Survey of Expression of Aflatoxin Production Regulator Genes (AflR and Nor1) in Aspergillus parasiticus by Euphorbia connata Bioss. and Pimpinella Anisum L.
Aspergillus species are pathogenic and saprophyticus fungi which is responsible new cases of invasive fungal infection across the world. Aflatoxins (produced by Aspergillus parasiticus) are extremely toxic secondary metabolites which contaminate food products. Several investigations have been performed on the removal of aflatoxin using medicinal herbs. In this survey, the effects of Euphorbia connata Bioss. and Pimpinella anisum L. as natural compounds were examined on Aspergillus parasiticus growth, the aflR and Nor1 genes expression. The antifungal susceptibility testing of herbal extracts were performed according to CLSI-M38-A2. Quantitative changes in level of genes expression were evaluated by Real-time PCR method. Results indicated that MIC in the extracts of E. connata Bioss and P. anisum L. against Asp. parasiticus growth were 31.25mg/ml and 125 mg/ ml respectively. The Nor1 gene was detected at a higher effect than the aflR gene in Asp. parasiticus after treatment with E. connata extracts. There were statistically significant differences in Nor1 and aflR gene expressions after treated with E. connata (P=0.001). The Nor1 mRNA expression levels were down regulated by 75% whereas the aflR mRNA expression level was down regulated by 25% (P<0.001) compared to the control group. E. connata extract could be a suitable candidate for control of toxin production by A. parasiticus. However, the P. anisum extract has no effect on gene expression.
Introduction
Aspergillus species (spp.) are pathogenic and saprophyticus fungi which is responsible for annually 200,000 new cases of invasive fungal infection across the world [1, 2]. Rapid death, chronic hepatic and pulmonary disease and developmental disorders in children are the most notable complications of Aspergillus spp. exposure [3]. Prevalence of complications is so considerable that approximately 3 million people are only infected by chronic pulmonary aspergillosis [4]. Even if we diagnose and start treatment plan, the mortality rate of this fungi is 50% and is
100% in missed cases [1].
A significant part of high mortality trait originates from toxins. Aflatoxin in Asp. parasiticus is a life-threatening factor which produced to deteriorate human body structures both directly (inside the body) and indirectly (inside inappropriate food storehouses) [5, 6]. At least 32 enzyme and numerous gene profiles participate in aflatoxin synthesis pathway [7]. There are 2 genes in the route called aflR and Nor1 (aflR). The former gene is essential for aflatoxin production through inhibition of palindromic sequence in gene promotor [8]. The latter one plays the main role in production of an aflatoxin mediator named norsolorinic acid (Figure 1) [9, 10]. Is seems that the invasion of Asp. parasiticus might be controlled by gene suppression.
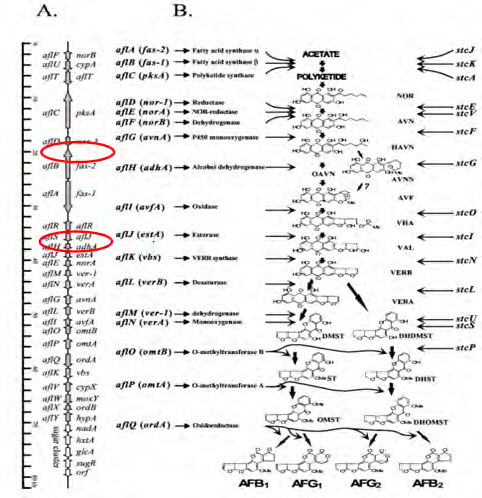
Screening of different medicinal potentials of herbals and natural products is one the enormous interest of scientists to discover new and novel lead compounds that they can solve new health problems such as fungi-toxigenic and so on all over the world [11]. This study is intended to screen two different plants from two families: E. connata and P. anisum.
E. connata from Euphorbiaceae family and in Iran they mainly grow in Kerman, Yazd and Fars provinces [12]. A lot of fractions from Euphorbia have toxic effects and this genus has been used as anti-wart agents in Iranian traditional medicine, diterpenes are major compounds of E. connata [13]. The other plant which is used in this research is a grassy annual plant named Pimpinella anisum from Umbelliferea family and it cultivated in the Mediterranean region, Mexico, Chile and East of Asia (Iran, India) [13, 14]. Various pharmacognostic compounds are determined in both extract and essential oil of this plant and anti-fungal activity is one of the different biological effects of them [15]. The goal of this study is evaluating of the anti-fungal activity of the E. connata and P. anisum extracts on the growth of fungi and aflR and Nor1 genes expression process in Asp. parasiticus.
Material and Methods
Fungal Cell Preparation
Asp. parasiticus strain (ATCC 15517) was incubated for 48h at 30°C on Sabouraud Dextrose Agar media (SDA) (Merck, Germany). Then fresh colony sub cultured on Potato dextrose agar media (PDA) (Merck, Germany) and kept at 30°C for 5-7 days to produce spores. The preparation of the spore suspension was performed according to the previously mentioned procedure. The concentration of spores was calculated using the 0.5Mc Farland turbidity that each well contained 1.5-2 ×108 CFUs/mL [16].
Preparation of E. connata and P. anisum extracts
For collection of plants, fresh stalks, leaves and flowers of E. connata and P. anisum were collected from wild areas of Iran in Kerman and Tehran provinces, after dried in dark place, the aerial parts were milled.
For extraction procedure, both extracts obtained by maceration technique, to do it, 100g of fine powdered parts of E. connata and P. anisum respectively, were immersed in 300ml methanol/water (80/20) and ethanol/water (70/30) and they were mixed on a shaker for 24h at room temperature and this procedure was repeated for more 3 times. After filtration, the extracts were concentrated by rotary evaporator at 40°C and they dried in a dry oven at 40°C and then kept in the refrigerator. To make homogeneous concentrations equal to 500mg/ml, each sample was prepared with sterile water/ Tween 80 (80/20) solvent.
Determination of Minimal Inhibitory Concentration (MIC)
MIC was performed according to CLSI-document M38-A2 [17]. Sterile Roswell Park Memorial Institute (RPMI) (Sigma chemical Co.) buffered to a pH of 7.0 with 0.165M morpholine propanesulfonic acid MOPS) was used.
750mg of E. connata and P. anisum powders were dissolved in 1.5ml sterile diluted water to get a concentration of 500mg/mL, and then diluted to the final concentrations of 250-0.97mg/mL in the mentioned medium according to the CLSI protocol.
Microdilution plates (96 U-shaped wells, Roskilde, Denmark) were used for this purpose. First, 200µL each of extract (500µg/mL) was added to the first well. Then 100µL of RPMI was added to the second well, and then 100µL of the extract in the first well was infused to the second well and carried on until the eighth well. The solution in the eighth well was adjusted to give a final concentration of about 0.97µg/mL of each extract. Then for each extract, 100μL of Asp. parasiticus suspension was cultured with 1mL extractions in 9mL RPMI medium and incubated at 72h/30C. Controls contained RPMI medium with fungal suspension (positive control) and RPMI medium (Negative control).
All tests were performed in triplicate. The micro dilution plates incubated (48h/35°C). MIC determined on the base of lowest concentrations that could prevent any recognizable growth, optical determination of MIC was based on the lowest concentration that made a 100% inhibition for E. connata and P. anisum extracts.
For RNA extraction, mycelia mass was collected and frozen in liquid nitrogen. Total RNA molecules were isolated from logarithmic phase of normal fungal cells and fungal cells treated with extracts according to standard protocol [18]. Spectrophotometer (Bio photometer, Eppendorf, Hamburg, Germany) was used for measuring RNA concentration, then equal concentration of RNA (1μg in 20μL) were used to cDNA synthesis according to the kit protocol (Cinnagen co.) by random hexamer primers. A House keeping gene in this protocol was β-actin gene (ACT1) as normalizer. aflR, Nor1 and ACT1 primers were designed using the primer 3 software and were Synthesize on the basis of published sequence in NCBI (Table 1).
Step-One-Plus real-time PCR system (Applied Biosystems, Foster city, CA) was used for performing real- time PCR. PCR setup and program have been previously [7].
| Gene | Primer Name | Sequence (5’-3’) | PCR Product Size (bp) |
|---|---|---|---|
| Nor1 | Nor1- F | 5’-GTCCAAGCAACAGGCCAAGT-3’ | 66 |
| Nor1- R | 5’-TCG TGCATGTTGGTGATGGT-3’ | ||
| aflR | aflR- F | 5’-CGGAACAGGGACTTCCGGCG-3’ | 200 |
| AflR- R | 5’-GGGTGGCGGGGGACTCTGAT-3’ | ||
| β-actin | β-actin- F | 5’-ACGGTATTGTTTCCAACTGGGACG-3’ | 110 |
| β-actin- R | 5’- TGGAGCTTCGGTCAACAAAACTGG-3’ |
Table 1: Primers for Real-Time PCR.
Results
In current study, for evaluation of antifungal activities of E. connata and P. anisum extracts, broth micro dilution method CLSI document M38-A2 was used.
It is noteworthy, that the inhibitory effect of E. connata on Asp. parasiticus growth was Significant but P. anisum did not reveal any inhibitory effect on the growth. The results demonstrated that the extract of E. connata, inhibited Asp. parasiticus growth at MIC values of 31.25mg/ ml. P. anisum which is inhibited Asp. parasiticus growth at MIC values of 125mg/ml.
Effect of E. connata and P. anisum extracts on aflR and Nor1 genes expression showed in Table 2. Real-time quantitative PCR results showing relative quantification of aflR and Nor1 gene expression levels (calculated according to ΔΔCt-method and represented the rate of aflR gene expression was significantly decreased after treating the Asp. parasiticus with E. connata compare to P. anisum. Here two scenarios are raise: a) It seems the plant extracts via direct blocking of the aflR gene confer to lowering gene expression b) May be over expression of other genes which involved the aflatoxin production play as negative regulatory effects of aflR gene and resulted to lowering aflR gene expression.
The Nor1 gene was detected at a higher effect than the aflR gene in Asp. parasiticus after treatment with E. connata extracts. β-actin gene showed stability in Asp. parasiticus in the presence of E. connata and P. anisum.
There were statistically significant differences in Nor1 and aflR gene expressions after treated with E. connata (P=0.001) (Figure 2). The Nor1 mRNA expression levels were down regulated by 75% whereas the aflR mRNA expression level was down regulated by 25% (P<0.001) compared to the control group (Figure 2).
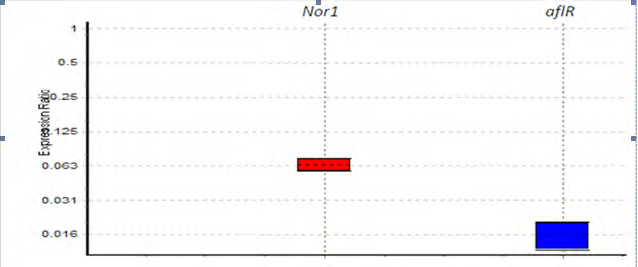
The results of expression of aflR and Nor1 genes did not show any significant difference after treated with P. anisum (Table 2).
| Extract | Gene | Type | Reaction Efficiency | Expression | Std Error | 95% CI | Result |
|---|---|---|---|---|---|---|---|
| E. connata | aflR | TRG | 1 | 0.015 | 0.011-0.020 | 0.011-0.020 | Down expression |
| Nor1 | TRG | 1 | 0.064 | 0.056-0.073 | 0.056-0.073 | Down expression | |
| P. anisum | aflR | TRG | 1 | 0.109 | 0.091-0.121 | 0.084-0.132 | Sample group is not different to control group |
| Nor1 | TRG | 1 | 0.108 | 0.082-0.129 | 0.071-0.151 | Sample group is not different to control group | |
| β-actin | REF | 1 | 1 | - | - | - |
Table 2: Relative expression of aflR and Nor1 genes using Real-Time PCR analysis.
aflR and Nor1 genes expression were normalized to the housekeeping gene, β-actin and analyzed by using REST© software (2008, v.2.0.7). The software uses the comparative Ct method (ΔΔCt) to analyse the data. A sensitive strain (positive control) of Asp. parasiticus was included in each run of the experiment as a positive control.
Discussion
Aflatoxins are the most important mycotoxins produced by Aspergillus species (Asp. flavus, Asp. parasiticus, Asp. sojae, and Asp. oryzae) [19]. Aflatoxin B1 has been identified as the most toxic aflatoxin by the International Agency for Research on Cancer [20].
More than 20 different enzymes are involved in aflatoxin synthesis pathways, and most genes associated with aflatoxin production are located in a 75 kb region of the fungal genome [21, 22, 23].
The aflR gene has been implicated in the regulation of aflatoxin biosynthesis [24, 25]. The biosynthesis is aflatoxin [26]. The aflR gene encodes the AflR protein, which has a zinc-finger motif of the GAL4 type and activates most of the structural genes of the aflatoxin production pathway such as Nor-1 [27]. Research has shown that the absence of the aflR gene or the presence of an abnormal form of this gene blocks the expression of other genes in the aflatoxin production pathway [28, 29]. Aspergillus flavus and Aspergillus parasiticus are able to grow on a variety of substrates and toxin production under humid conditions [30, 31]. High levels of aflatoxin as a carcinogen in the liver can cause acute liver necrosis, cirrhosis or hepatocellular carcinoma. It can also cause bleeding, edema, altered digestion, altered absorption or metabolism of foods, adverse effects on the lungs, heart and kidneys [32, 33]. Because aflatoxins are resistant to normal food processing conditions and are not degraded [31, 34, 35], therefore, an appropriate method is needed to break down aflatoxins while maintaining the quality and quality of the food. Available Aflatoxin degradation methods are costly and reduce the quality of food [36, 37, 38].
Reduction of the expression of aflR considered as an important target. As it has already been mentioned, Anti- fungal activity of methanol extract of E. connata against an aflatoxin-producing Asp. parasiticus have not evaluated yet. Most of scientific studies have been done on the essential oil of P. anisum and few articles showed this plant’s extract biological activities; According to Mehmet Musa Ozcan, et al. anti-fungal activity of essential oil of fruit of P. anisum was evaluated against Alternaria alternata, Asp. niger and Asp. parasiticus. Results of study showed that anise essential oil has anti-fungal activity and can use in food preparations [15]. In the other study, Bluma RV, et al. the application of essential oils of some plants such as: P. anisum and their impact on Aspergillus growth parameters and aflatoxin accumulation were assessed. The result showed that this essential oil has a significant inhibitory effect on growth rate and aflatoxin accumulation [39]. In 2016, another study was done by Alsalhi M, et al. [40] who published their study in 2016, referred to efficacy of anti-microbial and cytotoxicity of nanoparticles using P. anisum seeds. These nanoparticles have effect on selected pathogens: Staphylococcus pyogenes, Acinetobacter baumannii, Klebsiella pneumoniae, Salmonella typhi and Pseudomonas aeruginosa.
The main compounds of genus Euphorbia are a complex mixture of macro-cyclic diterpenoids that they have different profiles of the acylation of their polyol core [41]. According to various applications of terpenes such as antibiotics effects [42], it can be said that likely anti-fungal activity of methanol extract of E. connata has a correlation with these components.
The plant P. anisum contain many secondary bioactive substances such as: terpenes and flavonoids [13], these two main compounds are responsible for the most biological effects of these two genera.
Conclusion
The results by real-Time PCR provided insights into the patterns of the two genes’ mRNA expression levels in exposure with two extracts. In this study, the expression rates of aflR
and Nor1 genes were significantly different (P=0.003 and P=0.001, respectively) after treated with E. connata extract, according to results, 25% and 75% down regulation in the aflR and Nor1 genes were shown, respectively. So this extract could be a suitable candidate for control of toxin production by Asp. parasiticus. The data showed that the difference was not significant between two genes’ mRNA expression (aflR and Nor1) after treated with P. anisum (P=1).
References
-
Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, et al. (2012) Hidden killers: human fungal infections. Sci transl med 4(165): 165.
-
Hua SST, Sarreal SBL, Chang PK, Yu J (2019) Transcriptional Regulation of Aflatoxin Biosynthesis and Conidiation in _Aspergillus_ flavus by Wickerhamomyces anomalus WRL- 076 for Reduction of Aflatoxin Contamination. Toxins 11(2): 81.
-
Groopman JD, Kensler TW, Wild CP (2008) Protective interventions to prevent aflatoxin-induced carcinogenesis in developing countries. Annu Rev Public Health 29: 187-203.
-
Barac A, Kosmidis C, Izquierdo AA, Salzer HJF (2019) Chronic pulmonary aspergillosis update: A year in review. Medical Mycology 57(S2): 104-109.
-
El Darra N, Gambacorta L, Solfrizzo M (2019) Multimycotoxins occurrence in spices and herbs commercialized in Lebanon. Food Control 95: 63-70.
-
Omran GA, El Maali NTB, Ismail MA, Ahmed N, Mostafa M (2019) Differential Hepatic Gene Expression and Antioxidant Activity in Male and Female Rats Induced by Subchronic Aflatoxicosis B1. Mansoura Journal of Forensic Medicine and Clinical Toxicology 27(2): 13-28.
-
Arab Mazar Z, lotfali E, Ghadjari A, Gharehbolagh SA, Mohammadi R (2018) Survey of Expression of Aflatoxin Production Regulator Gene (_aflR_) in _Aspergillus_ Parasiticus by Alpinia Galanga L and Dorema Aucheri. Novelty in Biomedicine 6(1): 29-34.
-
Watson AJ, Fuller LJ, Jeenes DJ, Archer DB (1999) Homologs of Aflatoxin Biosynthesis Genes and Sequence of _aflR_ in _Aspergillus_ oryzae and _Aspergillus_ sojae. Appl Environ Microbiol 65(1): 307-310.
-
Douksouna Y, Masanga J, Nyerere A, Runo S, Ambang Z (2019) Towards Managing and Controlling Aflatoxin Producers within _Aspergillus_ Species in Infested Rice Grains Collected from Local Markets in Kenya. Toxins 11(9): 544.
-
Salehi P, Sonboli A, Eftekhar F, Nejad Ebrahimi S, Yousefzadi M (2005) Essential Oil Composition, Antibacterial and Antioxidant Activity of the Oil and Various Extracts of _Ziziphora clinopodioides_ subsp. rigida BOISS. RECH. f. from Iran. Biol Pharm Bull 28(10): 1892- 1896.
-
Kaushik K, Agarwal S (2019) The Role of Herbal Antifungal Agents for the Management of Fungal Diseases: A Systematic Review. Asian Journal of Pharmaceutical and Clinical Research 12(7): 34-40.
-
Erasmus LJC (2014) Impact of various boiling intervals on the antimicrobial efficacy and phytochemical profile of selected crude aqueous plant extracts, used by Bapedi Traditional Healers in the treatment of sexually transmitted infections. University of Limpopo, South Africa, pp: 1-215.
-
Kosalec I, Pepeljnjak S, Kustrak D (2005) Antifungal activity of fluid extract and essential oil from anise fruits (_Pimpinella anisum_ L., Apiaceae). Acta Pharm 55(4): 377-385.
-
Besharati Seidani A, Jabbari A, Yamini Y (2005) Headspace solvent microextraction: a very rapid method for identification of volatile components of Iranian _Pimpinella anisum_ seed. Analytica Chimica Acta 530(1): 155-161.
-
Ozcan MM, Chalchat JC (2006) Chemical composition and antifungal effect of anise (_Pimpinella anisum_ L.) fruit oil at ripening stage. Annals of Microbiology 56(4): 353- 358.
-
Mohseni R, Noorbakhsh F, Moazeni M, Omran AN, Rezaie S (2014) Antitoxin Characteristic of Licorice Extract: The Inhibitory Effect on Aflatoxin Production in _Aspergillus_ _parasiticus_. Journal of food safety 34(2): 119-125.
-
Rex JH, Alexander BD, Skaggs BA, Andes D, Chaturvedi V, et al. (2008) Reference method for broth dilution antifungal susceptibility testing of yeasts. Approved Standard-Third Edition. Clinical and Laboratory standards Institute 28(14): 1-7.
-
Noorbakhsh F, Lotfali E, Ghajari A, Ansari S, Mohammadi R, et al. (2017) The Effect of _Chenopodium album_ and _Apium nodiflorum_ on the Expression of the Regulatory Gene (_AflR_) that Produces Aflatoxin in _Aspergillus_ parasiticus. Herbal Medicines Journal 2(2): 60-65.
-
Hudler GW (2000) Magical mushrooms, mischievous molds. Princeton University Press, USA, pp: 248.
-
Abdel Hadi AM, Caley DP, Carter DRF, Magan N (2011) Control of aflatoxin production of _Aspergillus flavus_ and _Aspergillus parasiticus_ using RNA silencing technology by targeting _aflD_ (nor-1) gene. Toxins 3(6): 647-659.
-
Kujawa M (1993) Some naturally occurring substances: food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans 56. World Health Organization 38(3): 351.
-
Trail F, Mahanti N, Linz J (1995) Molecular biology of aflatoxin biosynthesis. Microbiology 141(4): 755-765.
-
Yuan GF, Liu CS, Chen CC (1995) Differentiation of _Aspergillus parasiticus_ from _Aspergillus sojae_ by random amplification of polymorphic DNA. Appl Environ Microbiol 61(6): 2384-2387.
-
Woloshuk C, Prieto R (1998) Genetic organization and function of the aflatoxin B1 biosynthetic genes. FEMS microbiol lett 160(2): 169-176.
-
Chang PK, Cary JW, Bhatnagar D, Cleveland TE, Bennett JW, et al. (1993) Cloning of the _Aspergillus_ parasiticus apa-2 gene associated with the regulation of aflatoxin biosynthesis. Appl Environ Microbiol 59(10): 3273- 3279.
-
Payne GA, Nystrom GJ, Bhatnagar D, Cleveland TE, Woloshuk CP (1993) Cloning of the afl-2 gene involved in aflatoxin biosynthesis from _Aspergillus_ flavus. Appl Environ Microbiol 59(1): 156-162.
-
Woloshuk CP, Foutz KR, Brewer JF, Bhatnagar D, Cleveland TE (1994) Molecular characterization of _aflR_, a regulatory locus for aflatoxin biosynthesis. Appl Environ Microbiol 60(7): 2408-2414.
-
Abdel Hadi A, Carter D, Magan N (2011) Discrimination between aflatoxigenic and non-aflatoxigenic _Aspergillus_ section Flavi strains from Egyptian peanuts using molecular and analytical techniques. World Mycotoxin Journal 4(1): 69-77.
-
Wang BS, Zhao Q, Wang YZ (2011) The uncertainty of assessing aflatoxin B1-producing ability using _aflR_ gene in _Aspergillus_ species. African Journal of Microbiology Research 5(31): 5603-5606.
-
Georgianna DR, Payne GA (2009) Genetic regulation of aflatoxin biosynthesis: from gene to genome. Fungal Genet Biol 46(2): 113-125.
-
Jahanshiri Z, Ghahfarokhi SM, Allameh A, Abyaneh RM (2012) Effect of Curcumin on _Aspergillus parasiticus_ growth and expression of major genes involved in the early and late stages of aflatoxin biosynthesis. Iran J public health 41(6): 72-79.
-
Yu J, Ehrlich KC (2011) Aflatoxin biosynthetic pathway and pathway genes. INTECH Open Access Publisher.
-
Cary JW, Ehrlich KC, Wright M, Chang PK, Bhatnagar D (2000) Generation of _aflR_ disruption mutants of _Aspergillus parasiticus_. Applied Microbiology and Biotechnology 53(6): 680-684.
-
Azab RM, Wail TM, Al Rahman HAM, Al Majd AMK, Al Agrab HM, et al. (2005) Detection and estimation of aflatoxin B1 in feeds and its biodegradation by bacteria and fungi. Egyptian Journal of Natural Toxins 2(1): 39- 54.
-
Cotty P (1989) Aflatoxin and sclerotial production by _Aspergillus flavus_: influence of pH [1988]. Food and Agriculture Organization of the United Nations 78(9): 1250-1253.
-
Abbas HK (2005) Aflatoxin and food safety. 1st (Edn.), Taylor & Francis Group, CRC Press, pp: 616.
-
Munoz R, Arena ME, Silva J, Gonzalez SN (2010) Inhibition of mycotoxin-producing _Aspergillus nomius_ VSC 23 by lactic acid bacteria and _Saccharomyces cerevisiae_. Brazilian Journal of Microbiology 41(4): 1019-1026.
-
Chang PK, Ehrlich KC, Yu J, Bhatnagar D, Cleveland TE (1995) Increased expression of _Aspergillus parasiticus_ _aflR_, encoding a sequence-specific DNA-binding protein, relieves nitrate inhibition of aflatoxin biosynthesis. Appl Environ Microbiol 61(6): 2372-2377.
-
Bluma RV, Etcheverry MG (2008) Application of essential oils in maize grain: Impact on _Aspergillus_ section _Flavi_ growth parameters and aflatoxin accumulation. Food Microbiology 25(2): 324-334.
-
AlSalhi MS, Devanesan S, Alfuraydi AA, Vishnubalaji R, Munusamy MA, et al. (2016) Green synthesis of silver nanoparticles using _Pimpinella anisum_ seeds: antimicrobial activity and cytotoxicity on human neonatal skin stromal cells and colon cancer cells. Int J Nanomedicine 11: 4439-4449.
-
Shadi S, Saeidi H, Ghanadian M, Rahimnejad MR, Aghaei M, et al. (2015) New macrocyclic diterpenes from _Euphorbia connata_ Boiss. with cytotoxic activities on human breast cancer cell lines. Nat Prod Res 29(7): 607- 614.
-
Rajeh MAB, Zuraini Z, Sasidharan S, Latha LY, Amutha S (2010) Assessment of Euphorbia hirta L. leaf, flower, stem and root extracts for their antibacterial and antifungal activity and brine shrimp lethality. Molecules 15(9): 6008-6018.
- Diversity of Candida sp and Antifungal Susceptibility Patterns in Digestive Candidiasis among People Living with HIV in CHU of Libreville, Gabon
- Vulvovaginal candidiasis: Retrospective study (2019- 2021) at the Centre Hospitalier National de Pikine, Suburban Dakar, Senegal
- Identification of Environmental Fungal Species in Clinical Services of University Hospital of Angre, Abidjan (Cote d’Ivoire)
- New Location of some Gasteroid Basidiomycetes in Western Kazakhstan
- Evaluation of Various Extracellular Enzymes of Ectomycorrhizal Mushrooms
- Morphology and Phylogeny of Lactarius Wallichianae sp. nov and Xerula magnispora sp. nov. from India