Production of Ferulic Acid Esterase from Aspergillus Oryzae (NCIM 1212)
The majority of valuable compounds used worldwide are produced by chemical synthesis at very low cost, though consumers prefer to natural compounds because of their health awareness in daily life. Ferulic acid is one of the most valuable compounds having widespread application in various fields such as in baking, cosmetic, beverage and pharma industries, also used as a raw material for the production of vanillin, the most flavouring compound. Ferulic acid esterase is the enzyme that hydrolyzes the ester bond between the ferulic acid and polysaccharides present in plant cell wall and release ferulic acid from agro residues. The production of ferulic acid esterase activity by strains was detected in agar plate assay. The assay involves ethyl ferulate as a sole carbon source in specific media. Of the examined strains Aspergillus oryzae (NCIM 1212) showed the highest level of ferulic acid esterase activity 281 U/mg at the optimum pH and temperature of 5.5 and 35°C. Supplementation of soyabean meal (0.75%) as nitrogen source and sucrose (0.1%) as additional carbon source favored enzyme production.
Introduction
Ferulic acid (FA) is the most abundant phenolic acid in malt and barley [1]. FA along with pcoumaric acid form hemicellulosic polysaccharides in plants. Their ester cross-links are important plant cell wall component that preserve the cell wall structure. This phenolic acid and their derivatives such as ferulic acid (4-hydroxy-3- methoxycinnamic acid) and p-coumaric acid (4- hydroxycinnamic acid) are esterified to the 5’-OH group of arabinose residues which present as side chain of xylans. Synergistic action between main chain degrading enzymes such as endo-β-(1,4)-xylanases and accessory enzyme such as ferulic acid esterase require for the complete degradation of plant cell wall structure which remove FA [1]. FA has wide applications in various industries due to its antioxidant and antimicrobial activity. It also possesses potential therapeutic and physiological properties including anti-inflammatory, anti-diabetic, anti-cancer and anti-ageing effects [2]. Ferulic acid is also involved in the natural production of vanillin (a flavouring compound have application in beverage, foods, perfumes and pharmaceuticals) through its microbial biotransformation.
Potential application of ferulic acid in various industries leads to its release from the walls of agro- residues in large scale. The release of FA from agro- residues involves the use of microbial carboxylic acid esterases enzymes such as ferulic acid esterase (FAE). Numerous reports have been described that FA are released by the enzymes of microbes: Streptomyces olivochromogenes, Penicillium pinophilum, Aspergillus spp. and Chrysosporium lucknowense [3]. Enzymatic method provides environment friendly and very mild method for the release of FA [4]. Ferulic acid esterases (FAE; E.C. 3.1.1.73) are a subclass of carboxylic acid esterase’s that play a key role in degrading the plant cell wall structure by cleaving the ester bond between hydroxycinnamic acids esterified to arabinoxylans and pectins present in plant cell walls [5].
Material and Method
Chemicals- Ethyl ferulate (98%) and ferulic acid (99%) were procured from Sigma Aldrich, HPLC grade. Methanol and ethanol were purchased from HI media. Other chemicals were of analytical grade. Microbial media and medium components were procured from HI media laboratories, Pvt., Ltd., Mumbai, India.
Microbial Strains and Culture Conditions
The strain was obtained from Departmental culture collection of Department of Bio-Engineering, Birla Institute of Technology, Mesra Ranchi. The sub-culturing of strain was done on the slants of potato dextrose agar (PDA). The 6 days old fungal culture from slant was further transferred into the potato dextrose broth (PDB) and again allowed to grow for 6 days at room temperature which was then used for the further inoculation of media.
Method of Cultivation for Ferulic Acid Esterase Production
The Aspergillus oryzae (NCIM 1212) was inoculated in potato dextrose broth for 6 days at 30°C which then transferred into 250 ml flask containing 50 ml of EPM (enzyme production media) having following composition (g/L), (NH4)2SO4 (1.5), KH2PO4 (0.5), Na2HPO4 (1.5), MgSO4.7H2O (0.25 g), CaCl2 (0.05 g), FeCl3 (0.01 g), yeast extract (1.0 g), pH 5.0 and supplemented with 1% ethyl ferulate as C-source dissolved in 1% (w/v) dimethylformamide. Media containing fungal culture ere then incu ated at for -10 days. EPM inoculated with culture was filtered through filter paper after 2-10 days of incubation for the removal of fungal mycelium and spores. Quantification of enzyme activity was done by spectrophotometer and HPLC analysis.
Determination of FAE Activity
FAE activity was quantified by HPLC using ethyl ferulae as substrate. HPLC reaction was prepared by 0.1 ml of substrate (6m M ethyl ferulae) and 0.5 ml of crude enzyme solution, incubated at 37 ºC water bath for 1h. After incubation the reaction was terminated by adding equal volume of stop solution (methanol: acetic acid, 4:1), filtered through a 0.2 µm syringe filter and quantified by HPLC. HPLC analysis was done on a reverse-phase HPLC equipped with a C18 column (Waters XTerra RP18 5 µm, 4.6×150 mm). HPLC setup includes a waters 1525 binary HPLC pump, waters 717 plus auto sampler and waters 2996 photodiode array detector. The analysis of result was done using the Empower Pro software. The 20 µl volume of sample was injected and the elution of product (ferulic acid) and substrate (ethyl ferulae) was done in an isocratic solvent system of water: methanol (40:60) ratio as the mobile phase at the flow rate of 1 ml/min for 10 min and the peak of product and substrate was monitored at 254 and 310 nm. Ferulic acid esterase activity was calculated from the ferulic acid standard curve. One unit of ferulic acid esterase activity was defined as the amount of enzyme (FAE) required releasing 1 µmol of ferulic acid per minute and specific activity was given in units/mg of protein. Total protein concentration was executed by standard Bradford’s method.
Optimization of Production Conditions for FAE
The optimal parameters were further studied to enhance the FAE production by fungal strain Aspergillus oryzae (NCIM 1212). Optimization of media components (C-source and N-source) and environmental factors (days, pH and temperature) was performed to enhance the FAE production.
Optimization of Environmental Factors (Days, pH and Temperature)
Initially the effect of days was optimized by incubating the culture for 2-10 days and quantified the enzyme activity. Effect of pH on the FAE activity was examined by preparing minimal media with pH range of 4-8. In order to examine the effect of temperature with respect to days on FAE production the media was incubated at temperature range of 25- 45 ºC for 2-10 days. Then the HPL analysis and Bradford’s assay of crude extract of enzyme was done to calculate specific activity of enzyme.
Optimization of Media Components (Carbon Source and Nitrogen Source)
In the submerged fermentation by Aspergillus oryzae (NCIM 1212), the effect of different C-source on FAE activity was examined included potato starch, sucrose, dextrose, D-fructose, and lactose at a concentration of 1% (w/v) aong with nitrogen sources by using yeast extract, soyabean meal, yeast extract, peptone, beef extract and casein having 0.1% (w/v) concentration. After the optimization of carbon and nitrogen source giving maximum specific activity of enzyme, the concentration of carbon and nitrogen source were optimized for different concentrations.
Result and Discussion
Optimization of Environmental Factors
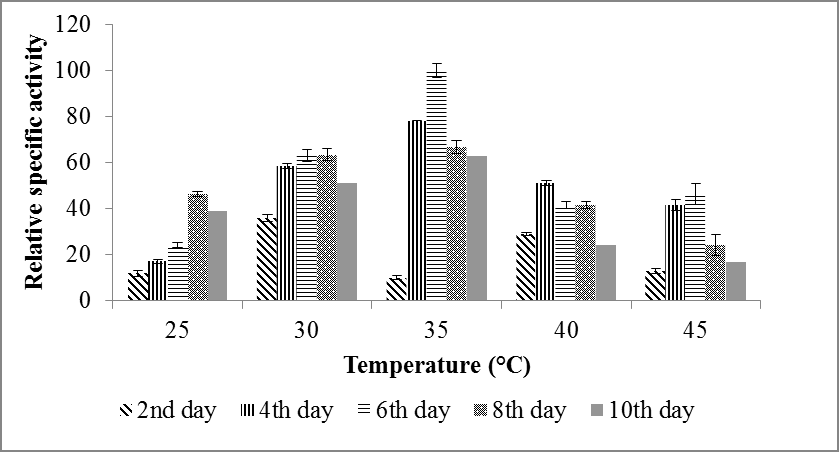
Aspergillus oryzae (NCIM 1212) shows the presence of maximum enzyme activity at 35°C for 6 days incubation. However, comparatively low FAE activity was detected at 2 days, 4 days, 8 days and 10 days of incubation (Figure 1). Microorganisms are indeed susceptible to temperature as it affects the enzyme catalyzed reactions crucial for their survival. The temperature influences the metabolic activity of the cells. There may be low enzyme activity at a temperature below the optimal range and enzyme proteins may denature at higher temperature [6].
Optimization of pH
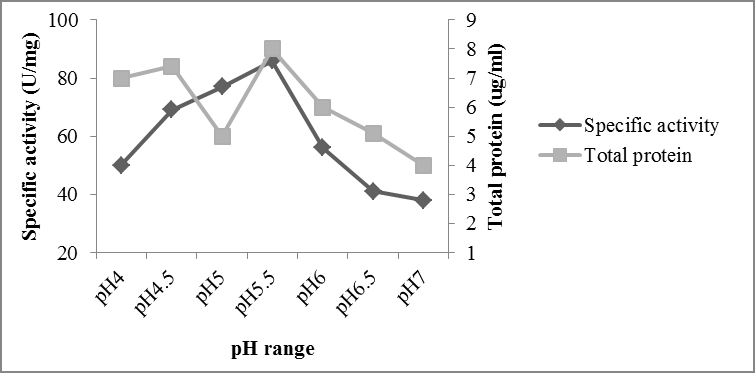
The production of FAE from Aspergillus oryzae (NCIM 1212) was observed in the range of 4.0 to 7.0, with the optimum pH of 5.5 (Figure 2). The optimal pH value obtained is very similar to that obtained for Aspergillus niger strain CFR 1105 (Hegde et al. 2009). There was significant enzyme production in the pH range of 5.0-6.0; however the specific activity of enzyme was very low above pH 7.0. Requirement of slightly acidic condition for the better growth of fungus Aspergillus oryzae (NCIM 1212) incorporated in good production of FAE in pH 5.5 [7].
Optimization of Temperature
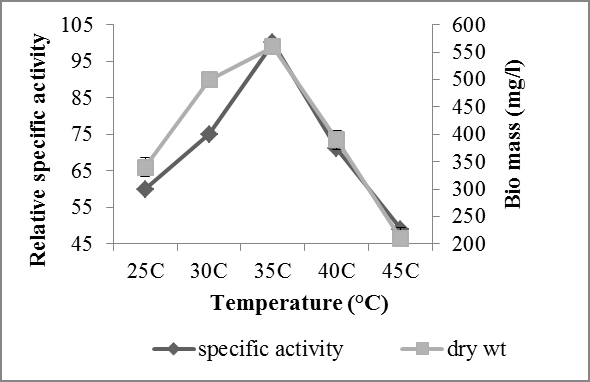
Effect of different temperatures (25°C, 30°C, 35°C, 40°C and 45°C) were investigated for the ferulic acid esterase production. Maximum enzyme production was observed in temperature range of 30-35°C and 35°C was observed as optimum temperature for the production of FAE from Aspergillus oryzae (NCIM 1212) showing specific activity of enzyme 78.6 U/mg of protein (Figure 3). The specific activity of enzyme decreased significantly after temperature of 35°C that may be due to the loss of enzyme activity because of degradation of enzyme’s secondary structure at high temperature. The temperature range observed by Kumar, et al. [8] for the maximum production of FAE was 25-50°C with the optimum temperature of 30°C which shows sufficient similarity with result obtained in our study. They also observed decrease in enzyme production at temperature above 35°C and below 30°C.
Optimization of Carbon Source
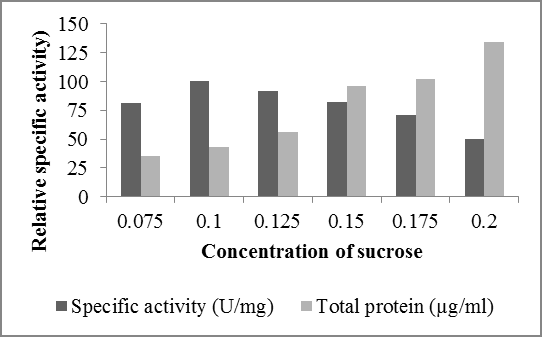
Cellular growth and metabolism of microorganisms require good carbon and nitrogen source which are essential constituents of the fermentation medium. In this study apart from ethyleferulate additional C-source was added to enhance the growth of fungal culture resulting increase the production of FAE. To test the effect of different additional carbon source on FAE production, Sucrose was observed as the most effective carbon source
150 Relative specific activity
100
50
0 Additional carbon source (0.1% w/v) Specific activity (U/mg) Total protein (µg/ml) showing maximum specific activity of enzyme FAE (100 U/mg of protein) produced by Aspergillus oryzae (NCIM 1212). However lactose, fructose, potato starch and dextrose show respectively less specific activity (Figure 4). In the study carried out by Kumar, et al. [8] potato starch (860 U/gds); fructose (848 U/gds) and sucrose (839U/gds) were observed as suitable carbon sources that favouredferulic acid esterase production that shows relevance with this study.
(a) (b) Figure 4: (a) Effect of nitrogen source on Ferulic acid esterase production. (b) Effect of different concentrations of soybean meal on Ferulic acid esterase production.
Optimization of Carbon Source
Among all the nitrogen sources optimized (soybean meal, yeast extract, beef extract, peptone and casein), highest specific activity of Ferulic acid esterase was observed for 0.1 % (w/v) of soybean meal that was
145.24 U/mg of protein followed by beef extract and yeast extract which showed comparatively lesser specific. Casein and peptone showed minimum specific activities (Figure 5).
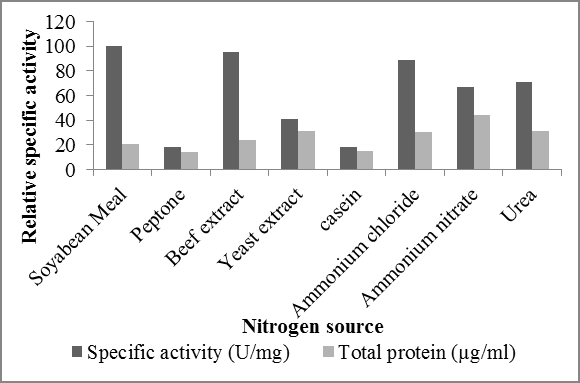
(a) (b) Figure 5: (a) Effect of nitrogen source on Ferulic acid esterase production, (b) Effect of different concentrations of soybean meal on Ferulic acid esterase production.
In earlier study carried out by Shin and Chen, it was found out that complex nitrogen sources such as yeast extract and peptone do not support FAE production as observed in this study too. That shows that yeast extract and peptone are not suitable nitrogen sources for FAE production. We also studied the effect of concentration of soybean meal that was observed as most appropriate C- source for Ferulic acid esterase production, by adding different concentration (0.25-2.0%, w/v) of soybean meal into the minimal media. Maximum specific activity was observed when 0.75% (w/v) soybean meal was used as nitrogen source. However, very low FAE activity was observed when 1.75 and above percentage was used. Thus it shows that high concentration of nitrogen source doesn’t support ferulic acid esterase production as observed by Kumar, et al. [8, 9, 10, 11].
References
-
Dianhui Wu, Guolin C, Xiaomin Li, Bing Li, Jian Lu (2018) Cloning and expression of ferulic acid esterase gene and its effect on wort filterability. Biotechnology Letters 40(4): 711-717.
-
Srinivasan M, Sudheer A, Menon V (2007) Ferulic Acid: Therapeutic potential through its antioxidant property. J Clin Biochem Nutr 40(2): 92-100.
-
Kuhnel S, Pouvreau L, Appeldoorn MM, Hinz SW, Schols HA, et al. (2012) The ferulic acid esterases of Chrysosporium lucknowense C1: Purification, characterization and their potential application in biorefinery. Enzyme microbial Technol 50(1): 77-85.
-
Hasyierah MSN, Zulkali MMD, Syahidah KIK (2008) Ferulic acid from lignocellulosic biomass: review. In: Malaysian University conference on engineering and technology. Putri Brasmara, Perlis, Malaysia.
-
Wong DWS (2006) Feruloyl esterase: a key enzyme in biomass degradation. Appl Biochem Biotechnol 133(2): 87-112.
-
Singh, Surabhi, Nigam VK, Sachan A (2015) Parametric optimization of ferulic acid esterase production from Mucor hiemalis NCIM 837. International journal of Pharmacy and Pharmaceutical Science 7(2): 230-233.
-
Kumar CG, Kamle A, Kamal A (2013) Purification and biochemical characterization of feruloyl esterases from Aspergillus terreus MTCC 11096. Biotechnol Prog 29(4): 924-932.
-
Kumar M, Thakur V, Raghava GP (2008) COPid: composition based protein identification. In Silico Biol 8(2): 121-128.
-
Donaghy J, Kelly PF, Mckay AM (1998) Detection of ferulic acid esterase production by Bacillus spp. and lactobacilli. Appl Microbiol Biotechnol 50(2): 257- 260.
-
Hegde S, Muralikrishna G (2009) Isolation and partial characterization of alkaline feruloylesterases from Aspergillusniger CFR 1105 grown on wheat bran. World Journal of Microbiology and Biotechnology 25(11): 1963–1969.
-
Liyama K, Lam TBL, Stone BA (1994) Covalent Cross- links in the cell wall. Plant Physiol 104(2): 315-320.
- Diversity of Candida sp and Antifungal Susceptibility Patterns in Digestive Candidiasis among People Living with HIV in CHU of Libreville, Gabon
- Vulvovaginal candidiasis: Retrospective study (2019- 2021) at the Centre Hospitalier National de Pikine, Suburban Dakar, Senegal
- Identification of Environmental Fungal Species in Clinical Services of University Hospital of Angre, Abidjan (Cote d’Ivoire)
- New Location of some Gasteroid Basidiomycetes in Western Kazakhstan
- Evaluation of Various Extracellular Enzymes of Ectomycorrhizal Mushrooms
- Morphology and Phylogeny of Lactarius Wallichianae sp. nov and Xerula magnispora sp. nov. from India