Effect of Introducing Foreign DNA into Nile Tilapia, Oreochromis Niloticus Gonads on Male Fertility of the Progeny Produced Under Salinity Stress
This work was conducted to study the effect of transfer foreign DNA isolated from sea bream, Sparus aurata or Artemia, Artemia salina into the ovaries and testes of O.niloticus adult on male fertility of the progeny produced under different salinity levels through studying the histological changes of testes. The results showed that male fertility of O. niloticus decreased as a result of introducing foreign DNA into the gonads of their parents. This decrease depends on the type of DNA since the results of the testes investigation revealed that O. niloticus received Artemia DNA displayed normal lobules of 40 %. In contrast, O. niloticus received sea bream DNA, showed only 28 % of normal lobules. The results also showed that the fertility of males decreased with increasing salinity levels up to 36 ppt. Although all fish received foreign DNA displayed low fertility, Nile tilapia received Artemia DNA showed more salinity tolerance and highest fertility than Nile tilapia received sea bream DNA.
Introduction
The scarcity in freshwater in many nations and its growing needs in agriculture and different city sports has multiplied the stress to expand aquaculture in brackish water and seawater [1]. Tilapia are essential species, mainly for tropical aquaculture and euryhaline fish, that may stay and thrive in a huge variety of salinity from freshwater to complete seawater despite the fact that a few species tolerate a wider Salinity variety than others [2, 3]. Tilapia (together with all species) is the second one maximum critical organization of farmed fish after carps and the maximum extensively grown of any farmed fish. In 2004 tilapia moved as much as the 8th maximum famous seafood withinside the USA worldwide manufacturing of all species of tilapia, and is projected to boom from 1.five million lots in 2003 to 2.five million lots through 2010, with a income price of extra than USD five billion. Most of this superior manufacturing is anticipated to be attributed to Nile tilapia [4] A most important purpose in introducing new genetic substances into the fish genome is to set up new, progressed business traces in aquaculture [5, 6]. Genetically changed fish provide new capability for elevated manufacturing of cultured organisms. This generation lets in the advent of recent trends, or development of antique ones, in a manner this is not possible to be achieved with conventional breeding methods [7]. Many species of genetically changed fish had been advanced for the reason that first genetically changed goldfish turned into suggested Aleström P, et al. [8]. The manufacturing of genetically changed fish has emerge as a famous method now no longer simplest for generating ideal trends however additionally for analyzing mechanisms of developmental law of diverse genes and gene promoters like quicker boom rate, sickness resistance, bloodless or salinity tolerance [5, 6]. Therefore, the goal of this paintings turned into to look at the impact of switch overseas DNA remoted from sea bream, Sparus aurata or Artemia, Artemia salina into the ovaries and testes of O. niloticus person on men fertility of the progeny produced beneathneath exclusive salinity ranges via look at the histological modifications of testes and Microsatellite DNA (ISSR) analysis.
Materials and Methods
The experimental work was undertaken at Fish farm and the laboratory of Breeding and Production of Fish, Animal and Fish Production Department, Faculty of Agriculture (Saba- Basha), Alexandria University, Alexandria, Egypt.
Fish Origin
The Nile tilapia used in this study descended from a randomly mating population at the private Fish Farm, El- Behera Governorate, Egypt.
Experimental Design
Preparation of Genomic DNA: A high molecular weight DNA was isolated according to the [9] method by reducing liver sample from sea bream, S. aurata and whole tissue sample of Artemia, A. salina. The extracted DNA was restricted by the Eco R1 restriction enzyme type II. It digested DNA between guanine and adenine [10]. Then, a 20 μg/0.1 ml/fish concentration was adjusted by extrapolating the dilutions for each type of DNA extracted using 0.1 x SSC buffers [11]. Injection of Genomic DNA into Fish Gonads: Adult Nile tilapia, O. niloticus with an average live weight (63.00 ±1.30 g/male and 55.50 ± 1.50 g/female), were chosen. The readiness of females to spawn was ascertained by examining the degree of swelling of the urogenital papilla [12]. Males were examined by stripping sperm [13]. Two males and four females were injected directly with the foreign DNA into fish gonads using a hypodermic needle; besides, the control group (two males and four females) was carried out. To inoculate the adult fish, the needle was inserted into the oviduct and sperm duct [11, 14]. Immediately after DNA treatments were carried out, each treated fish was stocked separately in fibreglass tanks (total volume 350 litre) supplied with dechlorinated water with adequate aeration. Brood fish were fed twice daily pellet containing 26% protein up to satiation for six days a week.
Base Generation (F0)
Culture Conditions: Base generation (F0) offspring produced from genetically modified Nile tilapia and their control were collected, counted and weighed. Fry was transferred separately to glass aquaria (100 x 34 x 50 cm) at a density of 1 fish/10 L and randomly divided for subsequent different salinity treatments. The glass aquaria were supplied with fresh dechlorinated tap water and aeration. Water temperatures were maintained at 28.00°C. Saline Water Acclimation: Two salinity levels (18 and 36 ppt) were prepared by mixing freshwater with crude natural salt [5] obtained from El-Nasr Company for salt, Borg El- Arab, Alexandria, Egypt, beside the third group of freshwater used as control. Fry obtained from each treatment of DNA, and their control was gradually acclimated to the respective salinities by raising the salinity at the rate of 4 ppt daily [15]. Moreover, the fourth group of each treatment of DNA and their control were transferred directly to 16 ppt. Water in each glass aquaria was partially changed once daily and every three days. Fries were fed three times daily with pellets containing 38% protein, to satiation, six days a week for 70 days. Then, all the fish were fed on a diet containing 32% to satiation, six days a week up to the end of the experiment (90 days). Fish were weighed and counted biweekly. A refractometer (S/Mill-E, ATAGO Co., LTD) was used to measure salinity.
Histological Characteristics of Gonads
By the end of the experiment, testes of ten males from each treatment were carefully removed and separately fixed in 10% formal saline solution. The fixed testes were washed in 70% ethyl alcohol for three days before dehydration, then cleared and embedded in paraffin wax for the histological study. Sections of 6-10 µm thick were stained with Eirlich hematoxylin and Eosin (H&E).
Microsatellite DNA (ISSR) Analysis
DNA was extracted from the muscle, liver and gonad tissues of the base generation (F0) of genetically modified Nile tilapia injected with Artemia DNA. In contrast, their control DNA was extracted from liver tissue only, following the method described by [9]. In this work, five base long oligonucleotide primers (Table 1) were used to initiate polymerase chain reaction (PCR) amplification. Primers were randomly selected based on GC content and annealing temperature for amplification. The PCR amplifications were performed following the procedure of [16, 17]. The reaction (25 µl) was carried out using 0.8 U of Taq DNA polymerase (Fanzyme), 25 pmol dNTPs and 25 pmol of random primer, 2.5 µl. 10X TaqDNA polymerase buffer and 40 ng of genomic DNA. The final reaction mixture was placed in a DNA thermal cycler (Eppendorf®). The PCR programme included an initial denaturation step at 94°C for 2 min followed by 45 cycles with 94°C for 30 s. For DNA denaturation, the annealing temperature of each primer, as indicated in Table 1, extends at 72°C for 30 s and finally extends at 72°C for 10 min. The samples were cooled at 4°C.
The amplified DNA fragments were separated on 1.5% agarose gel and stained with ethidium bromide. DNA marker (bp1000, 900 to 200) was used in this study. Moreover, to ensure that the amplified DNA bands originated from genomic DNA and not from primer artefacts, negative control (without DNA source) was carried out for each primer.
| Primers code | Sequences 5'-3' |
| 1 | GAGAGAGAGAGAGAGAT |
| 2 | CACACACACACAT |
| 3 | TATATATATATATATATAG |
| 4 | AGAGAGAGAGAGAGTG |
| 5 | AGAGAGAGAGAGAGTC |
Table 1: The sequences of the different primers used.
Results
Histopathological Findings
The ripe testes of O. niloticus manipulate male reared at freshwater had been composed specially of nests or lobules separated via way of means of interlobular tissue and confirmed greater energetic spermatogenesis in any respect degrees of improvement. Spermatozoa fill the lobular lumen (Figures 1a,b). The interlobular connective tissue is tremendously thin (6 mm in thickness) and wealthy in interstitial cells and blood capillaries. Also, the results confirmed marked asynchronous improvement in spermatogenesis at some point of the once a year reproductive cycle (Figure 1b). Treatment combination. The amplified sample become visualized on an UV trans illuminator and photographed via way of means of a Gel Documentation system.
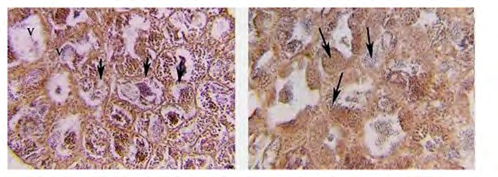
(a) (b) Figure 1a: Photomicrograph of cross-section (CS) in the testes of the control group of O. niloticus reared at freshwater, showing different stages of spermatogenesis within the sharp boundary somniferous lobules (arrows), vacuoles (v). (H and E) X 250. Figure 1b: Magnified section of the figure (1a) showing different stages of spermatogenesis (arrows). (H and E) X400.
Cross-phase of testes of O. niloticus manipulate male reared at 18 ppt of salinity confirmed that the proportion of ordinary lobules turned into 70% from overall cells. In the cross-phase of bizarre testes, spermatozoa had been detected with few numbers of spermatids. The sections had been characterised through the presence of discharge sperms and areas unfastened from permatogenesis as indicated in (Figure
2a, 2b). The testes of O. niloticus manage male, reared at 18 ppt of salinity, turned into taken into consideration as extra or much less much like ordinary tilapia; the proportion of similarity turned into approximately 70%. The interlobular connective tissue turned into ordinary in shape, besides a few tubules ruptures and disintegrated to shape big tubules (Figure 2b).
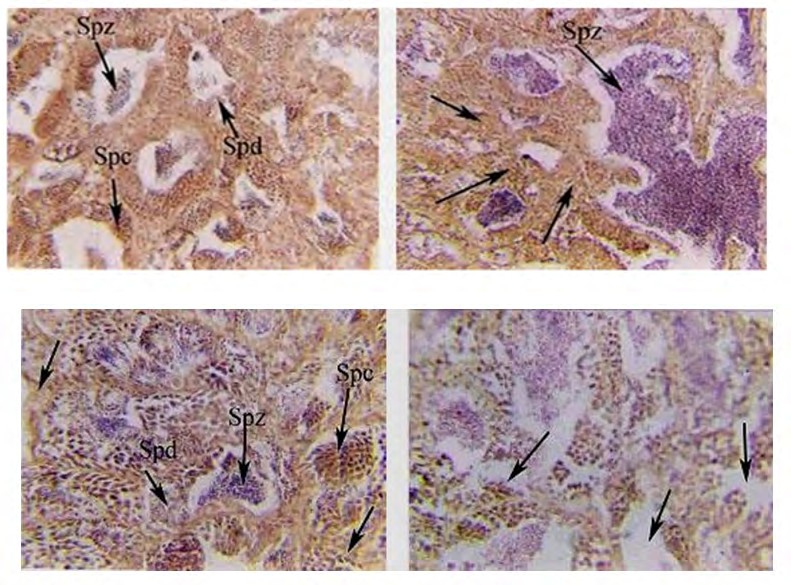
(2a) (2b) Figure 2a: Photomicrograph of cross-section (CS) in the testes of the control group of O. niloticus reared at 18 ppt of salinity, showing a different stage of spermatogenesis (spermatocyte (spc) and spermatid (spd)) and a moderate numberoftrapped sperms.(spz). (H and E) X250. Figure 2b: Magnified section of the figure (2a) showing undifferentiated connective tissues (arrows) and abnormal somniferous lobules with a large number oftrappedspermatozoa (spz). (H and E) X 400.
In the cross-segment of testes of O. niloticus manipulate male reared at 36 ppt of salinity percent of regular lobules became approximately 20% of the full lobules. In the cross- segment of deformed O. niloticus testes, all spermatogenesis ranges had been detected with hypertrophied characters. The collagenous connective tissue fibers had been detected in a few locations of the testes. These fibers elevated withinside the interlobular connective. Tissue in addition to pigmentation. Elongated nucleoli of connective tissue cells are commonly observed in affiliation with the fibers, as indicated in (Figure 3a). Other locations of the testes have been characterised with the aid of using the presence of vacuoles areas loose from spermatogenesis (no detection of secondary spermatocytes), and mild numbers of spermatozoa have been detected as indicated in (Figure 3b).
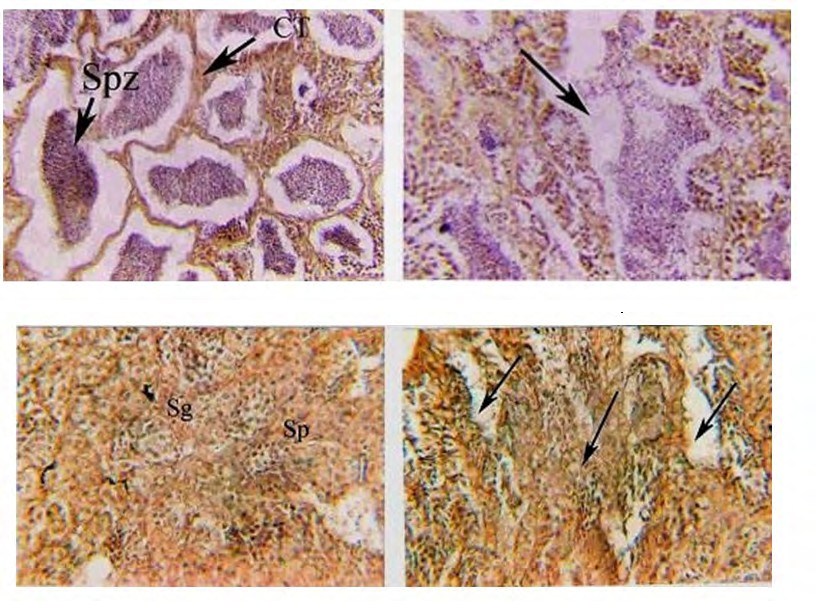
(3a) (3b) Figure 3a: Photomicrograph of cross-section (CS) in the testes of the control group of O. niloticus reared at 36 ppt of salinity, showing collagenous connective tissue fibers (arrows), spermatocyte (Spc), spermatid (Spd) and spermatozoa (Spz). (H and E) X 250. Figure 3b: Magnification of Fig. (3a) showing spaces free from spermatogenesis (arrows) and absence of interlobular connective tissue. (H and E) X400.
The percentage of normal lobules of genetically modified O. niloticus treated with sea bream DNA reared in freshwater was about 28% from total cells. The deformed testes in cross-section were characterized by some deflection of interlobular tissue from its original position. In some places, the interlobular tissues were absent, and in other places, the interstitial cells were active spermatogonia observed. Also, the testes were characterized with different spermatogenic activities, and the spermatozoa were discharged gradually from somniferous lobules as indicated in (Figure 4a). There are many elongated lobules without detectable margins (Figure 4b).
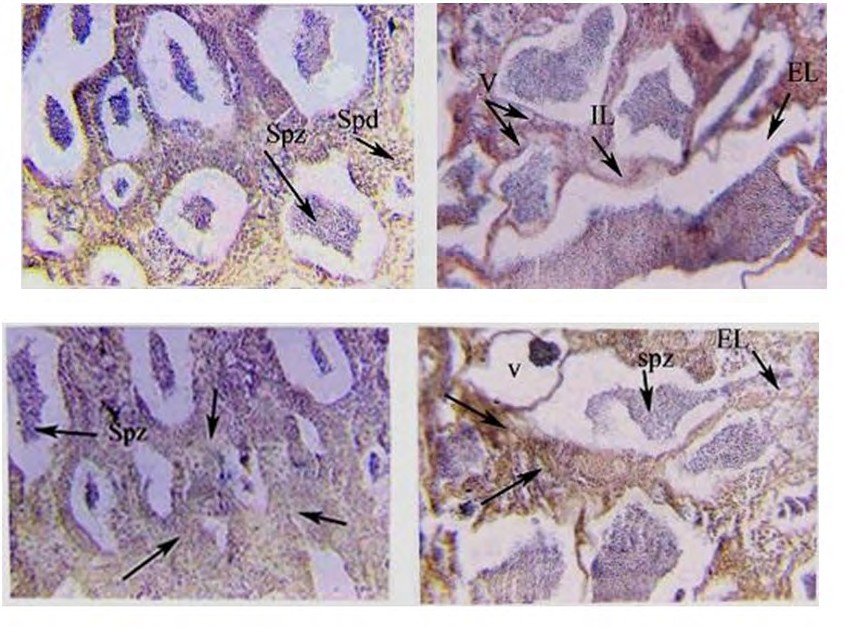
(4a) (4b) Figure 4a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with sea bream DNA and reared at freshwater, showing absence of interlobular tissue, a moderate number of spermatozoa (Spz) and spermatid (Spd). (H and E) X250. Figure 4b: Magnification of Fig. (4a), showing, deformed of Interlobular connective tissue (IL) and in some places absence of different stages of spermatogenesis, vacuoles (V), fibres and elongated lobules (EL). (H and E) X400.
In cross-section in testes of genetically modified O. niloticus treated with sea bream DNA reared at 18 ppt of salinity, the percentage of normal lobules was about 24% of the total lobules. The testes were characterized mainly by the presence of atretic lobules with abnormal and hypertrophy interlobular connective tissue. These lobules associated with more or less moderate or few numbers of spermatozoa, as shown in (Figure 5a). Other abnormal testes were characterized by increased thickness of interlobular connective tissue to reach about 78 µm and elongation of seminiferous (Figure 5b).A random distribution of lobules with Undifferentiated boundaries with the elongated nucleus of collagenous connective tissue fibrous. The only spermatozoon was detected in the thin seminiferous lobules, as indicated in (Figure 5b).
(5a) (5b) Figure 5a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with sea bream DNA and reared at 18 ppt of salinity, showing absence of interlobular tissue, a moderate number of spermatozoa (Spz) and spermatid (Spd). (H and E)X 250. Figure 5b: Magnification of Fig. (5a), showing, deformed of Interlobular connective tissue (IL) and in some places absence of different stages of spermatogenesis, vacuoles (V), fibers and elongated lobules (EL). (H and E) X400.
The percentage of normal lobules genetically modified O. niloticus, treated with sea bream DNA reared at 36 ppt of salinity was about 10% of the total cells. The deformed testes were characterized by increased thickness of interlobular connective tissue hypertrophied in some places (Figure 6a). In the section of testes, there were vacuoles and spaces free from spermatogenesis. The trapped spermatozoa were intermingled with undifferentiated tissue as indicated in (Figure 6b).
(6a) (6b) Figure 6a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with sea bream DNA and reared at 36 ppt of salinity, showing increase thickness of interlobular connective tissue (arrow) in the section and largely trapped spermatozoa (Spz ). (H and E) X 250. Figure 6b: Magnified section of figure (6a) showing elongated seminiferous lobules with a large amount of trapped sperms (arrows) (H and E) X 400.
In cross-section of testes of genetically modified O. niloticus treated with Artemia DNA reared at freshwater, the percentage of normal lobules was about 40% of the total lobules. Also, spermatogonia and primary spermatocytes were only detected. The deformed testes were characterized by the absence of interlobular connective tissue and other different stages of spermatogenesis. In addition, there were acuoles and spaces free from spermatogenesis as indicated in (Figures 7 a,b). The results showed that undifferentiated interlobular connective tissue marked atrophic changes were observed. The testicular capsule becomes thickened and fibrosed, which is considered a sign of infertility and shortage in the secretion of gonadal hormones.
(7a) (7b) Figure 7a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with Artemia DNA and reared at freshwater, showing: atretic lobules with only spermatogonia (Sg) and primary spermatocyte (sp) detected. (H and E) X 250. Figure 7b: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with Artemia DNA and reared at freshwater, showing spaces free from cells (arrows) and undifferentiated tissues (arrows). (H and E) X 400.
The percentage of normal lobules of genetically modified O. niloticus treated with ArtemiaDNA reared at 18 ppt of salinity was about 38% of the total cells from the light microscopic observation. The hypertrophy in some parts of interlobular connective tissue with an elongated nucleus and abnormal distribution and other treated testes were characterized with large elongated lobules with sharp boundaries (Figure 8a). Testes were characterized by a large number of spermatozoa cells and undifferentiated tissue in the cross-section. Seminiferous tubules were noticed with the discharged amount of sperms as indicated in (Figure 8b). Testes of genetically modified O. niloticus treated with Artemia DNA reared at 18 ppt showed a large number of trapped spermatozoa and disintegrated interlobular connective tissue with atretic spaces between the lobules as indicated in (Figure 8b).
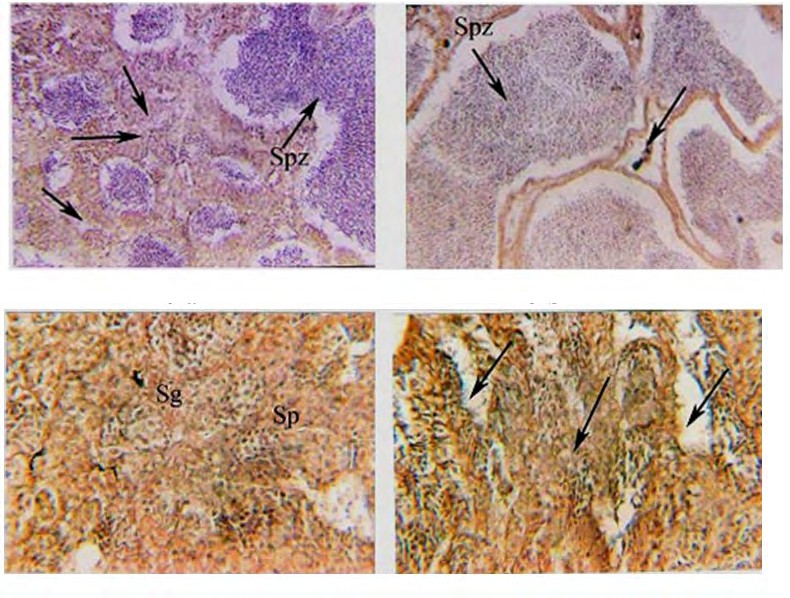
(8a) (8b) Figure 8a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with Artemia DNA and reared at 18 ppt of salinity, showing only a large amount of trapped ripe sperms in the seminiferous lobules (Spz) and hypertrophied interstitial connective tissues were detected (arrows) X 250. Figure 8b: Magnified section of fig (8a) showing elongated seminiferous lobules with sharp boundaries, trapped spermatozoa (Spz), atretic space between lobules. (H and E) X 400.
(9a) (9b) Figure 9a: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with Artemia DNA and reared in freshwater, showing: atretic lobules with only spermatogonia (Sg) and primary spermatocyte (sp) detected. (H and E) X 250. Figure 9b: Photomicrograph of cross-section (CS) in the testes of genetically modified O. niloticus treated with Artemia DNA and reared in freshwater, showing spaces free from cells (arrows) and undifferentiated tissues (arrows). (H and E) X 400.
The percentage of normal lobules of genetically modified O. niloticus treated with ArtemiaDNA reared at 36 ppt was about 18% of the total cells. A large number of hypertrophied were observed in the interlobular connective tissue (Figure 9a). Spermatozoa were moderately detected with complete degeneration of interlobular connective tissue (Figure 9b). Other parts of the testes were characterized by the absence of interlobular connective tissue, with thin tubular atretic seminiferous tubules (Figure 9b). Atretic tissue and vacuoles were detected in many spaces between spermatogenic cells (Figure 9a,b). Also, the results showed that hypertrophied or deformed interlobular connective tissue was considered a good sign of infertility and a shortage of gonadal hormones.
Genotype Analysis (ISSR Fingerprinting Analysis)
All DNA samples from genetically modified O. niloticus treated with different types of DNA and their control were examined using an ISSR marker. Five primers were used to determine DNA fingerprinting in genetically modified O. niloticus treated with Artemia- DNA and control fish. The results showed that no amplification was detected in the control reactions (without DNA source). All amplification products were reproducible when reactions were repeated using the same reaction conditions (Figure 10). The results also showed that the number of amplified bands detected varied, depending on the primers and DNA treatment. The highly genetic polymorphic percentage ranged from (11.20 to 34.20%) using different ISSR primers (Table 2 and Figure 10). It may be due to the differences in DNA molecule among normal and modified fish due to direct injection of foreign DNA isolated from Artemia. Moreover, some fragments of foreign DNA may be randomly integrated into O. niloticus genomes.
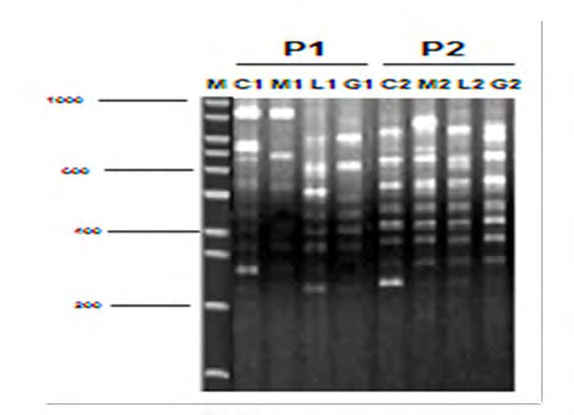
| Sample type | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Muscles | Liver | Gonads | |||||||||
| Total bands | Polymorphic bands | Polymorphism % | Total bands | Polymorphic bands | Polymorphism % | Total bands | Polymorphic bands | Polymorphism % | Total bands | Polymorphic bands | Polymorphism | |
| Primers | ||||||||||||
| 1 | 20 | 9 | 45 | 22 | 7 | 31 | 23 | 15 | 65 | 22 | 9 | 40 |
| 2 | 24 | 4 | 16 | 26 | 2 | 7 | 30 | 6 | 20 | 29 | 12 | 41 |
| 3 | 26 | 2 | 7 | 27 | 2 | 7 | 26 | 3 | 23 | 26 | 6 | 23 |
| 4 | 26 | 9 | 34 | 26 | 3 | 11 | 22 | 5 | 22 | 26 | 10 | 38 |
| 5 | 19 | 1 | 5 | 18 | 0 | 0 | 17 | 2 | 11 | 17 | 5 | 29 |
| Average % Polymorphm | 21.4 | 11.20 | 28.20 | 34.20 |
Table 2: Total amplified, Polymorphic, Monomorphic and Polymorphism % among Control, Muscles, Liver and Gonads of genetically mod
Discussion
Anthracyclines, Assem SS & SY EZ [18, 19] reported that the effects of direct intramuscular injection of foreign DNA into O. niloticus or T. zillii could be limited to germ cells of fish. Testes of males injected with various amounts of foreign DNA showed a large number of abnormalities. They add that further studies about the establishment of these effects on the following generations are needed. Moreover, Assem SS, et al. [20] stated that the effects of the transfer of foreign DNA into T. zillii gonads on the histological changes of testes of the first and second generations derived from fast- growing genetically modified redbelly tilapia were studied. The testes of F1 were hypertrophy in the interlobular connective tissue with abnormal distribution, while the second generation of F2, a large number of vacuoles with much atretic tissue, was observed. The abnormal tests of F2 showed a few spermatozoa and deformed interlobular connective tissue.
The present work results are consistent with these findings, since the abnormal shape of interlobular connective tissue was considered evidence of infertility and shortage in steroid hormone secretion as indicated by SY EZ [21]. Moreover, the damage of interlobular tissue was considered a good sign of infertility, and the fish cannot produce steroid needed for gametogenesis. Also, the abnormal of the interlobular connective tissue shows a shortage in function of these collagenous tissues [22]. El-Zaeem and Assem, et al. [18] reported that these collagenous connective tissue cells with elongated nuclei were usually associated with fibers. The results of the present work showed that undifferentiated interlobular connective tissue marked atrophic changes were observed. The testicular capsule becomes thickened, and fibrosis was considered a sign of infertility and shortage in the secretion of gonadal hormones. Cochran, et al. [23] postulated that the primary function of interlobular connective tissue is to produce steroid needed for gametogenesis and expression of secondary sexual characters. Fahmy, et al. [24] reported that any deformation in the interlobular connective tissue was considered as indication of disturbance and shortage in secretion of gonadal hormones in Chrysichthys rueppelli. Also, Hines, et al. [25] reported that abnormal shape of the interlobular connective tissue showed a shortage in function of these tissues and all the interlobular cells; these interstitial cells are involved in steroid synthesis carried out in steps. Also, the results showed that hypertrophied or deformed interlobular connective tissue was considereda good sign of infertility and shortage of gonadal hormones, as indicated by Cochran, et al. [23] and Hines,et al. [25], who postulated that the primary function of interlobular cells is to produce steroid needed for gametogenesis and expression of secondary sex characteristics. The present work results also showed that male fertility of O. niloticus decreased as a result of introducing foreign DNA into the gonads of their parents.
This decrease depends on the type of DNA described by [5], since the present work showed that O. niloticus received Artemia DNA and reared at freshwater displayed normal lobules of 40 %. In comparison, O. niloticus received DNA sea bream showed only 28 % of normal lobules. These results were consistent with the finding reported by Penman, et al. [26] that there was a trend towards increased mortality of rainbow trout eggs with increasing DNA concentration. Abdul Razak, et al. [27] reported that gonad somatic index (GSI) of transgenic Nile tilapia males had a lower value (0.0326) compared to non- transgenic siblings (0.0736). The present work results also showed that the fertility of males decreased with increasing salinity levels up to 32 ppt. Although all fish that received foreign DNA displayed low fertility, Nile tilapia received Artemia DNA showed more salinity tolerance and highest fertility than Nile tilapia received sea bream DNA. In this connection, El-Zaeem, et al. [5, 28] stated that increasing salinity levels led to dropped Gonadosomatic index and nonsuccess of reproduction. Also, Fineman-Kalio, et al. [29] indicated that spawning of O. niloticus was inhibited by high salinity, and the gonad development and spawning occurred at salinities of 17-19 ppt. With increasing water salinity from 25 to 30 ppt, the onset of reproduction was delayed while reproduction stopped completely at salinity above 30 ppt. The specific characterization of the RAPD method (random, uncharacterized multiple genome loci; dominant nature of markers; and the possibility of migration of non-homologous bands) result in limitations based on RAPD analysis alone. Despite these limitations, the RAPD analysis can be used effectively to assess genetic variation among fish species [30]. The main advantages of RAPD and ISSR markers are the possibility of working with anonymous DNA, relatively low expense, fast and simple to produce [31].
The results obtained by El-Zaeem, et al. [28] suggested that hypersaline genetically modified O. niloticus with a higher growth rate which can be produced by the transfer of a foreign DNA isolated from sea bream and Artemia as a feasible and fast methodology compared to interspecific hybridization, which is one classical breeding methods [5]. According to these results, genetically modified O. niloticus treated with sea bream and Artemia-DNA surpassed most of the productive performance traits under different salinity levels up to 3 ppt. The present work results are consistent with these findings since the present work showed that O. niloticus received Artemia DNA and reared at freshwater displayed normal lobules of 40 %. In comparison, O. niloticus received DNA sea bream showed only 28 % of normal lobules. El-Zaeem, et al. [32] recommends using foreign DNA transformation to induce sterility in Nile tilapia males and therefore remedy the trouble of overpopulation in business pond cultures. The only limitation of this methodology may be the time required to handle and inject the broodstock. The outcomes of the existing paintings are steady with those findings.
Finally, the results showed that males’ fertility decreased with increasing salinity levels up to 36 ppt. This matched with Barman HK, et al. [33] that reported the high salinity affected the hematological, expression of some genes and histological features of Nile tilapia.
Conclusion
Our result revealed that the fertility of males reduced with rising salinity levels up to 36 ppt. in addition; O. niloticus received Artemia DNA displayed normal lobules of 40 %. In contrast, O. niloticus received DNA sea bream showed only 28 % of normal lobules. Corresponding to these results, genetically modified O. niloticus received sea bream, and Artemia-DNA outstripped most of the productive performance traits under different levels of salinity up to 3 ppt.
References
-
El-Sayed AFM (2006) Tilapia culture in salt water: environmental requirements, nutritional implications and economic potentials. Avances en Nutricion Acuicola pp: 1-12.
-
Güner Y, Özden O, Çağirgan H, Altunok M, Kizak V (2005) Effects of salinity on the osmoregulatory functions of the gills in Nile tilapia (Oreochromis niloticus). Turk J Vet Anim Sci 29: 1259-1266.
-
Kamal AHMM, Mair GC (2005) Salinity tolerance in superior genotypes of tilapia, Oreochromis niloticus, Oreochromis mossambicus and their hybrids. Aquaculture 247(1-4): 189-201.
-
Austin K, Mohammed A (2013) An Econometric Analysis of Factors from the Farmers’ perspective That Affect Profitability of Tilapia Production in Trinidad. Ag Econ pp: 1-16.
-
El-Zaeem SY, Ahmed MMM, Salama ME, El-Maremie HA (2011) Production of salinity tolerant Nile tilapia, Oreochromis niloticus through traditional and modern breeding methods: II. Application of genetically modified breeding by introducing foreign DNA into fish gonads. African Journal of Biotechnology 10(4): 684-695.
-
El-Zaeem SY (2012) Extraordinary Mullet growth through direct injection of foreign DNA. African Journal of Biotechnology 11(33): 8295-8301.
-
Aleström P, de la Fuente J (1999) Genetically modified fish in aquaculture:Technical, environmental and management considerations. Biotecnologia Aplicada 16: 127-130.
-
Zhu Z, He L, Chen S (1985) Novel gene transfer into the fertilized eggs of gold fish (Carassius auratus L. 1758). Journal of Applied Ichthyology 1(1): 31-34.
-
Bardakci F, Skibinski D (1994) Application of the RAPD technique in tilapia fish: species and subspecies identification. Heredity (Edinb) 73(Pt 2)_:_ 117-123.
-
Wu JJT, Lu AL (1994) Escherichia coli mutY-dependent mismatch repair involves DNA polymerase I and a short repair tract. Mol Gen Genet 244: 444-450.
-
El-Zaeem S (2001) Breeding studies in Tilapia. Faculty of Agriculture (Saba-Basha), Alexandria University, Egypt.
-
Hussain MG, Chatterji A, McAndrew BJ, Johnstone R (1991) Triploidy induction in Nile tilapia, Oreochromis niloticus L. using pressure, heat and cold shocks. Theor Appl Genet 81(1): 6-12.
-
Wester RC, Foote RH (1972) Ouabain effect on bovine spermatozoan motility and testosterone binding 141(1): 26-30.
-
Lu JK, Fu BH, Wu JL, Chen TT (2002) Production of transgenic silver sea bream (Sparus sarba) by different gene transfer methods. Mar Biotechnol 4(3): 328-337.
-
Watanabe WO, Kuo CM (1985) Observations on the reproductive performance of Nile tilapia (Oreochromis niloticus) in laboratory aquaria at various salinities. Aquaculture 49(3-4): 315-323.
-
Williams JG, Hanafey MK, Rafalski JA, Tingey SV (1993) [51] Genetic analysis using random amplified polymorphic DNA markers. Methods in Enzymology 218: 704-740.
-
Williams JG, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV (1990) DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18(22): 6531- 6535.
-
Assem SS, El-Zaeem SY (2005) Application of biotechnology in fish breeding. II:production of highly immune genetically modified redbelly tilapia, Tilapia zillii. African Journal of Biotechnology 4(5): 449-459.
-
SY EZ (2004) Alteration of the productive performance characterestics of Oreochromis niloticus and Tilapia zillii under the effect of foreign DNA injection. Egypt J Aquat Biol Fish 8: 261-278.
-
El-Zaeem S, Assem S (2006) Evaluation of the first and second generations delivered from fastgrowing genetically-modified Tilapia zillii: Histological and genetic approach. J Appl Sci Res 7: 364-374.
-
Sulistyo I, Fontaine P, Rinchard J, Gardeur JN, Migaud H, et al. (2000) Reproductive cycle and plasma levels of steroids in male Eurasian perch Perca fluviatilis. Aquat Living Resour 13(2): 99-106.
-
Rosenblum PM, Pudney J, Callard IP (1987) Gonadal morphology, enzymehistochemistryand plasma steroid levels during the annual reproductive cycle of male and female brown bullhead catfish, Ictalurus nebulosus Lesueur. J Fish Biol 31: 325-341.
-
Cochran RC (1992) In vivo and in vitro evidence for the role of hormones in fish spermatogenesis. J Exp Zool 261(2): 143-150.
-
Fahmy AFA (2006) Reproductive biology and induced spawning of one species of family Bagridae pp: 241.
-
Hines GA, Boots LR, Wibbels T, Watts SA (1999) Steroid Levels and Steroid Metabolism in Relation to Early Gonadal Development in the TilapiaOreochromis niloticus (Teleostei: Cyprinoidei). Gen Comp Endocrinol 114(2): 235-248.
-
Penman DJ, Beeching AJ, Penn S, Maclean N (1990) Factors affecting survival and integration following microinjection of novel DNA into rainbow trout eggs. Aquaculture 85(1): 35-50.
-
Growth performance and gonadal development of growth enhanced transgenic tilapia Oreochromis niloticus (L.) following heat- shock-induced triploidy. Mar Biotechnol 1(6): 533-544.
-
El-Zaeem SY (2011) Growth comparison of Nile tilapia (Oreochromis niloticus) and Blue tilapia, (Oreochromis aureus) as affected by classical and modern breeding methods. African Journal of Biotechnology 10(56): 12071-12078.
-
Kalio AF (1988) Preliminary observations on the effect of salinity on the reproduction and growth of freshwater Nile tilapia, Oreochromis niloticus (L.), cultured in brackishwater ponds. Aquac Res 19_:_ 313-320.
-
Barman HK, Barat A, Yadav BM, Banerjee S, Meher PK (2003) Genetic variation between four species of Indian major carps as revealed by random amplified polymorphic DNA assay. Aquaculture 217(1-4): 115- 123.
-
El-Dahhar AA (2007) Animal and Fish Production Department Faculty of Agriculture (Saba Basha) Alexandria University.
-
El-Zaeem SY (2015) A novel method for inducing sterility in Nile tilapia males.Aquac Res 46(3): 765-768.
-
Mohamed NA, Saad MF, Shukry M, El-Keredy AMS, Nasif O, et al. (2021) Physiological and ion changes of Nile tilapia (Oreochromis niloticus) under the effect of salinity stress. Aquaculture Reports 19: 100567.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review