Involvement of Secondary Metabolites and Extracellular Lytic Enzymes Produced by Plant Growth Promoting Rhizobacteria in Inhibiting the Soilborne Pathogens in Faba Bean Plants
Plant growth promoting rhizobacteria (PGPR) viz. Pseudomonas fluorrescens, Bacillus megaterium, B. subtilis, and B. cereus and their metabolic products may play a pivotal role in controlling root rot and wilt diseases in faba bean plants caused by Rhizctonia solani, Fusarium solani, F. oxysporium, Macrophomia phaseolina and promote plant growth under greenhouse and field conditions. Cell cultures, extracellular metabolites, volatile metabolites of all tested PGPR strains were suppressed the linear growth of all tested pathogenic fungi in vitro. P. fluorescens followed by B. megaterium were more active than B. subtilis and B. cereus in reduction of the tested fungi radial growth. All PGPR strains were able to produce IAA, HCN, siderophore, Ammonia in media growth. P. fluorescens produced the highest levels of cyanide hydrogen and ammonia followed by B. subtilis, while the higher level of IAA was produced by B. subtilis followed by Ps. fluorescens. Also, B. megaterium was the most PGPR strain produced siderophore followed by Ps. fluorescens. All the tested PGPR strains successfully solubilized inorganic phosphate on Pikovskya’s agar medium. Also, all plant growth promoting rhizobacteria strains (PGPR) were able to produce mycolytic enzymes viz. cellulase, chitinase β-1,3-glucanase, amylase and protease except B. cereus and B. megaterium not able to produce protease and amylase. B. megaterium recorded the highest activities of chitinase, β-1,3-glucanase, while, B. cereus produced the lowest levels of all tested enzymes.
Introduction
Faba bean (Vicia faba L.), family Fabaceae considered the most important nutritive popular food crop in the world and Egypt. It plays a major role in the Egyptian diet as a source of protein. Faba bean crop rich in protein (protein content ranges from 26 to 41%) and the supply of essential amino acids [1]. Several investigators recorded that diseases found on faba bean considered the most destructive and cause considerable losses in yield (estimated at over 50%). Several roots rot and wilt pathogens such as R. solani, F. oxysporium, F. solani and M. phaseolina are reported to attack faba bean roots and stem base causing serious losses in seed germination and plant stand as well Abdel-Monaim MF, et al. [2].
Plant growth promoting rhizobacteria (PGPR) such as Bacillus strains and P. flruorescens are the major root colonizers [3], and can elicit plant defense resistance. Different mechanisms have been reported for their performance such as production of antibiotics, siderophore cyanide hydrogen, competition for nutrition and space; induce resistance, inactivation of pathogen’s enzymes and enhancement of root and plant development [4]. PGPR can prevent the proliferation of fungal and other pathogens by producing siderophores that bind most of the FeIII in the vicinity of the plant root, preventing pathogens from growing close to the plant roots. This is achieved by the PGPR out- competing the pathogens for available iron. Plants are not affected by the localized depletion of soil iron as most plants cantolerate much lower iron concentrations (~1000 fold less) than microorganisms. Similarly, cyanide is a secondary metabolite produced by Gram-negative bacteria.
HCN and CO2 are formed from glycine and catalysed by HCN synthesis. HCN production by strains of PGPR suppresses disease, whereas mutant strains unable to synthesise HCN lose their ability to protect plants from disease. Also, many antifungal metabolites have been produced and shown to be effective in vitro. These antifungal metabolites are also suspected to have antifungal activity in vivo. These metabolites include ammonia, butyrolactones, 2-4-diacetylphloroglucinol, kanosamine, Oligomycin A, Oomycin A, phenazine-1-carboxylic acid (PCA), pyoluterin (Plt), pyrrolnitrin (pln), viscosinamide, xanthobaccin and zwittermycin A. Whipps JM, et al. [5]. In addition, certain fungi have been shown to be sensitive to particular combinations of metabolites.
The microorganisms isolated from the rhizosphere of various crop have the ability to produce indole acetic acid as secondary metabolites due to rich supply of substrates. Indole acetic acid (IAA) helps in the production of longer roots with increased number of root hairs and lateral roots which are involved in nutrient uptake [6]. IAA stimulates cell elongation by modifying certain conditions such as the increase in osmotic contents of the cell, increase in permeability of water into cell, decrease in wall pressure, an increase in cell wall synthesis and inducing specific RNA and protein synthesis. It promotes embial activity, inhibit it promotes embial activity, inhibit or delay abscission of leaves, induce flowering and fruiting [7].
Also, Chitinase, glucanase and other hydrolytic enzymes have many roles in a wide range of different biological systems. These enzymes are usually extracellular, of low molecular weight and high stability. In addition, they may be produced in multiple forms or isozymes that differ in charge, size, regulation, stability and ability to degrade cell walls [8]. The objectives of the present study were to test the possible role of in vitro production of mycolytic enzymes viz. chitinase, β-1, 3-glucanase, protease siderophores, HCN and IAA by PGPR strains viz. B. subtilis and B. megaterium in suppression of F. oxysporum f. sp. ciceris. In addition, to study the control wilt disease under artificial inoculation in greenhouse and under natural infection under field conditions as well as its effect on growth and yield parameters under field conditions.
Material and Methods
Research Site
All experiments were carried out at the Agricultural Research Station in the New Valley, New Valley Governorate and Egypt.
Source of Fungal Pathogen
The highly pathogenic isolate each of R. solani, F. solani, F. oxysporum and M. phaseolina isolated from diseased chickpea plants collected from New Valley Governorate was used in this study Khalil SM, et al. [9].
Preparation of Fungal Inoculum
The inoculum of pathogenic fungi was prepared from one week old culture grown on 50 mL potato dextrose broth (PDB) medium in conical flask (250 mL) and incubated at 25 ± 1°C. The content of flask was homogenized in a blender for one min. Plastic pots were filled with sterilized soil and mixing with fungal inocula at rate of 100 mL homogenized culture per pot, seven days before planting.
Source of the PGPR
Plant growth promoting rhizobacteria (PGPR) namely, B. megaterium ((isolate BMM5), B. subtilis (isolate BSM1) and B. cereus (BCM8) and Ps. fluorescens (PFM1) were obtained from the Laboratory. of Plant Pathology Department, New Valley Agric. Res. Station.
In Vitro Screening of Inhibitory Effect of PGPR
The tested isolates of antagonistic PGPR were streaked at one side on PDA medium in plates and incubated for 24 hours at 25°C±1, then one disc (7 mm in diameter) of tested pathogenic fungi viz. R. solani, F. solani, M. phaseolina and F. oxysporum was placed on the opposite side [10]. The inoculated plates with pathogenic fungus only were used as control. After 3-7 days incubation, linear growth of pathogenic fungi in all treatments was recorded. The decrease of percentage that occurred in linear growth of the pathogenic fungi was determined at the end of the experiment using formula as follows: Reduction in linear growth = [(R1- R2)/R1] ×100 R1= the radius of normal growth in control plates; R2= the radius of inhibited growth
Enzymatic Activities and Secondary Metabolites of the PGPR Strains
Production of Indole Acetic Acid (IAA)
The production of IAA was determined by colorimetric measurement at 530 nm using Salkowski’s reagent. Bacteria were grown under shaking (120 rpm) for 2 days at 30ºC in Luria Bertani broth medium (Tryptone 10.0 g, Yeast Extract 5.0 g Sodium Chloride 10.0 g) supplemented with tryptophan (1 mg mL−1) as IAA precursor. After incubation, the cells were centrifuged (3,000 rpm for 10 min at 4°C) and 1 mL of supernatant was combined with 2 mL of Salkowski’s reagent (150 mL of 95-98% H2SO4, 7.5 mL of 0.5 M FeCl3•6H2O, and 250 mL distilled water) and incubated for 30 min at room temperature. The quantification of IAA was carried out using a standard curve of pure IAA (Sigma−Aldrich, Co.).
Production of Hydrogen Cyanide (HCN)
Bacterial strains were grown at 25±2°C on a rotary shaker in Tryptic Soy Broth (TSB). Filter paper (Whatman No. 1) was cut into uniform strips of 10 cm long and 0.5 cm wide saturated with alkaline picrate solution and placed inside the conical flasks in a hanging position. After incubation at 25±2°C for 48 h, the sodium picrate present in the filter paper was reduced to reddish compound in proportion to the amount of hydrocyanic acid evolved. The developed color was eluted in 10 mL of distilled water and the absorbance was measured at 625 nm.
Production of Siderophore
Bacterial strains were grown in KB broth for 3 days at 25±2°C and centrifuged at 3000 rpm for 10 min and the supernatants were collected. The pH of the supernatant was adjusted to 2.0 with diluted HCl and equal quantity of ethyl acetate was added in a separating funnel, mixed well and ethyl acetate fraction was collected. Five milliliters of ethyl acetate fraction were mixed with 5mL of Hathway’s reagent (1.0 mL of 0.1M FeCl3 in 0.1 N HCl to 100 mL distilled water1.0 mL of potassium ferricyanide). The absorbance for dihydroxy phenols was read at 700 nm. A standard curve was prepared using dihydroxy benzoic acid. The quantity of siderophore synthesized was expressed as μmol benzoic acid/mL of culture filtrate.
Ammonia Production
Freshly grown culture was inoculated into peptone water and incubated for 7, 10, and 13 days at 27±2°C. Broth was collected, centrifuged and the amount of ammonia in the supernatant was estimated by means of Nesslerization reaction 1 mL Nessler’s reagent was added to 1 mL of supernatant and volume of this mixture was made up to 10 mL by addition of ammonia-free distilled water. Development of brown to yellow color was a positive test for ammonia production and optical density was measured by spectrophotometer at 450 nm [11]. The concentration of ammonia was estimated based on a standard curve of ammonium sulfate ranging from 0.1 to 1 μmol mL-1.
Phosphate Solubilization
To detect the ability of organisms to solubilize phosphate, isolates were spot inoculated using sterile tooth picks onto Pikovskaya’s Agar medium containing (per liter) tricalcium phosphate 2.5 g, glucose 13 g, (NH4) SO4 0.5 g, NaCl 0.2 g, MgSO4.7H2O 0.1 g, KCl 0.2 g, yeast extract 0.5 g, MnSO4 trace, FeSO4.7H2O trace, agar 15 g, pH adjusted to 7.2. Plates were incubated for three days at 28 ± 2°C [12]. The strains showing halo zone of tricalcium phosphate solubilization were considered positive. Phosphate solubilization index (SI) was calculated by measuring the diameter of halo zone and bacterial colony using the formula Edi-Premoto M, et al. [13].
SI (Solubility Index) = colony diameter + halo zone diameter/colony diameter
Assay of Chitinase
PGPR strains were cultured at 25±2 °C for 96 hr. on a rotary shaker in 250 mL conical flasks containing 50 mL of chitin–peptone medium (glucose 0.5%, peptone 0.2%, colloidal chitin 0.2%, K2HPO4 0.1%, MgSO4 .7H2O 0.05% and NaCl 0.05%, pH 6.8). The cultures were centrifuged at 12,000 g for 20 min at 4°C and the supernatant was used as enzyme source. Colloidal chitin was prepared from crab shell chitin according to Berger LR, et al. [14]. The reaction mixture contained 0.25 mL of enzyme solution, 0.3 mL of 1M sodium acetate buffer (pH 5.3) and 0.5 mL of colloidal chitin (0.1%). The reaction mixture was incubated at 50°C for 4 h in a water bath. Chitinase activity was determined by measuring the release of reducing sugars by the method of Nielsen MN, et al. [15]. One unit of chitinase was determined as 1 nmol of N-acetyl-β-D-glucosamine (GlcNAc) released per minute per mg of protein.
Assay of β-1, 3-glucanase
Bacterial strains were grown at 25±2°C for 96 hr. on a rotary shaker in 250 mL conical flasks containing 50 mL of peptone medium containing laminarin (0.2%) (Laminaria digitata; Sigma). The cultures were then centrifuged at 12,000 g for 20 min at 4°C and the supernatant was used as enzyme source. The reaction mixture contained 0.25 mL of enzyme solution, 0.3 mL of 0.1M phosphate buffer (pH 5.5) and 0.5 mL of laminarin (0.2%). The reaction mixture was incubated at 40°C for 2 h in water bath. β- 1,3-glucanase activity was determined as 1 nmol of glucose released/min./ mg of protein.
Protease Production
Bacterial strains were streaked on casein agar and/ or incubated at 28 ± 2°C for four days. At the end of the incubation, the plates were observed for halo zone around the colonies, which indicates the presence of protease. The halo zone diameters were recorded.
Cellulase Production
Carboxy methyl cellulose (CMC) agar plates were prepared by screening for cellulose enzyme production according to the method by Kasana RC, et al. [16]. A sterile paper disc was dipped into microbial culture and transferred onto the CMC agar plates. The plates were incubated overnight at 33°C. After incubation, the plates were flooded with Congo red solution for 15min; followed by de-staining with the salt solution for 15min. unstained areas indicate where the CMC has been degraded due to production of cellulose by the bacterial strain. The halo zone diameters were recorded.
Amylase Production
Amylase production was determined by the method described by Alariya SS, et al. [17]. Fresh bacterial cultures were inoculated on Starch Agar plate and incubated at room temperature for 24-48 hr. Plate was flooded by Iodine solution after incubation. The hydrolysis of starch is observed as colorless zone around growth with violet background and is reported as trace to 4.
Determination of Extracellular Compounds
A 0.2 μm cellophane membrane was placed on PDA plates and 200μL of antagonistic bacterial suspension (1 × 107 cfu/mL) were inoculated in the center of plates. The plates were incubated at 25±2°C for 48 h, then the membrane with the grown bacterial strains was removed and the plate was inoculated in the middle with a five mm disk of a pure culture of R. solani, F. solani, M. phaseolina and F. oxysporum, individually. Plates were incubated at 25±2°C for 7 days and the radial growth of the pathogen was measured. Sterile double-distilled water replaced the bacterial suspension in control plates. There were four replicates for each treatment [18, 19].
Production of Volatile Antibiotics
Firstly, 200μL of bacterial suspension (1 × 10 7 cfu/mL) from each strain were spread on the surface of a Petri plate containing nutrient agar medium and incubated at 25±2°C for two days. In another Petri plate containing PDA medium, a 5 mm disk of a 7-days-old culture of tested pathogenic fungi viz. R. solani, F. solani, M. phaseolina and F. oxysporum was placed at the center. Then both half plates were placed face to face preventing any physical contact between the pathogen and the bacterial suspension [20]. The plates were sealed with parafilm. In the control plates, pathogenic fungi were placed at the center. Plates were incubated at 25±2°C for 48 hr. and the percentage of inhibition was calculated for each isolate in four replicates for each treatment.
Statistical Analysis
Analyses of variance were carried out using MSTATC, 1991 program (Ver. 2.10). Least significant difference was employed to test for significant difference between treatments at p ≤ 0.05 [21].
Results and Discussion
In vitro Screening Inhibitory Effect of PGPR
Plant growth promoting rhizobacteria viz., B. megaterium, B. subtilis, B. cereus and Ps. fluorescens strains were evaluated for antagonistic effect against R. solani, F. solani, M. phaseolina and F. oxysporum on Petri dishes containing PDA medium. Data in Figure 1 show that all PGPR strains succeeded in reducing the radial growth of all tested pathogenic fungi. Ps. fluorescens followed by B. megaterium were more active than B. subtilis and B. cereus for suppressing redial growth of tested fungi. On the other hand, F. solani followed by F. oxysporum was more affecting with PGPR, while R. solani and M. phaseolina was the lowest one. Such results confirm previous reports indicating that PGPR has the potential to inhibit growth of soil borne pathogens [22, 23, 24]. Glick BR, et al. [25] reported that the most effective mechanism that a PGPR can employ to prevent proliferation of phytopathogens in the synthesis of antibiotics.
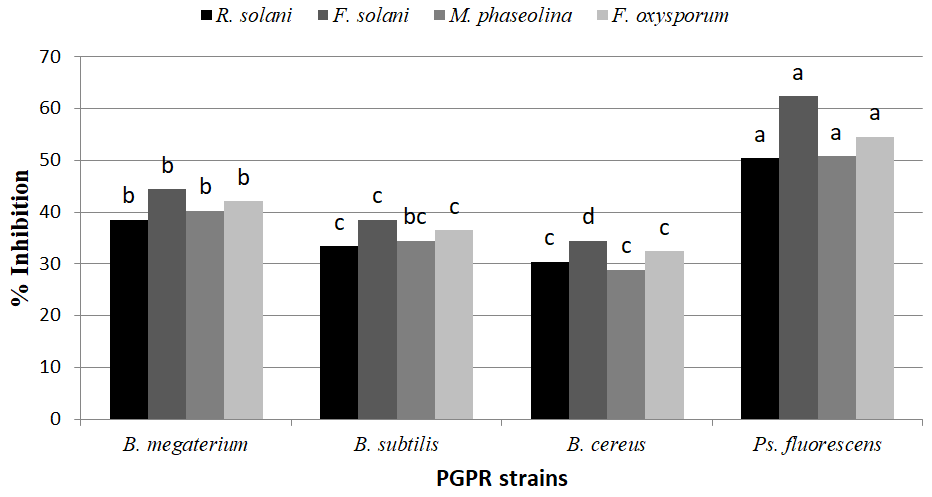
Enzymatic Activities and Secondary Metabolites of the PGPR Strains
Results of testing 4 PGPR strains (B. megaterium, B. subtilis, B. cereus, Ps. fluorescens) in vitro against tested pathogenic fungi showed that different strains exhibited different combinations of antimicrobial metabolites such as HCN, Siderophore, IAA, Ammonia production and Phosphate solubilisation. Pseudomonas fluorescens produced the highest levels of cyanide hydrogen and ammonia followed by B. subtilis. While, the higher level of IAA was produced B. subtilis followed by Ps. fluorescens. On the other hand, B. megaterium is the most PGPR strain produced of siderophore followed by Ps. fluorescens. On the other hand, all the tested PGPR strains successfully solubilized inorganic phosphate on Pikovskya’s agar medium (Table 1), indicated by forming a clear halo around their PGPR colony. The maximum phosphate solubilization after 72 hr. for in plates inoculated with B. megaterium (21.3mm) followed by Ps. fluorescens (18.7mm), while plates inoculated with B. subtilis and B.
cereus where recorded phosphate solubilization 13.8 and 14.8 mm, respectively.
On the other hand, all plant growth promoting rhizobacteria strains (PGPR) were able to produce mycolytic enzymes viz. cellulase, chitinase β-1,3-glucanase, amylase and protease in growth media except B. cereus and B. megaterium not able to produce protease and amylase in media growth, respectively (Table 1). B. megaterium recorded the highest activities of chitinase, β-1,3-glucanase in media growth compared with the other PGPR strains, whereas produced 23.12 and 80.12 nmol/min/mg protein, respectively. While, Ps. fluoresces produced the highest protease and amylase in media growth (20.08 and 14.75 halo zone diameter). B. subtilis produced the highest cellulase level in media growth (16.35 halo zone diameter). On the other hand, B. cereus produced the lowest levels of all tested enzymes (chitinase, β-1,3-glucanase, protease, cellulase and amylase) in media growth).
| Antifungal Metabolites | B. megaterium | B. subtilis | B. cereus | Ps. fluorescens |
|---|---|---|---|---|
| Production of IAA (ìg/mL) | 0.805 | 1.124 | 0.736 | 1.086 |
| Production of HCN (OD at 625 nm) | 0.098 | 0.075 | 0.157 | 0.198 |
| Production of siderophore (ìmol benzoic acid/mL) | 6.25 | 4.52 | 3.56 | 5.42 |
| Ammonia production (µg/mL) | 25.6 | 32.5 | 24.1 | 36.5 |
| Phosphate solubilization index (mm) | 21.3 | 13.8 | 14.8 | 18.7 |
| Chitinase activity (nmol/min/mg protein) | 23.12 | 17.45 | 15.43 | 19.69 |
| β-1,3-glucanase activity (nmol/min/mg protein) | 80.12 | 72.35 | 62.14 | 77.43 |
| Protease production (halo zone diameter with mm) | 14.35 | 8.36 | 0 | 20.08 |
| Cellulase production (halo zone diameter with mm) | 12.52 | 16.35 | 10.41 | 14.85 |
| Amylase production (halo zone diameter with mm) | 0 | 12.42 | 10.95 | 14.75 |
Table 1: In vitro production of various metabolites, enzymes by PGPR strains.
The results agree with Sarhan EAD, et al. [24]; Kumar S, et al. [26]; Choudhary SR, et al. [27]. Arora NK, et al. [28] found that the lytic enzymes produced by bio-control agents interfere with the growth and activities of pathogen by hydrolyzing the polymeric compounds, including chitin, proteins, celluloses, hemicelluloses, etc., and consequently killing or suppressing the growth of pathogens. Hydrolytic enzymes showed their ability to control plant pathogens. A hydrolytic enzyme like chitinase, glucanase, protease and cellulase can degrade the fungal cell-wall and causes the cell lysis of fungal pathogens. Also, Abdel-Monaim MF, et al. [20] found that PGPR strains viz. B. subtilis and B. megaterium were able to produce indole acetic acid (IAA), siderophore, hydrogen cyanide (HCN), extracellular compound and volatile antibiotics in vitro. In addition, both PGPR strains produced mycolytic enzymes viz. chitinase, β-1, 3-glucanase and protease in growth media. B. megaterium produced greater amounts of secondary metabolites than B. subtilis.
Effect of Extracellular Metabolites
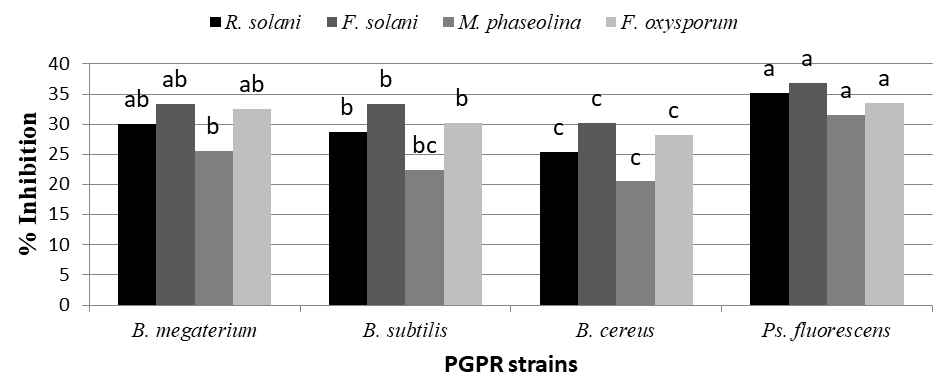
Data present in Figure 2 show that all plant growth promoting rhizobacteria (B. megaterium, B. subtilis, B. cereus and Ps. fluorescens produced extracellular metabolites. Ps. fluorescens and B. megaterium was significantly the highest produce extracellular metabolites against all tested pathogenic fungi. While B. cereus and B. subtilis recorded the lowest produce extracellular metabolites in case of all pathogenic fungi. On the other hand, F. solani was the most affecting with extracellular metabolites produced with PGPR, while M. phaseolina was the least affecting ones in this respect.
These results revealed that certain extracellular metabolites generated by PGPR affected mycelial growth and germ tube elongation. Some Bacillus species are known to produce numerous antimicrobial compounds which have been well characterized genetically and biochemically in vitro Arguelles-Arias A, et al. [29]. Among these antagonistic compounds, the predominant lipopeptides of the surfactin, iturin and fengycin families have been well reported for their potential against a wide range of plant pathogens Pathak KV, et al. [30].
Effect of Volatile Antibiotics
ANOVA results showed that the colony diameter of tested pathogenic fungi (R. solani, F. solani, M. phaseolina and F. oxysporum varied significantly (P ≤ 0.05) depending on antagonistic treatments tested. In fact, data given in Figure 3. Revealed that all tested pathogenic growth decrease, due to the inhibitory effects of volatile metabolites from the rhizobacteria strains tested were varied by differently PGPR strains and pathogenic fungi. Ps. fluorescens was recorded the highest inhibitory effect against all tested pathogenic fungi followed by B. megaterium, while B. subtilis and B. cereus gave the lowest ones against all pathogenic fungi in this respect. Volatile inhibitors that have been identified include ethylene, ammonia, allyl alcohol, acrylic acid, trimethylamine, benzaldehyde, and N,N-dimethyloctylamine [31, 32]. The spectrum of volatile compounds produced appears to depend not only on environmental conditions, such as nutrients, temperature, pH and moisture, but also on the organism itself. Ryu CM, et al. [33] demonstrated significant growth promotion of Arabidopsis by B. subtilis strain GB03 and B. amyloliquefaciens strain IN937a. Among VOCs released from strain GB03, 2,3-butanediol was found as a major component that promoted plant growth and elicited ISR against Erwinia carotovora subsp. carotovora. Several mutant lines of Arabidopsis, including brassinosteroid- and gibberellic acid-insensitive mutants, auxin-transport- deficient mutants, and cytokinin receptor-deficient mutants, were used to elucidate the signaling pathways that promote growth.
![Figure 3: Revealed that all tested pathogenic growth decrease, due to the inhibitory effects of volatile metabolites from the rhizobacteria strains tested were varied by differently PGPR strains and pathogenic fungi. Ps. fluorescens was recorded the highest inhibitory effect against all tested pathogenic fungi followed by B. megaterium, while B. subtilis and B. cereus gave the lowest ones against all pathogenic fungi in this respect. Volatile inhibitors that have been identified include ethylene, ammonia, allyl alcohol, acrylic acid, trimethylamine, benzaldehyde, and N,N-dimethyloctylamine [31,32]. The spectrum of volatile compounds produced appears to depend not only on environmental conditions, such as nutrients, temperature, pH and moisture, but also on the organism itself. Ryu CM, et al. [33] demonstrated significant growth promotion of Arabidopsis by B. subtilis strain GB03 and B. amyloliquefaciens strain IN937a. Among VOCs released from strain GB03, 2,3-butanediol was found as a major component that promoted plant growth and elicited ISR against Erwinia carotovora subsp. carotovora. Several mutant lines of Arabidopsis, including brassinosteroid- and gibberellic acid-insensitive mutants, auxin-transport- deficient mutants, and cytokinin receptor-deficient mutants, were used to elucidate the signaling pathways that promote growth.](/fulltextimages/8623/fig_3.png)
Conclusion
The application of PGPR and its secondary metabolites for controlling Fusarium wilt disease in fababean. Their low cost, low toxicity to man and environmental pollution make them ideal for seed soaking for disease control under field conditions and also for increased seed yield and seed content of protein.
Acknowledgments
The researchers extend their sincere thanks and appreciation to Prof. Dr. Montaser Fawzy Abdel-Moneim, Head of the Plant Diseases Department at the Regional Station for Agricultural Research in the New Valley, for his assistance in carrying out laboratory experiments at the station.
Funding
The research was funded by the Agricultural Research Center of the Egyptian Ministry of Agriculture within the ministry’s plan to raise the efficiency of scientific research in the Arab Republic of Egypt
Authors’ Contributions
Manal Sayed Mohammed implemented the research plan, applied it on the ground, and carried out experiments in the laboratory; Mohamed Hassan Abdel-Rahem Hassan developed a research plan and provided the necessary capabilities for the application and Amer Fayz Mahmoud & Kadry Mostafa Mohamed Morsy Contributed to data analysis, statistical analysis and research writing.
Competing Interest
The research is an introduction to its publication to help the researcher to be promoted to a higher degree
References
-
Fernandez M, Lopez-Jurado M, Aranda P, Urbano G (1996) Nutritional assessment of raw and processed faba bean (Vicia faba L.) cultivar major in growing rats. J Agric Food Chem 44(9): 2766-2772.
-
Abdel-Monaim MF (2013) Improvement of biocontrol of damping-off and root rot/wilt of faba bean by salicylic acid and hydrogen peroxide. Mycobiology 41(1): 47-55.
-
Souad Z, Belabid L, Bellahcene M (2013) Biocontrol of chickpea Fusarium wilt by Bacillus spp. Rhizobacteria. J Plant Prot Res 53(2): 177-183.
-
Karimi K, Amini1 J, Harighi B, Bahramnejad B (2012) Evaluation of biocontrol potential of Pseudomonas and Bacillus spp. against Fusarium wilt of chickpea. Aust J of Crop Sci 6(4): 695-703.
-
Whipps JM (2001) Microbial interactions and biocontrol in the rhizosphere. J Exp Bot 52(1): 487-511.
-
Datta C, Basu PS (2000) lndole acetic acid production by a Rhizobium species from root nodules of a leguminous shrub Cajanus cojan. Microbiol Res 155(2): 123-127.
-
Narasimhan A, Bist D, Suresh S, Shivakumar S (2013) Optimization of mycolytic enzymes Chitinase, β-1,3- Glucanase and Cellulase production by Bacillus subtilis, a potential biocontrol agent using one-factor approach. J Sci Ind Res 72(3): 172-178.
-
Ashwini N, Srividya S (2014) Potentiality of Bacillus subtilis as biocontrol gent for management of anthracnose disease of chilli caused by Colletotrichum gloeosporioides OGC1.3 Biotech 4(2): 127-136.
-
Khalil SM (2019) Utilizing Some Disease Resistance Inducing Agents and Magnetized Water in the Management of Root Rot and Wilt of Faba Bean. Ph. D. Thesis. Faculty Agriculture, Assuit University.
-
Kaur R, Kaur J, Singh RS, Alabouvette C (2007) Biological control of Fusarium oxysporumf sp ciceris by non- pathogenic Fusarium and fluorescent Pseudomonas. Int J Bot 3(1): 114-117.
-
Demutskaya LN, Kalinichenko IE (2010) Photometric determination of ammonium nitrogen with the nessler reagent in drinking water after its chlorination. J Water Chem Tech 32(2): 90-94.
-
Nautiyal CS (1999) An efficient microbiological growth medium for screening phosphate solubilizing bacteria. FEMS Microbiol Lett 170(1): 265-270.
-
Edi-Premoto M, Moawad AM, Vlek PLG (1996) Effect of phosphate solubilizing Pseudomonas putida on the growth of maize and its survival in the rhizosphere. Indonesian J Crop Sci 11: 13-23.
-
Berger LR, Reynolds DM (1958) The chitinase system of a strain of Streptomyces griseus. Biochim Biophys Acta 29(3): 522-534.
-
Nielsen MN, Sorensen J (1999) Chitinolytic activity of Pseudomonas fluorescens isolates from barley and sugar beet rhizosphere. FEMS Microbiol Ecol 30(3): 217-227.
-
Kasana RC, Salwan R, Dhar H, Dutt S, Gulati A (2008) A rapid and easy method for the detection of microbial cellulases on agar plates using gram’s iodine. Curr Microbiol 57(5): 503-507.
-
Alariya SS, Sethi S, Gupta S, Gupta BL (2013) Amylase activity of a starch degrading bacteria isolated from soil Arch Appl Sci Res 5(1): 15-24.
-
Naureen Z, Price AH, Hafeez FY, Roberts MR (2009) Identification of rice blast disease suppressing bacterial strains from the rhizosphere of rice grown in Pakistan. Crop Prot 28(12): 1052-1060.
-
Kraus J, Loper JE (1990) Biocontrol damping-off of cucumber by Pseudomonas fluorescens pf-5: mechanistic studies. In: Keel CB, et al. (Eds.). Plant growth promoting rhizobacteria. The Second International Workshop on Plant Growth Promoting Rhizobacteria, Interlacen, Switzerland, pp: 172-175.
-
Abdel-Monaim MF (2016) Efficacy of secondary metabolites and extracellular lytic enzymes of plant growth promoting rhizobacteria (PGPR) in controlling Fusarium wilt of chickpea. Egypt J Agric Res 94(3): 573-
-
Gomez KA, Gomez AA (1984) Statistical Procedures for Agricultural Research. A Wiley Interscience Publication, New York, pp: 1-690.
-
Abdel-Monaim MF (2010) Integrated management of damping-off, root and/or stem rot diseases of chickpea with sowing date, host resistance and bioagents. Egypt J Phytopathol 38(1): 45-61.
-
Susilowati A, Wahyudi AT, Lestari Y, Suwanto A, Wiyono S (2011) Potential Pseudomonas isolated from soybean rhizosphere as biocontrol against soil-borne phytopathogenic fungi. Hayati J Biosciences 18(2): 51- 56.
-
Sarhan EAD, Shehata HS (2014) Potential plant growth- promoting activity of Pseudomonas spp. and Bacillus spp. as biocontrol agents against damping-off in alfalfa. Plant Pathol J 13(1): 8-17.
-
Glick BR (1995) The enhancement of plant growth by free-living bacteria. Can J Microbiol 41(2): 109-117.
-
Kumar S, Pandey P, Maheshwari DK (2009) Reduction in dose of chemical fertilizers and growth enhancement of sesame (Sesamum indicum L.) with application of rhizospheric competent Pseudomonas aeruginosa LES4. Eur J Soil Biol 45(4): 334-340.
-
Choudhary SR, Sindhu SS (2015) Suppression of Rhizoctonia solani root rot disease of clusterbean (Cyamopsis tetragonoloba) and plant growth promotion by rhizosphere bacteria. Plant Pathology J 14(2): 48-57.
-
Arora NK, Kim MJ, Kang SC, Maheshwari DK (2007) Role of chitinase and beta-1,3-glucanase activities produced by a fluorescent pseudomonad and in vitro inhibition of Phytophthora capsici and Rhizoctonia solani. Can J Microbiol 53(2): 207-212.
-
Arguelles-Arias A, Ongena M, Halimi B, Lara Y, Brans A, et al. (2009) Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb Cell Fact 8: 63.
-
Pathak KV, Keharia H (2013) Characterization of fungal antagonistic bacilli isolated from aerial roots of banyan (Ficus benghalensis) using intact-cell MALDI-TOF mass spectrometry (ICMS). J Appl Microbiol 114(5): 1300- 1310.
-
Chuankun X, Minghe M, Leming Z, Keqin Z (2004) Soil volatile fungistasis and volatile fungistatic compounds. Soil Biol Biochem 36(12): 1997-2004.
-
El-Khoury W, Makkouk K (2010) Integrated plant disease management in developing countries. J Plant Pathol 92(4): S35-S42.
-
Ryu CM, Farag MA, Hu CH, Reddy MS, Kloepper JW (2004) Bacterial volatiles induce systemic resistance inArabidopsis. Plant Physiol 134(3): 1017-1026.
- Diversity of Candida sp and Antifungal Susceptibility Patterns in Digestive Candidiasis among People Living with HIV in CHU of Libreville, Gabon
- Vulvovaginal candidiasis: Retrospective study (2019- 2021) at the Centre Hospitalier National de Pikine, Suburban Dakar, Senegal
- Identification of Environmental Fungal Species in Clinical Services of University Hospital of Angre, Abidjan (Cote d’Ivoire)
- New Location of some Gasteroid Basidiomycetes in Western Kazakhstan
- Evaluation of Various Extracellular Enzymes of Ectomycorrhizal Mushrooms
- Morphology and Phylogeny of Lactarius Wallichianae sp. nov and Xerula magnispora sp. nov. from India