurred inAdamts1 -/-mice. Hence the need for ADAMTS1 in the events of ovulation along with fertilization in vivo is existent [83]. MMP2 is implicated in the degradation of ECM, in case of women with infertility, whose expression is higher in CCs, with an escalation of TIMP1 that constitutes a hampering molecule which controls MMPs along with ADAMTS1 in poor ovarian reserve subjects, in addition to a reduction in fertilization rate might be associated with reduction in fertility [84]. GJA1 along with serpin peptidase clade E(SERPINE2)are gene markers that have a probable correlation with the oocyte maturation, as well as PTRSS35 might be associated with the capacity of fertilization of oocyte [85]. The generating acrosome in spermatids possesses pituitary adenyl cyclase activated polypeptide (PACAP). The binding of PACAP type1 receptor that is existent in post ovulatory CCs with PACAP results in induction of acrosome reaction in mice [86].
PACAP results in escalation of penetration of sperm via the cumulus layer along with ZP. Liberation of a soluble factor from the CCs in reaction to PACAP results in stimulation of sperm motility, acrosome reaction along with fertilization [87]. In case of females PTX3, has a significant part in innate immunity against particular pathogens as well as fertilization. Production of PTX3 by CCs in mice, at the time of cumulus expansion, resides in the matrix. Furthermore, in humans expression of PTX3 takes place in CCs. in the context of mice with Ptx3-/- there is Correlation of infertility with robust aberrations of CCs, besides failure of oocyte fertilization in vivo. Normal concentrations of HA generation occurs in mice with Ptx3-/- although there is inability of stabilization of matrix. Hence PTX3 is a structural constituent of CCs along with ECM is necessary for female fertility [64]. Inter-α- inhibitor (ITIH) that is a proteoglycans is needed for mammalian reproduction. It comprises of two heavy chains (HC) HC1 along with HC2. Before ovulation, shift of HCs takes place onto HA for the production of HC- HA complexes along with an ECM generation as well as stabilization [85].
An earlier study illustrated that the expression of serine protease 35 (PTRSS35) takes place uniquely in the ovary, with the mRNA concentrations were correlated with the probability of oocyte fertilization. The mRNA concentrations of PTRSS35 in case of CCs of fertilized oocytes were significantly greater in contrast to unfertilized oocytes [85]. Partial restoration of the cleavage rate of the fertilization of the oocyte that were denuded, pointed that CCs liberated factors were significant for fertilization, however, attachment amongst oocytes as well as CCs is needed for the ideal fertilization as well as initial cleavage [84]. Human leukocyte antigen-G (HLA-G) possesses a part in the oocyte maturation along with embryo implantation. The observation in Human CCs was that lower expression of HLA-G correlated with bad oocyte quality in addition to lesser fertilization along with reduction in the generation of embryos [88]. Prior to fertilization the exposure of COC to hydrogen peroxide resulted in reduction in the cleavage rate, however, not in the demise of CCs or oocytes. Conversely, exposing oocyte that were denuded, (DO)to H2O2 causes total oocyte demise as well as total blockade of first cleavage [89]. Hence the CCs confer protection to oocytes from Oxidative stress (OS) at the time of fertilization
Cumulus Cells as well as Embryonic Generation
Selection of embryos that possess, the capacity of implantation has persisted as a main hurdle in infertility therapy while conducting assisted reproductive technology (ART). This selection is based on themorphological criteria of the oocytes along with embryos, like growth rate, early cleavage on day1, amounts of fragmentation along with blastocyst generation [90]. Numerous factors influence oocyte quality along with result in recurrent implantation failure that is implied. The regulation of nuclear along with cytoplasmic maturation of the oocytes are carried out by these factors via complicated events [1]. In view of the key part of CCs in the follicular generation, growth of oocytes, numerous studies illustrated that oocytes quality got estimated by viability of the CCs. Hence the various studies associated gene expression in cumulus cells with oocytes maturity, fertilization, embryonic generation, implantation in addition to pregnancy have been conducted [91].
During the cleavage stage, removing of CCs prior to IVF in bovine samples resulted in significant reduction in cleavage rate (25% in denuded oocytes (DO) vs 56% for COCs). Hence the depletion of CCs prior to IVF influenced the cleavage rate via depletion of a factor that is liberated by these cells. The maximum probability of this factor is progesterone [1]. Earlier studies with regards to transcriptomes in human CCs documented that in case of 611 genes that had differential expression in CCs from early along with non-early cleavage embryos, over expression of 24% existed in early cleavage in CCs. These genes were implicated in various signaling pathway that is inclusive of cell cycle, survival as well as demise signaling, chemokines as well as cytokines signaling, angiogenesis as well as lipid metabolism [91]. Phosphorylation controls the action of 7-dehydro cholesterol reductase (DHCR7) in addition to glutathione peroxidase (GPx), in CCs along with is necessary for embryonic generation. GPx is seen in a hypoxic milieu. Hypoxia as well as ROS in follicular fluid have a negative correlation with embryonic generation, pregnancy results, in addition to a significantly greater incidence of aneuploidy along with spindle abnormalities in oocytes [92]. Cholesterol works as the main precursor of progesterone along with estrogen generation, gets its own production from the induction of DHCR7. Hampering of cholesterol production in COCs caused a fall of progesterone concentrations, that resulted in a reduction in the rates of GVBD that is significant for embryonic generation [93].
The transient receptor potentialcation channel subfamily M, member 7 melastin (TRPM7) represents a protein which is controlled by EGF as well as inositol- triphosphate-3-kinase A (ITPKA). Crosstalk of TRPM7 with calmodulin2 downstream of EGF was seen for cleavage stage embryo generation [94, 95]. The reduction in cleavage along with blastocyst generation rates were documented in DO in contrast to intact COCs, that pointed that the reduction in first cleavage influenced the blastocyst generation rates for the reduction in DOs, however not the following, embryonic generation in bovine oocytes samples [89].
In view of the size of follicles generation, the small follicles of embryos in heterogenous cycles possessed a probability of incapacity of reaching the blastocyst in contrast to larger follicles of embryos in homogenous cycles [96]. Cumulus cell genes are associated with the blastocyst generation, along with HAS2, as well as GDF9 were the ones having greater expression in CCs from oocytes that generated into better quality blastocysts [91]. Embryonic generation is associated with the expression of the three gene that are correlated with GDF9 (HAS2, PTGS2 as well as, GREM1). The expression of these cumulus genes demonstrated a greater expression in case of oocytes progressing into embryos that were of higher quality (grade 3-5) in contrast to embryos that were of lower quality (grade1-2). Thus corroborating that the morphology of human embryos, subsequent to 3 days of culture is based on the expression of these CCs genes [96].
In a separate study that involved the evaluation of gene expression in humans with the utilization of reverse transcriptase-Polymerase Chain reaction (RT-PCR), the observation was that GREM1 along with HAS2, pointed towards higher quality embryos, however not PTX3 in higher quality embryos [18]. A greater PTX3 gene expression was seen in human COCs in the normal embryo generation on day3 in contrast to oocytes that had failed to fertilize [97]. Bone morphogenetic protein (BMP2) possesses functions in generation of the oocytes along with other follicular cells in case of hamster fetal ovaries in vitro. Furthermore, it ensured the occurrence of meiosis as well as anti-apoptosis on germ cells. BMPR2 was observed to be highly significant for FSH–modulated follicular generation in the pre ovulatory time phase in human GCs. The outcomes of this evaluation of human GCs pointed that significant alterations in expression of the BMP2 proteins was associated with oocytes along with embryo quality [98]. This expression of BMP2 proteins in human CCs has great significance for delineating the molecular pathway that is associated with oocytes along with embryo quality.
Moreover, embryo quality can be anticipated with utilization of apoptotic associated genes. Inhibitors of the apoptosis proteins family (IAP) gene family has survivin as the smallest member, that resides on the chromosome 17q25 which encodes a 142 amino acids proteins 9. The expression of survivin has been documented at the time of embryonic generation as well as in different normal, proliferating adult tissues like skin, endometrium, as well as GCs. In case of pig along with mouse zygotes the survivin accrual took place at chromosomes in the anaphase along with telophase at the time of first cleavage. In case of early mouse embryos, absence of first cleavage causes deletion of survivin action. Survivin participates in controlling production of early embryos in different events associated with cell division [99]. An earlier publication demonstrated that the expression of survivin was based on FSH along with LH gonadotrophins [36] FSH was implicated in the regulation of expression of survivin via the PI3K/AKT pathway, whereas LH was controlled by ERK1/2 pathway. The major functions of cell survival, occur via the manipulation of cell cycle, cell survival, along with cell demise. In the form of a member of IAP, survivin possesses the capacity of repression of apoptosis through hampering of caspase 3 along with 7 [17] that are apoptotic caspases [25].
It was documented that both FSH along with LH caused an escalation of caspase 3 along with 7 [26]. Caspase 3 is the one implicated primarily in bringing about apoptosis, that is inclusive of chromatin condensation, NEBD, plasma membrane blebbing, cells getting unassembled, in addition to generation of apoptotic body [25]. Conversely, caspase 7 has a significant part in the breakdown phase of apoptosis that causes production of reactive oxygen species (ROS), expression of survivin, caspase 3 along with7 might be done as genetic biomarkers in the context of assessment of oocytes along with embryos in case of an ART program implicated in CCs of PCOS subjects [100, 101].
It was documented that AMHR2 along with LIF demonstrated significant variation in expression amongst embryos that were of high besides of low quality. Intriguingly, once both AMHR2 along with LIF concentrations were low, a greater probability of generation of high quality existent. On greater expression of AMHR2 with lower expression of LIF, there was greater probability of generation of low quality embryos, in case of all other probabilities of AMHR2 along with LIF expression, the probability of generation of high along with low quality embryos was equal. As per this publication, a combination of AMHR2 with LIF illustrated a greater anticipation power with regards to detection of quality of embryos [102]. A set of greatly anticipating genes might cause the generation of a model having high anticipation power [102]. Hence the changes in CCs along with oocyte connection in folliculogenesis might have a following, impact on the generation of bad quality of embryos.
Cumulus Cell Biomarkers as well as Pregnancy Results
CCs might work in the form of a noninvasive evaluation of embryonic selection, besides its potential to end in a pregnancy, that are usually thrown away at the time of IVF/ Intracytoplasmic sperm injection (ICSI) events. Markers with regards to fertilization as well as embryo generation would cause escalation of pregnancy probability by selection of ideal oocytes along with embryos. Numerous studies have documented that certain gene whose expression occurred in cumulus cells might act as good biomarkers for embryo and pregnancy outcomes [103]. Significant escalation of implantation in addition to pregnancy outcomes was documented by the performance of cumulus assisted embryo transfer with the utilization of autologous cumulus cells [80]. Hence the cumulus cells possess a significant part in embryonic generation along with provision for enhancement of embryos-uterine adhesion in view of greater physical intimacy along with liberation of cytokines in addition to numerous growth factors requirement in the event of implantation [104].
Lnc RNAs, greater than 200 nucleotide are a newer class of transcripts. These Lnc RNAs, might control necessary pathways that aid in fertilization as well as embryonic generation. In the context of human CCs the relative expression concentrations of AK124742 as well as PSMD6 in the pregnancy group were significantly greater in contrast to non-pregnancy group. AK124742 is a Lnc RNAs that has been recently invented as the natural antisense to PSMD6. The AK124742 as well as PSMD6 expression was associated with embryo quality as well as pregnancy results. Hence the AK124742 as well as PSMD6 as an innovative pair of Lnc RNAs mRNAs gene pair might be believed to be a potential biomarker with regards to embryo selection [105].
Numerous genes that are implicated in mediating the cumulus matrix function as well as cumulus expansion might imply oocyte generational competence as well as pregnancy like VCAN [90], PTX3, as well as PTGS2. VCAN, that is an ECM proteoglycans, results in cross linking of HA in the matrix of expanded CCs, besides stabilization of HA in pericellular matrices [27]. PTX3 gene is implicated in generation of one more ECM protein that cross talks with HA in the matrix of expanded CCs [34, 35, 56]. Furthermore, it has been documented that the anticipation of pregnancy is based on the expression of SDC4 as well as VCAN [95, 106]. In humans, an association was observed amongst the cumulus gene as well as pregnancy results, with the expression of VCAN as well as PTGS2 was significantly greater in CCs from oocytes that resulted in pregnancy with final outcome of live birth rates. VCAN, -GREM1, phosphor fruckto kinase platelets (PFKP) in CCs, were associated with the birth weight in patients who were at 38wks gestation age. Hence In humans CCs, PTGS2, VCAN, PFKP along with GREM1 expression might detect the oocytes, that possess the greater capacity of generation resulting in escalation of implantation rates in addition to greater probability of generation throughout whole pregnancy [96]. Furthermore, it has been demonstrated that the expression concentrations of BMP 15 as well as GDF 9 mRNA in a pregnant group were significantly greater in contrast to non pregnant group [54]. Thus the utilization of BMP 15 as well as GDF 9 mRNA expression can be done for anticipation of clinical pregnancy results [45]. GDF 9 along with its downstream (HAS2 along with PTGS2) might have an association with embryo quality along with positive pregnancy. Escalation of concentrations of BMP 15 as well as GDF 9 expression have been correlated with positive pregnancy along with fertilization rate as well as embryo quality. Of the OLFs, GDF 9 is a significant candidate factor in view of its downstream genes inclusive of HAS2, COX2, GREM1 along with PTX3, were significantly escalated in CCs that were surrounding oocytes of high quality [16].
In humans CCs, the utilization of the gene expression of the can be done as an anticipation marker with regards to successful implantation. Akt1, Bcl1211, along with Shc1in the PI3k / AKT pathway were associated with oocyte maturation along with its capacity. The PI3K / AKT pathway was the crucial controller of function of COCs in addition to controlled the sustenance or stimulation of oocytes along with proliferation differentiation, as well as stress reaction of granulocytes [107]. AKT belongs to a member of serine / threonine kinase family, that is implicated in the control of numerous cellular events, that is inclusive of metabolism, proliferation, cell survival, growth along with angiogenesis. ARHGEF7, CCMND1, E2F1, HRAS as well as SSP1 were further inclusive of regulation of proliferation [99]. HAS2, that gets liberated by the oocyte is believed to be a marker of human pregnancy or live birth [95, 96]. Diaphanous related formin2 (DIAPH2), implicated in spindle dynamics as well as nibrin (NBN) along with alignment of chromosomes was significantly greater in embryos causing implantation along with clinical pregnancies as well as live birth [108]. In case of humans pronuclear fading along with expression of the DIAPH2 gene acted as independent prognostic factors with regards to clinical pregnancies as well as live birth [109]. CAMKID along with EFNB2 have been observed to aid in embryo selection for transfer that possess the maximum capacity of generation of pregnancy [93]. EFNB2 represents a transmembrane protein that belongs to the largest subfamily of receptor protein tyrosine kinases. This protein is implicated in modulatation of generational processes in particular in the nervous system along with erythropoiesis. The expression of EFNB2 in human MGCs was detailed in particular in the luteinization mainly.
CAMKID encodes a member of the Ca/calmodulin - dependent protein kinase 1 subfamily of serine /threonine kinases. Vascular endothelial growth factors (VEGF) causes escalation of EFNB2 in endothelial cells. The maximum expression of EFNB2 in CCs might be a greater expression of VEGF concentration that gets reflected in follicles with better vascularization of the follicles. Hence the greater concentration of VEGF in the follicular fluid had an association with the peri follicular vascularity grade, with a greater vascularization causing an enhancement of fertilization, embryo along with pregnancy [61].
Oxidative stress (OS) as well as hyperandrogenema in addition to disturbed gonadotrophin hormonal liberation can cause induction of apoptosis in MGCs as well as CCs. Significantly lesser apoptotic cells were existent in women that had pregnancy in contrast to those that did not achieve a pregnancy [36]. Apoptosis associated genes are implicated in bad oocytes along with embryo generation in addition to dysfunctional blastocyst production. Association of CCs apoptosis exists with embryo quality along with pregnancy results. Thus these CCs genes might possess adequate reliability as biomarkers for oocytes along with embryo selection along with associated with the bad quality of the subsequent embryos that got generated [36]. Survivin, (that represents an anti-apoptotic gene along with caspase 3 as well as 7(2 of the pro apoptotic genes) have been documented to be implicated in the probability of pregnancy. In case of CCs in Polycystic ovary syndrome (PCOS) women, the Survivin gene expression was lesser, whereas expression of caspase 3 as well as 7 was greater. Hence there might be an inverse association of the rate of granulocyte apoptosis with the generational ability of oocytes. CCs apoptosis might be associated with the pregnancy as well as live birth along with associated embryos with ART therapy. The percentage of apoptotic CCs are intricately associated with the results of Intracytoplasmic sperm injection (ICSI). It can be believed to cause anticipation of pregnancy as well as live birth, with the proportion of the apoptotic CCs are in dependent prognostic factors for these results. For each 1% enhancement of CCs apoptosis, the reduction in clinical along with live birth rates would be11-12% [110].
Oxidative stress (OS) causes stimulation of DNA injury along with starts apoptosis. Its markers are ROS, lipid peroxides [36], total, antioxidant capacity (TAC) as well as 8hydroxy-2’ deoxyguanosines. It has been demonstrated in studies that escalation of ROS resulted in correlation with absence of anti oxidant defences in patients with advanced maternal age, besides changing the function of mitochondrial CCs at the time of generation of oocytes via the free radical production [1]. In view of the significance of mitochondria in yield of energy to the embryos, the utilization of quantitation of mt DNA might be made in the form of implantation biomarkers. The contrast of implanted along with non-implanted groups observed that the average concentration of mt DNA was significantly greater in CCs that were surrounding oocytes along with successful implantation of embryos. With regards to the human embryos that were of equivalent quality, this concentration of mt DNA had an association with the capacity of embryo implantation [36]. Hence the quantitation of mt DNA of CCs represents an oocyte competence biomarkers in addition to aiding in embryo selection with the idea of shifting to obtain a successful pregnancy at the time of IVF events.
Chermula, et al. [111], evaluated the gene transcripts on 1,7,15,30 days of culture for CCs and GC for finding the answer of epithelial origin or not of CCs. Evaluation, in details of the genes that belonged to epithelial cell- associated ontological groups aided them to evaluate a total of 168 genes expressed in CCs (97 genes) and GCs (71 genes) during long-term in vitro culture. Expression changes of the evaluated genes aided the identification of the group of genes: TGFBR3, PTGS2, PRKX, AHI1, and IL11, which had a reduction of maximum of CAV1, and SFRP4 genes, with significant escalation of expression in the group of ANXA3, DKK1, CCND1, STC1, CAV1, and SFRP4 genes. These genes’ expression indicates CCs and GCs epithelialization events along with their epithelial origin. Expression alteration evaluation of genes involved in epithelization events in GCs and CCs during their in vitro culture made it possible to detail the maximum significantly changed 11 genes. Detailed evaluation of gene expression in these two cell populations at separate time intervals corraborates their ovarian surface epithelial origin. Furthermore, some gene expression profiles appear to have tumorigenic properties, suggesting that granulosa cells may play a role in cancerogenesis [111].
Azizollahi, et al. [112], tried to evaluate the superiority of GnRH agonist and/or antagonist protocols in IVF cycles. Thus they evaluated the, gene expression of cumulus cells (CCs) as well as clinical and embryological factors along with contrasted amongst the two groups (GnRH agonistand antagonist) during the IVF cycle. Of the Twenty-one infertile individuals recruited for the study with GnRH agonist (n=10), along with GnRH antagonist (n=11). The defined clinical embryological factors were contrasted amongst the two groups. Expression of BAX, BCL-2, SURVIVIN, ALCAM, and VCAN genes were evaluated in the CCs of the participants with the utilization of the real-time polymerase chain reaction (PCR) method. The average number of cumulus oocyte complex (COC), percentage of metaphase II (MII) oocytes, grade A embryo and clinical factors did not display noticeable alterations, amongst the two groups. BAX gene expression in the CCs of the group treated with GnRH agonist was remarkably higher than those received GnRH antagonist treatment (P<0. 001). The mRNA expression of BCL-2 and ALCM genes were markedly greater in the CCs of patients who underwent antagonist protocol in contrast to the group that received agonist protocol (P<0. 001). Despite no considerable alterations, in the oocyte quality, embryo development, along with clinical outcomes amongst the group treated with GnRH agonist and the one treated with antagonist protocol, the GnRH antagonist protocol was a little more favorable. Nevertheless, further clinical studies with utilization of using molecular evaluation are needed to get insight in this debatable topic [112].
Bisphenol A (BPA) and its analogs, bisphenol S (BPS) and bisphenol F (BPF), might influence fertility by changing oxidative stress pathways. Here, Nguyenposited that bisphenols-stimulated oxidative stress is implicated in causation of reduction of gamete quality. In both female (cumulus-oocyte-complexes—COCs) and male (spermatozoa), oxidative stress was measured by CM- H2DCFDA assay along with and crucial ROS scavengers (SOD1, SOD2, GPX1, GPX4, CAT) quantification at the mRNA and protein concentrations with utilization of qPCR and Western blot (COCs)/immunofluorescence (sperm). Either gamete was treated in five groups: control, vehicle, and 0. 05 mg/mL of BPA, BPS, or BPF. Their outcomes documented escalation of ROS in BPA-treated COCs however reduction in generation in BPS- and BPF-treated spermatozoa. Moreover, both reduction in mRNA and protein expression of SOD2, GPX1, and GPX4 were seen in BPA-treated COCs (p < 0. 05). In sperm, motility (p < 0. 03), however not morphology, was significantly changed by bisphenols. SOD1 mRNA expression was significantly escalated, whereas GPX4 was significantly reduced. These results support BPA’s capacity to change oxidative stress in oocytes along with to a lesser degree, in sperm. Nevertheless, BPS and BPF probably act via separate modes] [113].
Recently Richani D, et al. [114] reviewed the nutrient needs of oocytes & CCs. Glucose is a necessary need for both meiotic along with cytoplasmic maturation of the COC. Glucose is the stimulator of cumulus cell metabolism as well as and is necessary for energy generation, extracellular matrix generation along with provision of pyruvate to the oocyte for ATP generation. Mitochondria are the primary providers of ATP generation within the oocyte. Recent advances in real-time live cell imaging display dynamic fluctuations in ATP need throughout oocyte maturation. Cumulus cells have been demonstrated to play a central part in sustenance of enough oocyte ATP levels by provision of metabolic support via gap junctional connections. Newer understanding have emphasized the significance of oocyte lipid metabolism for oocyte oxidative phosphorylation for ATP generation, meiotic propagation along with generational competence. Within the last decade, different innovative approaches for escalation of the generational competence of oocytes undergoing IVM have been demonstrated, inclusive of modulation of cyclic nucleotides, the supplementation of precursors for the antioxidant glutathione or endogenous maturation mediators like epidermal growth factor-like peptides along with growth differentiation factor 9/bone morphogenetic protein 15. These IVM additives positively alter COC metabolic endpoints that are commonly correlated with oocyte competence. There remain significant challenges in the study of COC metabolism. Owing to the lack of non- invasive or in situ techniques for evaluation of metabolism, maximum investigations till date have used in vitro or ex vivo models. Furthermore, the challenges of estimation of oocyte along with cumulus cell metabolism separately while still existent in a complex has resulted in the frequent use of denuded oocytes, the results from which need interpretation with caution as the oocyte along with cumulus cell chamber are metabolically interdependent, along with oocytes do not naturally exist in a naked state until subsequent to fertilization. There are emerging tools, inclusive of live fluorescence imaging along with photonics probes, that might provide ways to measure the dynamic nature of metabolism in a single oocyte, potentially while in situ [112]. Hence there is a correlation amongst oocyte metabolism and oocyte generational competence. Advancing our insight of basic cellular and biochemical modes regulating oocyte metabolism might isolate new avenues to enhance oocyte quality along with evaluation of generational potential in assisted reproduction [114].
Conclusions
CCs represent separate undifferentiated granulosa cells. This differentiation of CCs get dictated by stimulation by gonadotrophins, (FSH as well as LH), with the lineage of CCs dictated by the effects of OLFs (BMP 15 as well as GDF 9). These 2 oocyte particular growth factors undergo binding with their receptors to cause activation of SMAD2/ SMAD3 pathways, besides result in induction of the cumulus gene expression profile that pointed that repression of LHR along with progesterone receptor expression, avoiding CCs from reaction with the endocrine stimuli of the MGCs. In the meatime the signals from the BMP 15 as well as GDF 9 cause activation of the crucial cumulus particular gene expression, like glycolysis enzymes along with cholesterol generation pathway enzymes that are necessary modulators for healthy oocyte working. CCs continue to be in direct communication with the oocyte via gap junctions contact with the oocyte plasma membrane. The properties of CCs are greater proliferation, greater AMH expression, lesser ability of steroidogenesis, along with lesser expression of LHR. CCs possess the capacity of liberation of HA for COC expansion. At the time of oocyte maturation, the COC expansion is significant for meiotic maturation, via rupture of gap junctions communication with the oocyte plasma membrane, that results in reduction of cAMP in oocyte. Subsequent to LH surge, expansion of COC occurs generation of HA by CCs. This event needs the generation of HA enriched matrix along with factors binding for the stabilization of the matrix. cumulus expansion needs the stimulation with a ligand, either FSH or EGF like peptides along with the activation of MAPK3/1 as well as MAPK14 kinase signaling pathways in CCs along with OLFs. Liberation of growth factors (BMP 15 as well as GDF 9) takes place that results in activation of SMAD2/3 pathways which cause induction of numerous genes that are inclusive of HAS2, COX2, PTGS2, GREM1, StAR along with repression of LHR. Expression of members of the TLR superfamily takes place via CCs which possess the capacity of reaction with particular ligands(that are matrix obtained or pathogen obtained), that results in activation of the innate immune associated genes as well as inflammation. These genes are inclusive of IL6, PTGS2, TNFA, TFAIP6, along with PDCD1. PGE2 that gets generated by IL6, PTGS2 along with TNFα, as well as other cytokines as well as chemokines get liberated from CCs. The break down of polymeric HA by hyaluronidase is believed to results in the production of HA fragments that result in the activation of TLR2 along with TLR4 [115].
Dependent on prior studies, it is thought that CCs might aid in anticipation of oocyte quality, fertilization as well as embryonic generation along with pregnancy. Liberation of numerous factors system takes place at the time of fertilization from the CCs for aiding sperm fertilization. PGE1, PGE2, PGF2, PSSS35, PACAP along with PTX3 get generated by the CCs to aid sperm motion, acrosome reaction as well as fertilization. A reduction in rates of fertilization occurred in humans along with numerous other mammalian species with knockout of these genes. Furthermore, CCs aid in cleavage along with blastocyst generation. Absence of CCs prior to IVF results in reduction of cleavage along with blastocyst generation. Utilization of TRPM7, ITPKA genes is made of for the anticipation of the quality of embryo generation at the cleavage state. Additionally, it is apparent that HAS2, GREM1, GDF9areassociated with the generation of morula along with blastocysts. Conversely, apoptotic along with antiapoptotic genes in CCs like caspase along with survivin Utilization is made for the anticipation of the quality of oocyte along with quality of embryonic generation. Moreover, it has been pointed that numerous genes implicated in modulation of cumulus matrix function along with expansion (VCAN, PTX3, PTGS2) apoptosis modulated genes (survivin, caspase 3 along with7), ROS genes that pointed to those being implicated in the oocyte competence along with the following generation of the embryo along with pregnancy. More recently, greater insight of CCs as well as its function along with its factors to aid in oocyte maturation along with fertilization. Only occasional studies are available in the context of utilization of long non coding RNAs for anticipation of embryo selection as well as pregnancy, with their role being uncertain. Further it is of significance to evaluate the part of lnc RNAs in pregnancy as well. Multiple women try to undergo infertility therapy, CCs factors along with invasive approaches are of significance to evaluate besides their utilization for attaining a successful pregnancy. Here we have tried to provide deeper understanding on the modes of CCs in different stages of generation along with areas that require to get explored in future to further gain insight of the intrafollicular milieu as associated with generational competence.
References
-
Dumesic DA, Meldrum DR, Katz Jaffe MG, Krischer RL, Schoolcraft WB (2015) follicular fluid and cumulus cells are critical for oocyte health. Fertil Steril 103(2): 303- 316.
-
Da Broi M, Giorgi V, Wang F, Keefe D, Albertini D, et al. (2018) Influence of follicular fluid and cumulus cells on oocyte quality. J Assist Reprod Genet 35(5): 735-751.
-
Kong P, Yin M, Tang C, Zhu X, Bukulmez O, et al. (2021) Effects of early cumulus cells removal on treatment outcomes in patients, undergoing invitro fertilization: a retrospective cohort study. Front Endocrinol 12: 669507.
-
Kochar Kaur K, Allahbadia GN, Singh M (2021) A Posit that Luteinizing Hormone Independent Escalation ofProgesterone Might be the Physiological Trigger for the Gonadotrophins Surge in Case of Human Menstrual Cycle. J Gynecol 6(3): 1-11(000221).
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2020) An Update on Future Utilization of Extracellular Vesicles in Evaluation Part of These in Maturation of Gametes, Fertilization along with Embryo Implantation for Escalating the Success of Artificial Reproductive Technology : A Systematic Review. Open Access Journal of Gynecology and Obstetrics 3(12): 10-29.
-
Kochar Kaur K, Allahbadia GN, Singh M (2021) Attempting Utilization of Precision Medicine in Addition to Artificial Intelligence in Reproductive medicine- An Introduction Along with their significance- A Review. Perceptions Reprod Med 4(4): 338-344.
-
Kochar Kaur K, Allahbadia GN, Singh M (2022) An update on the role of Bone morphogenetic protein and Growth Differentiation factor on follicular generation in human menstrual cycle with Clinical impact on ART success & PCO Spathogenesis/therapy-a systematic review. Acta Scientific Women Health 4(2): 58-72.
-
Demiray SB, Goker ENT, Tavmergen E, Yilmaz O, Calimimlioglu N, et al. (2019) Differential gene expression analysis of human cumulus cells. Clin Exp Reprod Med 46(2): 76-86.
-
Huang Z, Wells D (2010) The human oocyte and cumulus cells relationship: newinsights fromthe cumulus cells transcriptome. Mol Hum Reprod 16(10): 715-725.
-
Turatham B, Gao EM, Chian RC (2021) The function of cumulus cells in Oocytes growth and oocyte maturation and in subsequentovulation and fertilization. Cells 10(9): 2292.
-
Kidder GM, Mhawii AA (2002) Gap junctions and ovarian folliculogenesis. Reproduction 123(5): 613-620.
-
Eppig JJ (2001) Oocyte control of ovarian follicular development and function in mammals. Reproduction 122(6): 829-838.
-
Abedini Najabdi A (2015) Elucidation of the biological rolesof Wnt5 signaling infollicles development. Doctoral dissertation. The University of Montreal, Montreal, BC, Canada.
-
Salustri A, Yanagishita M, Underhil CB, Laurent TC, Hascall VC (1992) Localization and synthesis of hyaluronic acid in the cumulus cells and mural granulosa cells of the preovulatory follicle ion. Dev Biol 151(2): 541-551.
-
Zhou L, Kimata K (2001) Cumulus oophorus extracellular matrix:its construction and regulation. Cell Struct F 26(4): 189-196.
-
Diaz Fj, Wigglesworth K, Eppig JJ (2007) Oocytes arerequired for the pre antral granulosa cells to cumulus cells transition from mice. Dev Biol 305(1): 300- 311.
-
Hunzicker Dunn M, Maizels ET (2006) FSH signaling pathway in immature granulosa cells that target regulate ion target gene expression branching out from protein kinase A. Cell Signal 18(9): 1351-1359.
-
Baumgarten SC, Convissar SM, Fierro MA, Winston NJ, Scoccia B, et al. (2014) IGF1R signalingis necessary for FSH induced activation of AKT and differentiation of human cumulus granulosa cells. J Clin Endocrinol Metab 99(8): 2995-3004.
-
Stocco C, Baumgarten SC, Armouty M, FierroMA, Winston NJ, et al. (2017) Genome wide inter actions between FSH and insulin like growth factors in the regulation of human s granulosa cells differentiation. Hum Reprod 32(4): 905-914.
-
Chaffin CL, Vande Voort CA (2013) Follicle growth, ovulation, luteal formation in primates and rodents. :a comparative prospective. Exp Biol Med 238(5): 539-548.
-
Lei Z, Mishra S, Zou W, Xu B, Foltz M, et al (2001) Targeted disruption of Luteinizing hormone/ βhuman chorionic Gonadotropins receptor gene. Mol Endocrinol 15(1): 184-200.
-
Meduri G, Vuhaai-Luuthi M, Jolivet A, Milgrome E (1992) New functional zonations in theovary as shown by Immunohistochemistry of Luteinizing hormone receptor. Endocrinology 131(1): 366-373.
-
Su YQ, Suguira K, Eppig JJ (2009) Mouse Oocyte control of granulosa cells development and function: regulation of cumulus cells metabolism. Semin Reprod Med 27(1): 32-42.
-
Armstrong DT, Xia P, De Gannes G, Tekepaty FR, Khamsi F (1996) Differential effects of insulin like growth factors and follicle stimulating hormone on proliferation and differentiation of bovine cumulus and granulosa cells. Biol Reprod 54(2): 331-338.
-
Mazerbourg S, Klein C, Roh J, Kaive Oja N, Mottershead DG, et al. (2004) Growth Differentiation factor9 signaling is mediated by the type 1 receptor, activin receptor like 5. Mol Endocrinol 18(3): 653-665.
-
Mazerbourg S, Hsueh AJ (2006) Genomic analyses facilitates Identification, of receptor s and signaling pathway for i. Growth Differentiation factor9 and related orphan bone morphogenetic protein/ Growth Differentiation factor ligands. Hum Reprod Update 12(4): 373-383.
-
Peng J, LiQ, WigglesworthK, Rangarajan A, Kattamuri C, et al. (2013) Growth Differentiation factor 9: Bone morphogenetic protein 15 heterodimers are potent regulators of ovarian function. Proc Natl Acad Sci USA 110(8): E776- E785.
-
Sanfins A, Rodriques P, Albertini D (2018) GDF 9 and BMP 15 direct the follicle symphony. J Assist Reprod Genet 35(10): 1741-1750.
-
Sasseville M, Ritter LJ, Nguyen TM, Liu F, Mottershead DG, et al. (2010) Growth Differentiation factor9 signaling requires ERK 1/2activity in mouse granulosa and cumulus cells. J Cell Sci 123(pt 19): 3166-3176.
-
Wigglesworth K, Lee KB, Emori C, Suguira K, Eppig JJ (2015) Transcriptomic diversification of developing cumulus and granulosa cells in Mouse ovarian follicles. Biol Reprod 2015;92(1):23.
-
Borgho T, Povelsse BB, , Andersen CY, BorupR, Humaidan P, et al. (2013) Comparison of gene expression profiles in granulosa and cumulus cells after ovulation induction with either human chorionic Gonadotropins or gonadotropin releasing hormone agonist trigger. Fertil Steril 100(4): 994-1001.
-
Eppig JJ, Pendola FL, Wigglesworth K, Pendola JK (2005) Mouse Oocytes regulate metabolic coopeativity between granulosa cells and Oocytes: amino acids transport. Biol Reprod 73(2): 351-357
-
Hu YC, Wang PH, Yeh S, Wang RS, Xie C, et al. (2004) Subfertility and defective folliclulogenesis, in female mice lacking androgens receptor. Proc Natl Acad Sci USA 101(31): 11209-11214.
-
Gilchrist RB, Ritter LJ, Milymaa S, Kaive Oja N, Dragovic RA, et al. (2006) Molecular basis of oocyte paracrine signaling promotes granulose cells proliferation. J Cell Sci 119(18): 3811-3821.
-
Pangas SA, Matsuk MM (2005) The art and artifact of GDF 9 activity: cumulus expansion and the cumulus expansion enabling factor. Biol Reprod 73(4): 582-585.
-
Taugordeau A, Desquiret Dumas V, Hamel JF, Chupin S, Bouqret L, et al. (2019) The mitochondrial DNA content of cumulus cells may predict embryo implantation. J Assist Reprod Genet 36(2): 223-228.
-
Okudaira Y, Wakai T, Funahashi H (2017) Levels of cyclic AMP and cyclic GMP in porcine oocyte cumulus complexes and cumulus free oocytes derived from small and medium during the first 24h period of in vitro maturation. J Reprod Dev 63(2): 191-197.
-
Van Soom A, Tanghe S, De Pauw L, Maes D, De Gruif A (2002) Function of the cumulus oophorus before and during mammalian fertilization. Reprod Domest Anim 37(3): 144-151.
-
Zhang L, Jiang S, Wozniak PJ, Yang X, Godke RA (1995) Cumulus cell Function during bovine oocyte maturation, fertilization and embryo development, in vitro. Mol Reprod Dev 40(3): 338-44.
-
Turatham B, Sroyraya M (2017) Protein profile involved in mammalian oocyte maturation, and fertilization and early embryogenesis (preimplantation ). Cell Dev Biol 6(3): 1-9.
-
Suguira K, Su YQ, Diaz FJ, Pa ngas SA, Sharma S, et al. (2007) Oocytes derived BMP15 and FGF cooperate to promote glycolysis in cumulus cells. Development 134(14): 2593-2603.
-
Uhde K, van Tol HT, Stout TA, Roelen BA (2018) Metabolomic profiles of bovine cumulus cells and cumulus oocyte complex conditioned medium during maturation in vitro. Sci Rep 8: 9477.
-
XuM, Zeng Y, Chi D, Si L, Qu X, et al. (2018) The dynamic pattern of PLIN3 in pig oocytes and cumulus cells during in vitro maturation. Zygoye 26(1): 40-49.
-
Diaz FJ, Wigglesworth K, Eppig JJ (2007) Oocytes determine cumulus cells lineage in mouse ovarian follicles. J Cell Sci 120(Pt 8): 1330-1340.
-
Tanghe S, Van Soom A, Nauwynck H, Cory n M, De Gruif A (2002) Minireview: Functions of the cumulus oophorus during oocyte maturation, ovulation and fertilization. Mol Reprod Dev 61(3): 414-424.
-
Baena V, Terasaki M (2019) Three dimensional organization of transzonal projection, and other cytoplasmic extensions in the mouse ovarian follicles. Sci Rep 9(1): 1262.
-
Abbasi L, El Hayek S, Carvalho KF, Wang W, Yang Q, et al. (2021) Epidermal Growth factor receptor signaling u ncouples germ cells from the somatic follicular compartment at ovulation. Nat Commun 12: 1438.
-
Zhou CJ, Wu SN, Shen JP, Wang DH, Kong XW, et al. (2016) The beneficial effects of cumulus cells and oocytes cumulus cells gap junctions depends on oocyte maturation and fertilization methods in mice. Peer J 4: e1761.
-
Sasseville M, Gagnon MC, Guillemette C, Sullivan R, Gilchrist RB, et al. (2009) Regulation of gap junctions in porcine cumulus oocyte complexes: contributions of granulosa cells contacts, gonadotrophins and lipid rafts. Mol Endocrinol 23(5): 700-710.
-
Li Y, Li RQ, Ou SB, Zhang NF, Ren L, et al. (2014) Increased GDF 9 and BMP 15 mRNA levels in cumulus granulosa cells correlate with oocyte maturation, fertilization and embryo quality in humans. Reprod Biol Endocrinol 12: 81.
-
Su YQ, Wu Q, O’Briene MJ, Pendola FL, Denegre J N, et al. (2004) Synergistic roles of BMP 15 and GDF 9 in the Development and function of oocyte cumulus cells complexes in mice: genetic evidence of an oocyte granulosa cells regulatory loop. Dev Biol 276(1): 64-73.
-
Hussein TS, Froiland DA, Amato F, Thompson JG, Gilchrist RB (2005) Oocytes prevent cumulus cells apoptosis by maintaining morphogenic paracrine gradient of Bone morphogenetic proteins. J Cell Sci 118(Pt 22): 5257- 5268.
-
Carrette O, Nemade RV, Day AJ, Brickner A, Larsen WJ (2001) TSG-6is concentrated in the extracellular matrix of the mouse Cumulus Oocyte Complexes through hyaluronan and inter alpha inhibitor binding. Biol Reprod 65(1): 301-308.
-
Nagyova E, Salustri A, Nemcova L, Scsukova S, Kalous J, et al. (2020) Versican GI fragments establishes a strongly stabilized interaction with hyaluronan rich expanding matrix during oocyte maturation. Int J Mol Sci 21(7): 2267.
-
Russell D, Robert RL (2007) Molecular mechanisms of ovulation: coordination through cumuluscomplex. Hum Reprod Update 13(3): 289-312.
-
Ploutarchou P, Melo P, Day AJ, Milner CM, Williams SA (2015) Molecular analyses of the cumulus matrix: insights from mice with O’glyca n deficient Oocytes. Reproduction 149(5): 533-543.
-
Elvin JA, Yan M, Matsuk MM (2000) Growth Differentiation factor9 stimulates progesterone synthesis in granulosa cells via prostaglandin EEP Receptor pathway. Proc NatlAcad Sci USA 97(18): 10288-10293.
-
Alam MH, Lee J, Miyano T (2018) GDF 9 and BMP 15 induce development of antrum like structure, by bovine granulosa cells without Oocytes. J Reprod Dev 64(5): 423-431.
-
Nagyova E, Camaioni A, Scsukova S, Miyanorcica A, Prochazka R, et al. (2011) Activation of cumulus cells SMAD 2/3 and Epidermal Growth factor receptor pathway s are involved in porcine Oocytes - cumulus cells expansion and steroidogenesis. Mol Reprod Dev 78(6): 391-402.
-
Diaz FJ, O’Briene MJ, Wigglesworth K, Eppig JJ (2006) The preantral granulosa cells to cumulus cells transition in the mouse ovary: Development of competence to undergo expansion. Dev Biol 299(1): 91-104.
-
Oschner SA, Day AJ, Rugg MS, Breyer RM, Gomer RH, et al. (2003) Disrupted function of Tumor necrosis factor alpha stimulated gene 6 blocks cumulus cells oocyte complex expansion. Endocrinology 144(110): 4376- 4384.
-
Russell D, Salustri A (2006) Extracellular matrix of the cumulus oocyte complexes. Semin reprod med 24(4): 217-227.
-
Jang YJ, Park JL, Moon WJ, Dam PTM, Cho MK, et al. (2015) Cumulus cells expressed type1 interferons induce cumulus expansion in mice. Biol Reprod 92(1): 20.
-
Salustri A, Garlanda C. Hirsch E, De Acetis M, Mcaggno A, et al. (2004) PTX3 plays a key role in the organizations of the cumulus oophorus extracellular matrix and invitro fertilization. Development 131(7): 1577-1586.
-
Nagyova E, Nemcova L, Camaioni A (2021) Cumulus extracellular matrix is an important part of oocyte microenvironment in ovarian follicles: its remodeling and proteolytic degradation. Int J Mol Sci 23(1): 54.
-
Seres KB (2018) Characteristization of a novel Spindle domain in mammalian meiosis. The University of Cambridge, UK, pp: 1-221.
-
Matsuk MM, Burns KH, Vivieros MM, Eppig JJ (2002) Intercellular communication in the mammalian ovary: Oocytes carry the conversation. Science 296(5576): 2178-2180.
-
Richards JS (2005) Ovulation: New factors that prepare the oocyte for fertilization. Mol Cell Endocrinol 234(1- 2): 75-79.
-
Ma Y, Yang W, Ren P, Li X, Jin J, et al. (2021) Lysophosphatidic acid improves Oocytes quality during IVMby activating the ERK1/2 pathway in cumulus cells and Oocytes. Mol Hum Reprod 27(6): gaab032.
-
Ashkenazi H, Cao X, Motola S, Popliker M, Conti M, et al. (2005) Epidermal Growth factor family members: endoge nous mediators of the Ovulatory response. Endocrinology 146(1): 77-84.
-
Bouckenheimer J, Fauque P, Lecellier CH, Bruno C, Commes T, et al. (2018) Differential long non coding RNA expression profiles in human Oocytes and cumulus cells. Sci Rep 8: 2202.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2020) How does Epigenetics Regulate Development of Placenta and Placental Pathologies like PreEclampsia (PE), Intrauterine growth Restriction (IUGR)-With Main emphasis on PE. Advances in Bioengineering and Biomed Res.
-
Blaha M, Prochazka R, , Adamkova K, Nevoral J, Nemcova L (2017) Prostaglandin E2 stimulates the expression of cumulus expansion related genes in pigs. The role of protein kinase B. Prostaglandins Other Lipid Mediatr 130: 38-46.
-
Sugimura S, Yamanouchi T, Palmerini MG, Hashiyada Y, Imai K, et al. (2018) Effect of pre in vitro maturation with cAMP modulators on the acquisition of Oocytes Developmental competence in cattle. J Reprod Dev 64(3): 233-241.
-
Pogrmic-Majkic K, Samardzija G, Stojkov Mimic N, Vukoslavljevic J, Trninic Pjevic A, et al. (2018) Atrazine suppresses FSH induced steroidogenesis and LH dependentexpression of Ovulatory genes through PDE- cAMP signaling pathway in human cumulus granulosa cells. Mol Cell Endocrinol 461: 79-88.
-
Salustri A, Fulop C, Hascall VC, Camaioni A, diGiacomo M (2000) The role of Oocytes in Ovulation. In Ovulation; Springer, Berlin/Heidelberg, Germany, pp: 67-75.
-
Prochazka R, Blaha M, Nemcova L (2012) Signaling pathway s regulating FSH and amphiregulin induced meiotic resumption and cumulus cells expansion in the pig. Reproduction 144(4): 535-546.
-
Keeble S, Firman RC, Sarver BAJ, Clark NL, Simmons LW, et al. (2021) Evolutionary, proteomic, and experimental investigations suggest the extracellular matrix of a cumulus cells mediates fertilization outcomes. Biol Reprod 105(4): 1043-1055.
-
Chen H, Kui C, Chan HC (2013) Ca2+ mobilization in cumulus cells : role in oocyte maturation a acrosome reaction. Cell Calcium 53(1): 68-75.
-
Jaiswal BS, Tur-KspaI, Dor J, Eisenbach M (1999) Human sperm chemotaxis: is progesterone a chemoattractant? Biol Reprod 60(6): 1314-1319.
-
Viggiano J, Herrero M, Cebrakl E, Boquet M, de Gimeno M (1995) Prostaglandin synthesis in cumulus oocyte complexes: effects on in vitro fertilization in mice. Prostaglandin Leukot Essent Fatty Acids 53(4): 261-265.
-
Kulvinder Kochar Kaur, Allahbadia GN, Singh M (2021) Utilizing the Knowledge of Role of Matrix Matrixmetalloproteinases Along with Tissue Inhibitorsof Matrix Metalloproteinasesin Various Gynaecological Disorders with Great Emphasis onPreeclampsia, PCOs Along with Gynaecological Malignancies-A Systematic Review. J Gynecol 6(4): 000223.
-
Brown HM, Dunning KR, Robker RL, Boerboom D, Prichard M, et al. (2010) ADAMTS1cleavage of Versican mediates essential structure remodeling of the ovarian follicle and cumulus - oocyte matrix during Ovulation in mice. Biol Reprod 83(4): 549-557.
-
Luddi A, Gori M, Morocco C, Capaldo A, Pavone B, et al. (2018) Matrix Metalloproteinases and their inhibitors in human cumulus cells and granulosa cells as biomarkers for oocyte quality estimation. Fertil Steril 109(5): 930- 939.e3
-
Li SH, Lin MH, Hwy YM, , Lu CH, YehL Y, et al. (2015) Correlation of cumulus gene expression GJA1, PTRSS35, PTX3 and SERPINE2 with oocyte maturation, fertilization and embryo development. Reprod Biol Endocrinol 13: 93.
-
Tanni I, Aradate T, Matsuda K, Komiya A, Fuse H (2011) PACAP- mediated sperm - cumulus cells interaction promotes fertilization. Reproduction 141(2): 163-171.
-
Briggs DC, Langford Smith AWW, Birchenough HL, Jowitt TA, Kielty CM, et al. (2020) Inter-α- inhibitors heavy chain has an itegrin like 3 D structure mediating immune regulatory activities and stabilization, during Ovulation. J Biol Chem 295(1): 5278-5291.
-
Aftabsavad S, Noormohammadi Z, Moini A, Karimipoor M (2021) Effects of bisphenol A on alterations of ICAM- 1 and HLA-G gene expression and DNA methylation profiles in cumulus cells of infertilie women with poor response to ovarian stimulation. Sci Rep 11: 9595.
-
Fatehi AN, Roelen BAJ, Colenbrander B, schoevers EJ, Gadella BM, et al. (2005) Presence of cumulus cells during invitro fertilization protects the bovine Oocytes against Oxidative stress and improves first cleavage but does not affect further development. Zygote 13(2): 177- 185.
-
Assou S, Haouzi D, De Vos J, Hamamah S (2010) Human cumulus cells as biomarkers for embryo and pregnancy outcomes. Mol Hum Reprod 16(8): 531-538.
-
Kahrman S, Cetinkaya CP, Cetinkaya M, Tufecki MA, Ekmeci CG, et al. (2018) Is there a Correlation between follicle size and gene expression in cumulus cells and is gene expression an indicator of embryo development ? Reprod Biol Endocrinol 16(1): 69.
-
Choi YS, Cho S, Seo SK, Park JH, Kim SH, et al. (2015) Alterations in the intra follicular thiol-redox system in infertilie women with endometriosis. Reproduction 149(2): 155-162.
-
Prabhu AV, Luu W, Sharpe LJ, Brown AJ (2017) Phosphorylation regulates activity of 7-dehydro cholesterol reductase (DHCR7), a terminal enzyme of cholesterol synthesis. J Steroid Biochem Mol Biol 165: 363-368.
-
Wathlet S, Adriaenssens T, Segers I, Verheyen G, Janssens R, et al. (2012) New candidate genes to predict pregnancy outcome in Single embryo transfer cycles when using cumulus cell gene expression. Fertil Steril 98(2): 432-9. e1-4
-
Wathlet S, Adriaenssens T, Segers I, Verheyen G, van de Velde H, et al. (2011) Cumulus cell gene expression predicts better cleavage-stage embryo or blastocyst development and pregnancy for ICSI patients. Hum Reprod 26: 1035-1051.
-
McKenzie L, Pangas SA, Carson S, Kovanci E, Cisneros P, et al. (2004) Human cumulus granulosa cells gene expression:a predictor of fertilization and embryo selection in women underg oing IVF. Hum Reprod 19(12): 2869-2874.
-
Gebhardt KM, Feil DK, Dunning KR, Lane M, Russell DL (2011) Human cumulus cells gene expression as a biomarkers of pregnancy outcome after Single embryo transfer. Fertil Steril 96(1): 47-52.e2.
-
Demiray SB, Yilmaz O, Goker EN, Tavmergen E, Calimlioglu N, et al. (2017) Expression of the Bone morphogenetic protein-2( BMP2) in the human cumulus cells as a biomarker of oocyte s and embryo quality. J Hum Reprod Sci 10(3): 194-200.
-
Pan MH, Ju JQ, Li XH, Xu Y, Wang JD, et al. (2020) Inhibition of survivin induces spindle dis organization, chromosomal misalignment and DNA damage during mouse embryo development. Cell Cycle 19(17): 2148- 2157.
-
Salehi E, Aflatoonia R, Moieni A, Yamini A, Asadi E, et al. (2017) Apoptoti c biomarkers in cumulus cells in relation to embryo quality in Polycystic ovary syndrome. Arch Gynecol Obstet 296(6): 1219-1227.
-
Dekel N, Lawrence TS, Gilula NB, Beers WH (1981) Modulation of cell-cell communication in the cumulus oocyte complexes and the regulation of oocyte maturation by LH. Dev Biol 86(2): 356-362.
-
Devjak R, Burnik PT, Verdenik I, Fon Tacer K, Vrtacnic BF (2016) Embryo quality predictive models based on cumulus cells gene expression. Balkan J Med Genet 19(1): 5-12.
-
Burnik PT, Vrtacnic BF, Maver A, Lovrecic L (2015) Specific gene expression differences in cumulus cells as potential Cumulus biomarkers of pregnancy. Reprod Biomed Online 30(4): 426-433.
-
Parikh FR, Nadkarni SG, Naik NJ, Naik DJ, Uttamchandani SA (2006) Cumulus coculture and cumulus aided embryo transfer increases pregnancy rates in patients undergoing in vitro fertilization. Fertil Steril 86(4): 839-847.
-
Li J, Cao Y, Xu X, Xiang H, Zhang Z, Chen B, et al. (2015) Increased new lnc RNA-mRNA gene pair levels In human cumulus cells correlate with oocyte maturation and embryo development. Reprod Sci 22(8): 1008-1014.
-
Shen Q, Chen M, Zhao X, Liu Y, Ren X, et al. (2020) Versican expression level In cumulus cells is associated with human oocyte developmental competence. Syst Biol Reprod Med 66(3): 176-184.
-
Mohammadi SA, Soleimani M, Deldar H, Salehi M, Soudi S, et al. (2012) Prediction of oocyte developmental competences in ovine using glucose-6 phosphate dehydrogenase (G6DPH) activity determined at retrieval time. J Assist Reprod Genet 29(2): 153-158.
-
Artiini P, Tatone C, Sperduti S, D’Aurora M, Franchi S, et al. (2017) Cumulus cells surrounding oocytes with high developmental competence exhibit down- regulation of phosphoinositol 1,3 kinase/protein kinase B (PI3K/AKT) signalling genes involved in proliferation and survival. Hum Reprod 32(12): 2474-2484.
-
Tabibnejad N, Sheikha MH, Ghasemi N, Fesahat F, Soleimani M, et al. (2019) Association between early embryo morphokinetics plus cumulus cells gene expression and) assisted reproduction outcomes in Polycystic ovary syndrome women. Reprod Biomed Online 38(2): 139-151.
-
Tabibnejad N, Aflatoonian A, Mohammadzadeh I, Soleimani M, Sadeghian NT, et al. (2018) Assessing ICSI outcomes by combining noninvasive indications: early time lapse morphokinetics and apoptosis in associated cumulus cells among women withPolycystic ovary syndrome. Mol Reprod Dev 85(11): 865-874.
-
Chermula B, Kranc W, Celichowski P, Stelmach B, Piotrowska KH, et al. (2022) Cellular processes in human ovarian follicles are regulated by expression profiles of new gene markers- Clinical Approach. J Clin Med 11(1): 73.
-
Azizollahi S, Bagheri M, Haghollahi H, kenazi F, Mohammadi SM (2021) Clinical and molecular effects of GnRH agonists and antagonists on cumulus cells in the invitro fertilization cycle. Int J Fertil Steril 15(3): 202- 209.
-
Nguyen M, Sabry R, Davis OS, Fasetta LA (2022) Effects of BPA, BPS and, BPF, on Oxidative stress and antioxidant enzymes in bovine oocyte and spermatozoa. Genes (Basel) 13(1): 142.
-
Richani D, Dunning KR, Thompson JM, Gilchrist RB (2021) Metaolic codependence of oocytes with cumulus cells: essential role in determining oocyte competence. Hum Reprod Update 27(1): 27-47.
-
475896\ Eppig JJ and nnn bovine porcine granulosa cellsion during cumulus oocyte expansion gene expression Eppig JJ. during receptor implicated Oocytes ovary human quality mediated follicle symptoms embryo Matsuk MM with and tiation.
- Postpartum Maternal Mental Health - A Narrative Review
- Beta HCG in Cervico-Vaginal Secretion as a Predictor of Preterm Delivery
- Successful Management of Mid Trimester Foetal Death with Major Placenta Previa by Expectant Management Followed by Induction of Labour
- To Evaluate the Expression of Egr2 Gene in Term Low Birth Weight Newborns
- Impact of Maternal Obesity on Maternal and Foetal Outcomes: A Prospective Cohort Study from Northern India
- ‘’Benefit of Pulsatile GnRH Therapy in Treatment of Functional Hypothalamic Amenorrhea (FHA) and Congenital Hypogonadotropic Hypogonadism(CHH) in Infertile Patients Over Canonical Gonadotropins with IVF –A Short Communication’’