The Study of Cultural and Morphological Characteristics of Colletotrichum gloeosporioides causing anthracnose on Barringtonia Edulis
This study was conducted to investigate a characteristic fungal disease, causing leaf anthracnose on B. edulis. A symptom progression from an angular-asymmetrical yellow discoloration to an ulcer-like necrosis caused death to the whole leaf. Colony pigmentations observed among the six isolates ranged from grey to white with slightly raised aerial mycelium to dense cottony mycelium. Most of the isolates showed salmon to bright orange spore-masses that were arranged in concentric rings, and with aging these orange spore-masses evenly distributed towards the culture peripheral. The conidial shapes of all of the isolates were straight and cylindrical with the average conidia length and width, ranged from 55.6-62.8 and 16.8- 24μm. Together with differential tests using fungicide and a temperature regime revealed characteristics corresponding to Colletotrichum gloeosporioides.
Peter Buyoyu1,2* and Macquin Kilagi Maino3
Keywords: Cutnut; Colletotrichum gloeosporioides; anthracnose
Introduction
Cutnut belongs to the family Lecythidaceae, which produces edible nuts consumed as snacks by Melanesians, and is commonly distributed in some parts of Papua New Guinea (PNG), Solomon Islands and Vanuatu Bourke RM, et al. [1]. Cutnut is an income earner for Melanesians, where the nut is sold in the domestic markets for cash. The commercialization of edible Barringtonia spp. is not well established in PNG and Solomon Islands, while Vanuatu is the only country in the Western Pacific that engages in the commercial production and export of edible Barringtonia nuts Wah LC, et al. [2]. Apart from being an important indigenous edible nut and a potential commercial crop [2, 3], B. edulis has other benefits like medicine for minor ailments [4, 5]. Many farmers in Temota Islands of the Solomon Islands used cutnut for trelliswork mainly for betel piper (Piper betel); while in Kolombangara Islands cutnut is planted to indicate traditional land boundaries [6]. There are numerous limiting factors to the commercial production of B. edulis nuts despite growing interests in the commercial production of indigenous nuts in the Pacific, particularly, in the Melanesian region. The commercial production constraints for B. edulis nuts are mostly commodity-related, and therefore, priorities for research and development are mostly focused on processing, marketing and agronomy issues [7]. Although, crop protection was not considered as a priority for research and development in this quest for commercial production of indigenous nuts in the Pacific, this study would encourage more research to focus on the diseases of indigenous nuts for sustainable production.
Several foliar diseases have been reported, such as leaf spots caused by Pseudocercospora barringtoniicola, Pseudocercospora barringtoniigena and Cercospora barringtonia on inedible Barringtonia spp. especially B. asiatica [8], B. speciosa [8, 9], B. acutangula [8] and B. yunnanesis [8]. The ascomycetes, Phyllachora barringtoniicola and P. naqsii have also been reported causing leaf spots on inedible Barringtonia spp. Hyde KD, et al. [10] Interestingly, similar leaf spot symptom characteristics observed on B. asiatica were found to be caused by Deniquelata barringtoniae gen. et sp. nov. [11]. This is expected as many pathogens may induce similar symptoms and, therefore, there is the need to verify the causal agent in every instance. Notably, C. gloeosporioides, a known pathogen that causes anthracnose in several other plant species also exists asymptomatically as an endophyte in Barringtonia species [12]. Buyoyu P, et al. [13] reported the first occurrence of C. gloeosporioides species complex on B. edulis causing anthracnose on the leaves using molecular methods.
Materials and Methods
Study Site
The experiments were conducted at UNITECH Biotechnology Centre (UBC), Taraka Campus, Lae, PNG, in 2014 and 2015. Infected leaves were obtained from a cutnut plant growing at the staff residential area in the Taraka Campus of UNITECH, where symptomatic diseased leaves were initially observed.
Observation on Symptom Development
Field observations on symptom development were carried out over a period of 12 weeks in 2014. The infection progression from an initial stage of yellowing through to fully developed, ulcer-like infection (anthracnose) was routinely observed. Six developmental stages were subjectively described and photographed using a digital camera (Nikon) both under a dissecting microscope in the laboratory and out in the field.
Pathogen Isolation and Culture Maintenance
Infected B. edulis leaves were detached from the plant, placed in zip-locked bags and were brought to the laboratory for the study. The leaf samples were washed under running tap water before excising approximately 1-cm2 explants with equal proportions of infected and uninfected areas. The excised pieces were surface-sterilized with 0.5% sodium hypochlorite for 5 min, and then rinsed three times with double distilled water [14]. Potato dextrose agar (PDA) was amended with streptomycin sulphate (100 mg L-1) and used to inhibit bacterial growth. Pure cultures were obtained by slightly agitating seven-day-old cultures over fresh PDA to release the spores under laminar airflow, and the spore- inoculated cultures were incubated in the dark at 25°C. After 24 to 48 h, the germinating spores were identified under dissecting light microscope and then transferred to PDA media in 90-mm petri dishes using a sterile glass needle [15] and maintained as pure cultures in PDA slants.
Cultural and Morphological Examination
Observations based on colony colour and texture from pure cultures revealed six isolates. The growth rates for each of these isolates were measured from 10 culture-plates. Besides, conidia shape was observed from slides that were prepared using modified slide culture technique [16]. Under the aseptic condition of a Laminar Air Flow, an agar block (4.5 cm3) was placed on a sterile slide, and inoculated with hyphal-tips. A sterile coverslip was then aseptically placed over the inoculated agar block. Each of these slide cultures were then placed in petri dishes and incubated at 25°C for four days.
Furthermore, a modified fungicide test using PDA amended with Cu (OH)2 Martínez EP, et al. [17] was used as a selective medium to differentiate C. gloeosporioides and C. acutatum based on colony colour and growth rate. In preparing the PDA-Cu (OH)2 media, 50 mg L-1 of Cu [OH]2 was added to the PDA before dispensing into petri dishes. The cultures for the fungicide tests were then incubated at 25°C, and colony colors were described after seven days [17]. The six preliminarily described isolates were then subjected to temperature test at 35°C to differentiate the isolates into C. gloeosporioides and C. acutatum, as this thermal condition is restrictive to the growth of C. acutatum [18].
Pathogenicity test for the six isolates was carried out on ten detached leaves of B. edulis. Fifth fully opened leaves from meristem were detached and transported to the laboratory in zip-locked bags. The leaves were surface-sterilised with 70% ethanol, rinsed with double distilled water and then dried with sterile cheese cloth before inoculation. The leaves were inoculated with 0.5 mL of 105 spores/mL suspension, and incubated under sterile moisturized containers and incubated for 14 days [13].
Results
Disease Symptoms, Cultural and Morphological Characteristics
Disease symptom development was observed to progress from angular-asymmetrical yellow discoloration to irregular-sub-circular spots (Figure 1). These spots became sunken, forming ulcer-like necrosis as the spots increased in size. These enlarged spots then became confluent creating large necrotic patches with irregular margins. As these large patches became confluent, the proportion of necrosis on the leaf tissue increased in size, eventually killing the whole leaf.
![Figure 1: Anthracnose disease on Barringtonia edulis showing disease developmental stages characterised by angular- asymmetrical discolorations (yellowing), which are mainly found in the older leaves. a) Discolorations then advanced to irregular-subcircular leaf spots, which also are visible on the underside. b) Necrosis increased in size becoming sunken, ulcer-like large spots, which are characteristic of anthracnose disease. c) Individual spots confluent, forming large irregular-bordered patches. d) Advanced stages of the infection with zonated imprints of anthracnose and; e) Leaf apex completely affected by anthracnose f) Source: Buyoyu P, et al. [13].](/fulltextimages/8444/fig_1.png)
Figure 1: Anthracnose disease on Barringtonia edulis showing disease developmental stages characterised by angular- asymmetrical discolorations (yellowing), which are mainly found in the older leaves. a) Discolorations then advanced to irregular-subcircular leaf spots, which also are visible on the underside. b) Necrosis increased in size becoming sunken, ulcer-like large spots, which are characteristic of anthracnose disease. c) Individual spots confluent, forming large irregular-bordered patches. d) Advanced stages of the infection with zonated imprints of anthracnose and; e) Leaf apex completely affected by anthracnose f) Source: Buyoyu P, et al. [13].
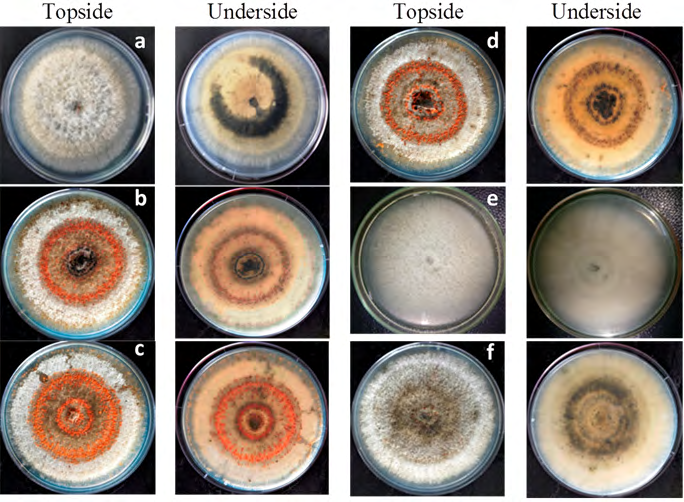
The main colony pigmentations observed among the six isolates were grey, greyish white to white with slightly raised aerial mycelium (BE02, BE03, BE04 and BE05) to dense cottony mycelium (BE01 and BE07). In most of the isolates, salmon to bright orange spore-masses were arranged in concentric rings, and with aging these orange spore-masses evenly distributed towards the culture periphery. Observing from the underside of the culture plates, there were concentric rings of arcevuli observed in isolates BE01, BE02 and BE04. Morphologically, the conidial shapes of all the isolates were straight and cylindrical (Table 1). The average conidia length and width, ranged from 55.6-62.8 and 16.8-24 µm (N=30), respectively.
| Isolate | aShape | Conidia size (µm)* | Growth rate (mm.d-1)** |
|---|---|---|---|
| Length x Width min-(mean)-max | |||
| BE 01 | St, C | 12-(13.5)-16 x 3-(4)-6 | 5.86 ± 0.16 bc |
| BE 02 | St, C | 12-(15)-18 x 3-(5.3)-6 | 5.86 ± 0.08 bc |
| BE 03 | St, C, OA | 12-(15.6)-18 x 3-(4.8)-6 | 5.38 ± 0.17 ab |
| BE 04 | St, C | 12-(12)-18 x 3-(6)-6 | 5.10 ± 0.05 a |
| BE 05 | St, C | 12-(13.9)-15 x 3-(4.2)-6 | 6.05 ± 0.17 c |
| BE 07 | St, C | 12-(13.6)-15 x 3-(4.2)-6 | 6.05 ± 0.17 c |
Table 1: Morphometric measurements of conidia and mycelial growth rates of the fungal isolates.
aShape of Conidia: St = Straight, C = Cylindrical, OA = Obtuse apex; * n = 30; **n = 5, same letters = means have no significant difference (p<0.05). Table 1: Morphometric measurements of conidia and mycelial growth rates of the fungal isolates.
The six isolates tested on PDA amended with Cu (OH)2 showed variable cultural characteristic. Three of isolates, (namely; BE01, BE05 and BE07) exhibited fast growing habit with olive coloured colonies, one (BE02) in salmon pigmented colony, and two (BE03 and BE04) having slow growing habit with greyish colonies (Table 2). The temperature test, on the other hand, showed five isolates namely, BE01, BE03, BE04, BE05 and BE07 (Figures 2a- f), as having dark grey pigmented colonies with olive undersides, while isolate BE05 (Figure 2e) expressed a light grey topside with a jasmine-coloured underside. Contrarily, isolate BE02 (Figure 2b) showed dark black coloured colony with salmon colony peripherals. Moreover, symptom expression on the artificially infected detached leaves, and the cultural and morphological characteristics of the re isolates on PDA media were consistent with that of the initial isolates.
| Isolate | FGO (Fast growing olive)a | FGS (Fast growing salmon)a | SGG (Slow growing grey)a | SGO (Slow growing orange)a | Classification of isolates |
|---|---|---|---|---|---|
| BE01 | + | - | - | - | C. gloeosporioides |
| BE02 | - | + | - | - | C. gloeosporioides |
| BE03 | - | - | + | - | C. gloeosporioides |
| BE04 | - | - | + | - | C. gloeosporioides |
| BE05 | + | - | - | - | C. gloeosporioides |
| BE07 | + | - | - | - | C. gloeosporioides |
Table 2: Differentiation of Colletotrichum isolates based on colony colour on PDA amended with Copper hydroxide.
a(-) = no match; (+) = match Table 2: Differentiation of Colletotrichum isolates based on colony colour on PDA amended with Copper hydroxide.
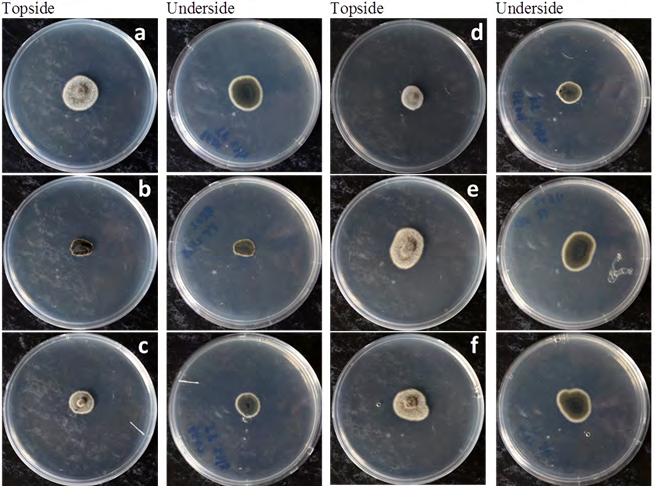
Furthermore, the temperature test indicated that the growth of isolates was not restricted when incubated at 35°C (Figure 3). The results of temperature test revealed the tolerance of isolates to the thermal condition that is restrictive to the growth of C. acutatum.
Discussion
Systematic observations in the current study were consistent with symptom development reported under controlled conditions. Disease symptom development progressed from different stages, which eventually reached advanced infection with characteristics of anthracnose. These symptoms were similar to anthracnose symptoms caused by Colletotrichum spp. on artificially inoculated raspberry, capsicum and tomato fruits [19]. Similar anthracnose symptoms were also observed on the leaves of yam (D. alata) caused by C. gloeosporioides complex in different parts of Nigeria based on in vitro diagnosis of cultural and morphological characteristics [20]. Use of cultural and morphological characteristics has been reported useful in characterising other Colletotrichum species [21].
Cultural profiles on solid PDA media, such as colony colour, colony texture and colony growth habit of all isolates indicated characteristics similar to cultural characteristics of Colletotrichum species. These variations in colony pigmentation observed from the current study agree with several studies reported on the colony colour of C. gloeosporioides [19, 22, 23]. The isolates with salmon pigmented colonies (Figures 2b-d) are also consistent with C. gloeosporioides isolates causing foliar anthracnose on Atractylodes ovate [24]. Similar observations of colony characteristics for Colletotrichum spp. were also observed in isolates from forest nurseries in Peninsula Malaysia Zakaria M, et al. [25] and certain herbaceous plants of agricultural and ornamental importance in Thailand [22]. Mycelial growth rates of all isolates in the current study were faster than the growth rates of Colletotrichum isolates reported by Zakaria M, et al. [25]. This could be due to the variation in culture conditions since the growth rate of many Colletotrichum spp. is affected by incubation conditions, and the optimal growth temperature is isolate-dependent [26].
Colletotrichum gloeosporioides and C. acutatum are the two major causes of anthracnose, and distinguishing the two species using cultural profiles alone is erratic, thus a better differentiating method is needed. Fungicide sensitivity test using PDA amended with Cu(OH)2 has successfully separated the isolates into respective morphotype groupings based on colony pigmentation and mycelial growth habit (Table 2), where all isolates exhibited cultural characteristics that depicted C. gloeosporioides [17]. The PDA amended with Cu(OH)2 has effectively separated the isolates into the following morphotypes; fast growing olive (FGO), slow growing salmon (SGS) and slow growing grey (SGG). These results are similar to C. gloeosporioides isolates from yam anthracnose Abang MM, et al. [27]. Furthermore, cultural incubation at 35°C did not restrict the mycelial growth in all isolates indicating that all isolates to be C. gloeosporioides as this thermal condition is restrictive for the growth of C. acutatum [18].
The conidia morphology of all isolates were mainly straight and cylindrical except isolate BE03, which showed straight, cylindrical conidia with tapering ends. Morphometric characteristics of the conidia from this study are alike with the conidia morphometrics reported in previous studies [22, 23, 28]. Conventional diagnostic techniques using morphological characteristics alone to identify Colletotrichum spp. Sutton BC, et al. [29] are often unreliable due to the ability of certain Colletotrichum spp. to produce secondary conidial structures [30]. These secondary conidial structures, in many cases, have totally different morphometrics compared to the primary conidial morphometrics, and this creates ambiguity leading to misidentifications of Colletotrichum spp. [30, 31].
Interestingly, this species is known for its cross-infection ability [14, 32]. Given that B. edulis is normally cultivated with other food crops in home-gardens, risk for cross-infection by C. gloeosporioides from infected B. edulis to other local food crops known to suffer from this particular pathogen. The findings from the current study provide another indication that C. gloeosporioides has a wider host range. C. gloeosporioides has been reported as a major inciting organism for anthracnose in many tropical and subtropical crops [33, 34].
Conclusion
Numerous investigations on the pathogenicity and host range of C. gloeosporioides have been reported, but the current study reports the pathogenic occurrence of C. gloeosporioides on B. edulis. Our report has provided information on a new pathosysmtems of C. gloeosporioides in PNG, where only known pathosysmtems are capsicum, yams, and coffee.
Acknowledgements
This work was financially supported by the UNITECH Research Committee, Papua New Guinea University of Technology. The authors thanked Professor Tom Okpul and Dr Stanis Malangen for proof-reading the manuscript and providing the comments to improve the paper.
References
-
Bourke RM (1996) Edible Indigenous Nuts in Papua New Guinea. In: Stevens ML, et al. (Eds.), South Pacific Indigenous Nuts, Lagon Resort, Port Vila, Vanuatu.
-
Wah LC (1996) Marketing Indigenous Nuts in Vanuatu a Private Enterprise Perspective. In: Stevens ML, et al. (Eds.), South Pacific Indigenous Nuts. Proceedings of a workshop held from 31 October to 4 November 1994 at Le Lagon Resort, Port Vila, Vanuatu. ACIAR Proceedings: no: 69, pp: 162-172.
-
Bourke RM (2010b) Indigenous Fruit in Papua New Guinea. In: Quartermain AR, et al. (Eds.), Fruits and Nuts: Research and Development Issues in Papua New Guinea, National Agricultural Research Institute, Papua New Guinea.
-
Kaur M, Singh G, Mohan C (2013) Barringtonia acutangula: A Traditional Medicinal Plant. Int J Pharm Sci Rev Res 23(1): 168-171.
-
Mishra S, Sahoo S (2013) Medicinal Properties and Biological Activities of Barringtonia acutangula Linn: A Review. World Journal of Pharmacy and Pharmaceutical Sciences 2(4): 1781-1788.
-
Pauku RL (2006) Barringtonia procera (cutnut), version 2.1. In: Elevitch CR, et al. (Eds.), Species profiles for pacific island agroforestry. Permanent Agriculture Resources (PAR).
-
Ferra P (1996) Country Priorities for Research and Development. In: Stevens ML, et al. (Eds.), South Pacific Indigenous Nuts. Lagon Resort, Port Vila, Vanuatu.
-
Braun U, Mouchacca J (2000) Cercosporoid hyphomycetes on Barringtonia spp. Sydowia 52: 73-77.
-
Braun U, Mouchacca J, McKenzie EHC (1999) Cercosporoid hyphomycetes from New Caledonia and some other South Pacific islands. New Zealand Journal of Botany 37(2): 297-327.
-
Hyde KD, Cai L, McKenzie EHC, Yang YL, Zhang JZ, et al. (2009a) Colletotrichum: a catalogue of confusion. Fungal Diversity 39: 1-17.
-
Ariyawansa HA, Maharachchikumbura SSN, Karunarathne SC, Chukeatirote E, Bahkali AH, et al. (2013) Deniquelata barringtoniae gen. et sp. nov, associated with leaf spots of Barringtonia asiatica. Phytotaxa 105(1): 11-20.
-
Lakshmi PJ, Selvi KV (2013) Anticancer potentials of secondary metabolites from endophytes of Barringtonia acutangula and its molecular characterization. Int J Curr Microbiol App Sci 2(2): 44-45.
-
Buyoyu P, Maino M, Okpul T (2017) First report of Colletotrichum gloeosporioides species complex causing anthracnose on leaves of cutnut, Barringtonia edulis, in Papua New Guinea. New Disease Reports 35(1): 7.
-
Phoulivong S, McKenzie EHC, Hyde KD (2012) Cross infection of Colletotrichum species: a case study with tropical fruits. Current Research in Environmental & Applied Mycology 2(2): 99-111.
-
Choi YW, Hyde KD, Ho WH (1999) Single spore isolation of fungi. Fungal Diversity 3: 29-38.
-
Wijedasa MH, Liyanapathirana LVC (2012) Evaluation of an alternative slide culture technique for the morphological identification of fungal species. Sri Lankan Journal of Infectious Diseases 2: 47-52.
-
Martinez EP, Hío JC, Osorio JA, Torres MF (2009) Identification of Colletotrichum species causing anthracnose on Tahiti lime, tree tomato and mango. Agron colomb 27(2): 211-218.
-
Grahovac M, Indic D, Vukovic S, Hrustc J, Gvozdenac S, et al. (2012) Morphological and ecological features as differentiation criteria for Colletotrichum species. Agriculture 99(2): 189-196.
-
Zivkovic S, Stojanovic S, Ivanovic Z, Trkulja N, Dolovac N, et al. (2010) Morphological and Molecular Identification of Colletotrichum acutatum from Tomato Fruit. Journal of Pesticide and Phytomedicine 25(3): 231-239.
-
Ayodele MA, Hughes Jd’A, Asiedu R (2004) Yam Anthracnose Disease: Field symptoms and laboratory diagnostics. International Institute of Tropical Agriculture.
-
Dias MD, Dias-Neto JJ, Santos MDM, Formento AN, Bizerra LVAS, et al. (2019) Current status of soyabean anthracnose associated with Colletotrichum truncatum in Brazil and Argentina. Plants 8(11): 495.
-
Photita W, Taylor PWJ, Ford R, Hyde KD, Lumyong S, et al. (2005) Morphological and molecular characterisation of Colletotrichum species form herbaceous plants in Thailand. Fungal Diversity 18: 117-133.
-
Prihastuti H, Cai L, Chen H, McKenzie EHC, Hyde KD, et al. (2009) Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Diversity 39: 89-109.
-
Hassan O, Kim JS, Romain BBND, Chang T (2022) An account of Colletotrichum species associated with anthracnose of Atractylodes ovata in South Korea based on morphology and molecular data. PLoS ONE 17(1): 0263084.
-
Zakaria M, Bailey R (2000) Morphology and cultural variation among Colletotrichum isolates obtained from tropical forest nurseries. Journal of Tropical Forest Science 12(1): 1-20.
-
Denoyes B, Baudry A (1995) Species identification and pathogenicity study of French Colletotrichum strains isolated from strawberry using morphological and cultural characteristics. Phytopathology 85(1): 53-57.
-
Abang MM, Winter S, Mignouna HD, Green KR, Asiedu R (2003) Molecular taxonomic, epidemiological and population genetic approaches to understanding yam anthracnose disease. African Journal of Biotechnology 2(12): 486-496.
-
Weir BS, Johnston PR, Damm U (2012) The Colletotrichum gloeosporioides species complex. Studies in Mycology 73: 115-180.
-
Sutton BC (1992) The genus Glomerella and its anamorph Colletotrichum. Colletotrichum: Biology, pathology and control. CAB International Wallingford. UK, pp: 1-26.
-
Damm U, Cannon PF, Woudenberg JHC, Crous PW (2012) The Colletotrichum acutatum species complex. Studies in Mycology 73: 37-113.
-
Hyde KD, Cai L, Cannon PF, Crouch JA, Crous PW, et al. (2009b) Colletotrichum -names in current use. Fungal Diversity 39: 147-182.
-
Sanders GM, Korsten L (2003) Comparison of cross inoculation potential of South African avocado and mango isolates of Colletotrichum gloeosporioides. Microbiology Research 158(2): 143-150.
-
Talhinhas P, Baroncelli R (2021) Colletotrichum species and Complexes: geographic distribution, host range and conservation status. Fungal Diversity 110: 109-198.
-
Jebb M (1992) Edible Barringtonias. Kew Magazine 9(4): 164-180.
- Diversity of Candida sp and Antifungal Susceptibility Patterns in Digestive Candidiasis among People Living with HIV in CHU of Libreville, Gabon
- Vulvovaginal candidiasis: Retrospective study (2019- 2021) at the Centre Hospitalier National de Pikine, Suburban Dakar, Senegal
- Identification of Environmental Fungal Species in Clinical Services of University Hospital of Angre, Abidjan (Cote d’Ivoire)
- New Location of some Gasteroid Basidiomycetes in Western Kazakhstan
- Evaluation of Various Extracellular Enzymes of Ectomycorrhizal Mushrooms
- Morphology and Phylogeny of Lactarius Wallichianae sp. nov and Xerula magnispora sp. nov. from India