Studies on the Growth and Development of Root-Knot Nematode Meloidogyne incognita of Phaseolus vulgaris in Tissue Culture Media
The study was conducted to study the growth and development of root-knot nematode, Meloidogyne incognita of Phaseolus vulgaris in tissue culture media. The data revealed the developmental common bean cv. Paulista of M.S media as the following: the seed swelling stage after three days, the radical appearance stage after five days. The cotyledons leaves appear after six days, the cotyledon leaves appear after eight days, and the seedling stage after twelve days. The life cycle of M. incognita completed about thirty-seven days in root tissues of Paulista cultivar. Microscopic examinations of the infected bean roots revealed the following: Second-stage juveniles of M. incognita penetrated the roots within 24 hrs after nematode inoculation. Third-stage juveniles were observed in roots (six-eight) days after nematode inoculation. While, fourth stage juveniles were observed in roots 12-14 days after nematode inoculation. The pre-adult females were seen inside roots within twenty days after nematode inoculation. The adult females were seen inside roots within twenty-seven days after nematode inoculation. Egg laying females with deposit egg masses of M. incognita was observed thirty-seven days after nematode inoculation.
Introduction
Common bean (Phaseolus vulgaris L.) is an economically important crop and one of the major grain legumes for human consumption in Latin America, Africa and Asia [1, 2]. Despite its importance, production growth rates are limited by viral, nematode, fungal, and bacterial pathogens, insects, lack of drought tolerances and nutritional deficiencies [3]. Therefore, there is considerable interest in the development of new bean cultivars with useful agronomical traits [3]. Plant biotechnology, together with conventional breeding methods, could facilitate common bean improvement since resistance or tolerance to biotic and abiotic stress could be increased and seed quality, plant architecture, and reproduction modes could be altered [4]. Nevertheless, a reliable and efficient in vitro culture Root-knot nematodes invade an array of important crops and have been found more damaging to vegetables, especially in the tropical and subtropical countries of the world [5]. So far, more than 100 species of Meloidogyne have been described throughout the world and, among these, four species viz. Meloidogyne incognita, M. javanica, M. arenaria and M. hapla, which are commonly found. More than 3000 plant species, almost all cultivated plants, have been recorded as the hosts of root- knot nematodes. The infested plants manifest symptoms of chlorosis, stunting, and unthrifty growth [6].
The life cycle of Meloidogyne includes six developmental stages: the egg stage, four juvenile stages (J1–J4), and the adult stage. The life cycle begins when the only infective and mobile stage, J2, penetrates the roots. Then, the nematode migrates through intercellular spaces to establish a permanent feeding site in the vascular cylinder, in which it will induce the differentiation of five to seven cells, named giant cells, that experience morphological, physiological, and molecular modifications caused by the J2 infection and will supply food to the nematode. The root tissue becomes distorted due to the hyperplasia around the cell, forming the root gall. Once the nematode establishes its feeding site, it will become sedentary and will grow in length and width (J3– J4), until reaching the adult stage. In favorable conditions, J2 develops in pear-shaped females. They lay eggs into a gelatinous matrix (egg mass), generally outside the gall. Embryogenesis will lead to a J1 developing inside the egg, which will molt to J2 that will emerge and migrate into the soil. Only males appear under unfavorable conditions, and they will migrate into the soil with no evidence of feeding on plants [7].
Plant tissue cultures have been utilized to propagate some species of tissue invading nematodes. In most studies on Meloidogyne spp., investigators have employed larvae or eggs from roots growing in infested soil. This technique is a simple, inexpensive method for maintaining an abundant supply of M. incognita in the laboratory. By utilizing glass capillary tubes for inoculations, single egg sacs can be isolated to establish and perpetuate pure nematode cultures. Most of the protocols for in vitro regeneration of common bean are based on direct organogenesis or shoot development of different types of exploits, viz. shoot apical meristems, cotyledonary and primary leaf node explants, petiole etc. [8, 9]. However, these procedures yield very low regeneration efficiency [10]. Another approach has been done through indirect in vitro regeneration. To date, there are only four reports on indirect regeneration of P. vulgaris. However, the frequency of shoot regeneration from callus is extremely low [11, 12] or higher depends on cultivars [13, 14]. Therefore, the objective of the present work was to study the growth and development of root-knot nematode Meloidogyne incognita of Phaseolous vuligaris in tissue culture media.
Materials and Methods
In vitro experiments were conducted at the Department of Agricultural Botany, Faculty of Agric., Suez Canal Univ. The cultivar cv. Paulista was obtained from the Vegetable Crop Res. Depar., Horticul. Res. Inst., Agric. Res. Center, Giza. Culturing of Nematode: The pure culture of M. incognita from single egg mass has been maintained on tomato seedling (Lycopersicon esculantum Mill cv. G.S) at 25±2°C in order to obtain sufficient numbers of second stage juveniles.
Preparation of the Medium
Murashige [15] a medium protocol multiplication were used. The pH of medium was 5.8 by using 1.0NHCLand 1.0 NaOH, and agar was added after adjusting the pH. All used equipment and media were autoclaved for 20 min at 121°C.
Culture Preparation and Inoculation
Seeds of P. vulagris cv. Paulista were surface sterilized by immersing them for 3min in 70% ethanol, followed were surface sterilized with 0.1 sodium hypochlorite (NaOCl) [16] for 10 min and then washed twice with sterile water. Each seed was germinated under aseptic condition in a jar containing 30 ml of the culture medium. Culture was incubated at 28±2° C under 16 hrs photo periods in growth cabinet. Twelve days after incubation, the freshly hatching second stage juveniles of root-knot nematode (j2) were collected and sterilized by 2.5% Clorox for 3 min and then washed with sterile distilled water twice. Finely drawn capillary tubes (1 mm in diameter and 50 mm in length) were used to pick up the concentrated larval suspensions from the depression slide preparations. Using 100 X magnification, the larvae drawn into each capillary tube were counted. Glass capillary tubes containing about 500 j2s /seedling were used to inoculate the host plant (common bean). Capillary tubes containing larvae were inserted into the root. All inoculated plants were grown under greenhouse conditions. Observations were made for at least 2 weeks on all plants. Some plants were examined and studied for intervals: 1, 2, 4, 6, 8, 10, 15,18,20,25,28,30,35, and 37 days following nematode incubation. Microscopic examinations of fresh and fixed sections were made periodically. For the demonstration of nematodes in roots, sections were prepared and stained according to the methods of [17, 18].
Results and Discussion
Developmental Stages of in Vitro Propagation of Common Bean Bean Cv. Paulista by Direct Organogenesis from the Embryonic Axis of Mature Seed The developmental common bean cv. Paulista plant stages of M.S media revealed in Figure 1 as the following: the seed stage in Figure 1A, the seed swelling stage after 3 days Figure 1B, the radical appearance stage after 5 days Figure 1C. The cotyledons leaves appear after 6 days Figure 1D, the cotyledon leaves appear after 8 days Figure 1E, the seedling stage after 12 days Figure 1F. The use of tissue culture for plant regeneration and the introduction of foreign genes that confer pathogens resistance or drought tolerance are good options for bean cultivation. Most of the published protocols are based on direct organogenesis or shoot development from meristematic cells [11].
![Figure 1: The developmental plant stages of common bean cv. Paulista on M.S. med. A: Seeds on M.S media; B: Seed swelling; C: Radical appearance; D: Cotyledon leaf appearance; E: Cotyledon leaf appearance; F: Seedlings; H: Galls on the root Some recent examples of direct organogenesis may be found in the literature [10]. Until now several types of cells, tissues and organs (cotyledonary nodes, embryonic axes, auxiliary shoots, cotyledon with split embryo axis, internodes, hypocotyls, leaves, leaf petioles or intact seedlings) have been used to induce the regeneration pathways [1,12,19]. Cotyledons with part of the embryonic axis as explained have been used to regenerate whole plants [20]. These protocols are very efficient.](/fulltextimages/10072/fig_1.png)
Figure 1: The developmental plant stages of common bean cv. Paulista on M.S. med. A: Seeds on M.S media; B: Seed swelling; C: Radical appearance; D: Cotyledon leaf appearance; E: Cotyledon leaf appearance; F: Seedlings; H: Galls on the root Some recent examples of direct organogenesis may be found in the literature [10]. Until now several types of cells, tissues and organs (cotyledonary nodes, embryonic axes, auxiliary shoots, cotyledon with split embryo axis, internodes, hypocotyls, leaves, leaf petioles or intact seedlings) have been used to induce the regeneration pathways [1, 12, 19]. Cotyledons with part of the embryonic axis as explained have been used to regenerate whole plants [20]. These protocols are very efficient.
Life cycle and Developmental Stages of M. incognita on Tissue Culture
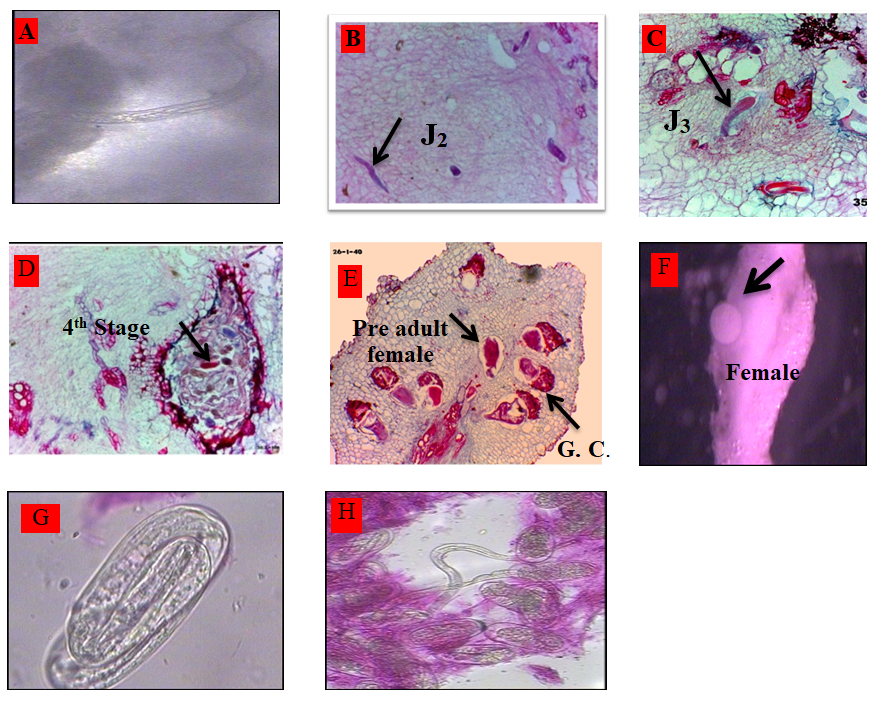
The life cycle of M. incognita completed about 37 days in root tissues of P. vuligaris cv. Paulista. Microscopic examinations of the infected bean roots revealed in Figure 2 as the following: Second stage juveniles of M. incognita penetrated the roots within 24 hr after nematode inoculation Figure 2A. Second–stage juvenile (j2) observed in roots 2-3 days after nematode inoculation Figure 2B. Third stage juveniles were observed in roots 6-8 days after nematode inoculation. Figure 2C. Fourth stage juveniles were observed in roots 12-14 days after nematode inoculation Figure 2D. The pre adult females were seen inside roots within 20 days after nematode inoculation Figure 2E. The adult females were seen inside roots within 27 days after nematode inoculation. Figure 2F. Egg laying females with deposit egg masses of M. incognita were observed 37 days after nematode inoculation.
Many investigators reported that there were many factors that affected the life cycle of Meloidogyne spp., the results obtained in the present study indicated that M. incognita, which reared on transforming the root culture was able to complete its life cycle on new transformed root tissue culture.
Data illustrated that M. incognita completed its life cycle in 37 days on full M.S medium. Second-stage juveniles of M. incognita penetrated the roots within 24 hr from inoculation. The adult females were seen inside roots within 27 days after nematode inoculation, and completed the life cycle in 37 days on full M.S medium. These results are in agreement with [21]. While [22] mentioned that Belonolaimus longicaudatus, completed three molts within 29 days on B5 medium. The benefit of using root tissue culture for studying the life cycle of M. incognita, in vitro as obligated indo parasite on bean plant, while encourage studying other life cycles of plant-parasitic nematodes either ecto or endo parasite in a laboratory and developing suitable medium for each taking into our consideration the difficulties of such studies in soil.
Conclusion
Root-knot nematode, Meloidogyne incognita completed its life cycle about thirty seven days in root tissues of Paulista cultivar on tissue culture media. Second-stage juveniles of M. incognita penetrated the roots within 24 hrs., and third- stage juveniles were observed in roots (six-eight) days after nematode inoculation. Fourth-stage juveniles were observed in roots 12-14 days, and the pre-adult females were seen inside roots within twenty days after nematode inoculation. The adult females were seen inside roots within twenty-seven days after nematode inoculation. While, egg laying females with deposit egg masses of M. incognita was observed thirty-seven days after nematode inoculation.
References
-
Delgado Sanchez P, Saucedo Ruiz M, Guzman Maldonado SH, Villordo Pineda E, Gonzalez Chavira M, et al. (2006) An organogenic plant regeneration system for common bean (Phaseolus vulgaris L.). Plant Sci 170 (4): 822-827.
-
Varisai Mohamed S, Sung JM, Jeng TL, Wang CS (2006) Organogenesis of Phaseolus angularis L.: high efficiency of adventitious shoot regeneration from etiolated seedlings in the presence of N6-benzylaminopurine and thidiazuron. Plant Cell Tissue and Organ Culture 86(2): 187-199.
-
Aragao FJL, Barros LMG, Brasileiro ACM, Ribeiro SG, Smith FD, et al. (1996) Inheritance of foreign genes in transgenic bean (Phaseolus vulgaris L.) co-transformed via particle bombardment. Theoretical and Applied Genetics 93(1): 142-150.
-
Veltcheva M, Svetleva D, Petkova SP, Perl P (2005) In vitro regeneration and genetic transformation of common bean (Phaseolus vulgaris L.) Problems and progress. Scientia Horticulturae 107(1): 2-10.
-
Tariq Khan M, Munir A, Mukhtar T, Hallmann J, Heuer H (2017) Distribution of root-knot nematode species and their virulence on vegetables in northern temperate agro ecosystems of the Pakistani-administered territories of Azad Jammu and Kashmir. J of Plant Dis. and Prot 124: 201-212.
-
Archana B, Saxena R (2012) Nematicidal effect of root extract of certain medicinal plants in control of Meloidogyne incognita in vitro and in vivo conditions. Pakistan J Nematology 30(2): 179-187.
-
Gine A, Monfort P, Sorribas FJ (2021) Creation and Validation of a Temperature-Based Phenology Model for Meloidogyne incognita on Common Bean. Plants 10(2): 240.
-
Nagl W, Ignacimuthu S, Becker J (1997) Genetic Engineering and Regeneration of Phaseolus and Vigna. State of the Art and New Attempts. Plant Physiology 150(6): 625-644.
-
Veltcheva MR, Lilova Svetleva D (2005) In vitro regeneration of Phaseolus vulgaris L. via organogenesis from petiole explants. J of Central European Agric 6(1): 53-58.
-
Ahmed E, Ahmed EE, Bisztray GYD, Velich I (2002) Plant regeneration from seedling explants of common bean (P vulgaris L) Acta Biologica Szegediensis 46(3-4): 27-28.
-
Arellano J, Fuentes S, Castilloespana P, Hernandez G (2009a) Regeneration of different cultivars of common bean (Phaseolus vulgaris L.) via indirect organogenesis. Plant Cell Tissue and Organ Culture 96 (1): 11-18.
-
Mahamune SE, Bansode RP, Sangle SM, Waghmare VA, Pandhur NB, et al. (2011) Callus induction from various explants of French bean (Phaseolus vulgaris L.). J of Experimental Sci 2(4): 15-16.
-
Zambre MA, De Clercq J, Vranova E, Van Montagu M, Angenon G, et al. (1998) Plant regeneration from embryo-derived callus in Phaseolus vulgaris L. (common bean) and P. acutifolius A. Gray (tepary bean). Plant Cell Reports 17(8): 626-630.
-
Mohamed MF, Coyne DP, Read PE (1993) Shoot organogenesis in callus induced from pedicel explants of common bean (Phaseolus vulgaris L.). J America Society Horticultural Science 118(1): 158-162.
-
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassay with tobacco. Physiol Plant 15(3): 473-497.
-
Koenning SR, Barker KR (1985) Genotobiotic Techniques for Plant Parasitic Nematodes. In: An Advanced Treatise on Meloidogyne, Barker, et al. (Eds.), Department of Plant Pathology, North Carolina State University, USA pp: 49- 66.
-
McBeth CW, Taylor AL, Smith AL (1941) Note on staining nematodes in root tissue. Proc Helm Soc Wash 8(1): 26.
-
Bybd Jr DW, Kirkpatrick T, Barker K (1983) An improved technique for clearing and staining plant tissues for detection of nematodes. J of Nematology 15(1): 142-143.
-
Hnatuszko Konka K, Kowalczyk T, Gerszberg A, Wiktorek Smagur A, Kononowicz AK (2014) Phaseolus vulgaris Recalcitrant potential. Biotechnology Advances 32(7): 1205-1215.
-
Dang W, Wei ZM (2009) High frequency plant regeneration from the cotyledonary node of common bean. Biologia Plantarum, 53(2): 312-316.
-
Verdejo S, Jaffee BA, Mankau R (1988) Reproduction of Meloidogyne javaniva on plant roots genetically transformed by Agrobacterium rhizogenes. J Nematol 20(4): 599-604.
-
Huang X, Becker O (1997) In vitro culture and feeding behavior of Belonolaimus longicaudatus on excised Zea mays roots. J Nematol 29(3): 411-415.
- Diversity of Candida sp and Antifungal Susceptibility Patterns in Digestive Candidiasis among People Living with HIV in CHU of Libreville, Gabon
- Vulvovaginal candidiasis: Retrospective study (2019- 2021) at the Centre Hospitalier National de Pikine, Suburban Dakar, Senegal
- Identification of Environmental Fungal Species in Clinical Services of University Hospital of Angre, Abidjan (Cote d’Ivoire)
- New Location of some Gasteroid Basidiomycetes in Western Kazakhstan
- Evaluation of Various Extracellular Enzymes of Ectomycorrhizal Mushrooms
- Morphology and Phylogeny of Lactarius Wallichianae sp. nov and Xerula magnispora sp. nov. from India