Evolutionary History of Procyonids (Carnivora: Procyonidae): Comparative Cytogenetics of Nasua Nasua and Procyon Cancrivorus
The chromosome number of all species of Procyonidae was very conserved. Most have very strong chromosomal homology among members of the family, as well as with different species of the suborder Feliformia. Many chromosomes have similar and sometimes identical G-band patterns, while others differ due to Robertsonian translocations and inversions. In this work, the karyotype of two species of the family Procyonidae, Nasua nasua (2n=38; NF = 74) and Procyon cancrivorus (2n=38; NF = 76), the latter described cytogenetically for the first time, is characterized using conventional staining and differential G, C and NOR banding techniques. Pericentric and paracentric inversions were observed in Procyon cancrivorus and Nasua nasua, respectively. Finally, the G-band karyotypes of both species were compared with the Ancestral Carnivore Karyotype proposed in the literature, providing a homology and conservation on the karyotype evolution of this order.
Introduction
Belonging to the genus Procyon, P. cancrivorus (Cuvier) is a solitary animal of nocturnal habits, living near bodies of water. It has a short, dense coat of dark brown to gray color, easily identified by a dark face mask. Its distribution includes Central and South America [2]. Exclusive to South America, Nasua nasua (Linnaeus) is a social animal of daytime habits, and can live in large groups. Its color varies according to the geographical distribution, between reddish and brown tones, and has great mobility in its long snout. Both P. cancrivorus and N. nasua are medium-sized animals with a long ringtail, presenting sexual dimorphism with males larger than females [2].
The procyonids described karyotypically so far are very similar, presenting 2n=38, they are difficult to differentiate only on the basis of chromosome morphology. In addition to the similarities between those belonging to the family, there are great resemblances with the Felids, but there is no evidence to establish a close relationship between them [3]. So far, karyotypes with G-banding of four species have been described, Bassariscus astutus (Lichtenstein), Bassaricyon gabbii (Allen), Procyon lotor (Linnaeus) and Potos flavus (Schreber). The karyotypes of these species have the same chromosome number (2n = 38) and their structural characteristics and patterns of Banding G and NOR’s are very similar [4]. They share great chromosomal homology with vivérrids and even with mustelids, but essentially with felines. P. lotor and B. astutus share 15 homologous autosomes with Felines [5]. The karyotype and banding pattern G of P. lotor and B. astutus is exactly the same, and both share the same number and chromosomal morphology with ACK [4, 6].
The G-banding is one of the main techniques for cytogenetic investigations in mammals, by precise chromosomal identification and characterization, allowing analysing rearrangements and polymorphisms in a large database, through the pattern of positive and negative bands observed on chromosomes [7, 8]. Beyond the G-banding, other techniques are used to make cytogenetic correlations between species, such as the NOR-banding, a technique that allows the identification of nucleolar organizing regions, also widely used in mammalian studies [9, 10]. Additionally with the classical techniques of banding, other comparisons are necessary for a better understanding of the evolutionary history of the group. The ancestral carnivore karyotype (ACK) was developed from conventional banding and chromosomal painting of various carnivorous species, making itself the best reference for chromosomal investigations in this order [11].
Of the 14 species that make up this New World taxonomic group, P. cancrivorus is the only one that has not been cytogenetically described. In order to get a more comprehensive understanding of the mechanisms underlying the chromosomal evolution of Procyonids species, we used conventional cytogenetic methodologies on Nasua nasua and Procyon cancrivorus. These methods were used to examine their karyotype structure, distribution of heterocromatin constitutive, nucleolus organizer region. In addition, G-banding was done on the species, highlighting some inversions.
Material and Methods
Animals
The Ecological Park depends on the under secretariat of Ecology and Sustainable Development of the Ministry of ecology of Misiones Argentina. “El Puma” is a natural area with the category of multipurpose reserve dedicated to research, self-sufficient production of native flora and fauna, natural spaces suitable for outdoor recreation and experimental sites for models of sustainable use of natural resources. Blood samples were obtained from confiscated or rehabilitated animals that were in the “El Puma” Ecological Park during the years 2008 to 2012. Chromosome collection was performed using long term lymphocyte cultures by protocol [12]. Species of the Order Carnivora belonging to the family Procyonidae were chromosomally analyzed, seven males and three females of N. nasua, and three males of P. cancrivorus.
Chromosomal Preparations and Classic Cytogenetics
Metaphases were obtained from preparation of long- term lymphocyte cultures, were performed in PBMax Medium (Gibco) for 72 hours according to Moorhead PS, et al. [12] with modifications. The metaphases obtained were analyzed using conventional techniques and the distribution of heterochromatin blocks are analyzed by C-banding, according to the protocol proposed by Perelman PL, et al. [13] and NOR according to Howell WM, et al. [14]. For the photomicrography was used an Olympus Cx31 microscope with a built-in camera, Olympus model E-330. The 40 best metaphases of each species were chosen and photographed. The assembly of the karyotypes was carried out using the GIMP program and were assembled according to Levan A, et al. [15]. To perform the biometric analysis, the software MICROMEASURE 3.3 ©.
Results
The individuals analyzed presented the same diploid number 2n=38 chromosomes. N. nasua with a fundamental number NF=74 (36 bi-armed chromosomes and 2 telocentric chromosomes), while all P. cancrivorus chromosomes are bi- armed, NF=76 (Figures 1 and 2).

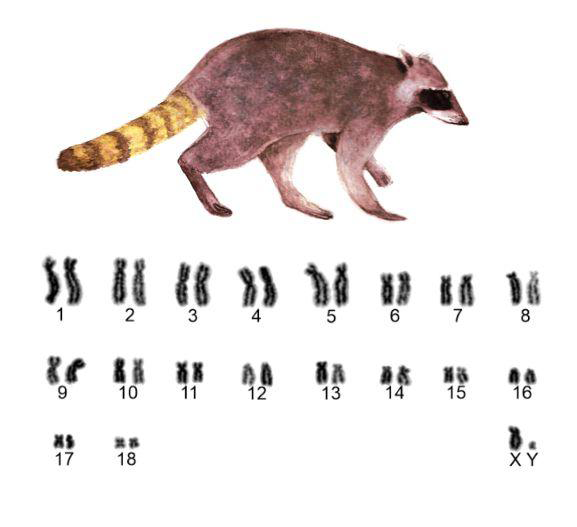
The centromeric index (CI), the standard error (SE) and the chromosomal morphology are shown in Table 2. In both species the pairs 1, 2, 6, 9, 13 and 14 are submetacentric (sm), par 5 is subtelocentric (st), pairs 3, 4, 17 and 18 are metacentric (m), the latter two representing the smallest chromosomes of this autosomal complement. As for the differences, N. nasua presents pairs 7, 8, 11 and 12 submetacentric (sm), subtelocentric pair 10 (st), metacentric pair 15 (m), finally, par 16 representing the only telocentric autosome pair of the studied species. P. cancrivoros, presents the pairs 10, 11 and 15 submetacentric (sm), pairs 7, 8, 12 and 16 subtelocentric (st). In the pair 15 of N. nasua and P. cancrivoros, secondary constrictions are clearly observed. The sexual pair is also similar in both species, with the x chromosome of the submetacentric type (sm), and the telocentric Y chromosome (t).
| N° chromosomes | Nasua nasua | Procyon cancrivorus | ||||
|---|---|---|---|---|---|---|
| CI | SE | CM | IC | SE | CM | |
| 1 | 0,29 | 0,03 | sm | 0,25 | 0,00 | sm |
| 2 | 0,33 | 0,03 | sm | 0,25 | 0,02 | sm |
| 3 | 0,41 | 0,05 | m | 0,43 | 0,01 | m |
| 4 | 0,42 | 0,04 | m | 0,45 | 0,03 | m |
| 5 | 0,21 | 0,12 | st | 0,22 | 0,02 | st |
| 6 | 0,34 | 0,07 | sm | 0,30 | 0,00 | sm |
| 7 | 0,32 | 0,07 | sm | 0,19 | 0,01 | st |
| 8 | 0,29 | 0,05 | sm | 0,21 | 0,03 | st |
| 9 | 0,29 | 0,09 | sm | 0,35 | 0,02 | sm |
| 10 | 0,22 | 0,05 | st | 0,34 | 0,01 | sm |
| 11 | 0,34 | 0,08 | sm | 0,36 | 0,05 | sm |
| 12 | 0,33 | 0,04 | sm | 0,19 | 0,01 | st |
| 13 | 0,33 | 0,04 | sm | 0,30 | 0,03 | sm |
| 14 | 0,34 | 0,04 | sm | 0,32 | 0,01 | sm |
| 15 | 0,38 | 0,05 | m | 0,36 | 0,03 | sm |
| 16 | 0,11 | 0,00 | t | 0,25 | 0,03 | st |
| 17 | 0,38 | 0,14 | m | 0,40 | 0,02 | m |
| 18 | 0,40 | 0,07 | m | 0,45 | 0,02 | m |
| X | 0,34 | 0,07 | sm | 0,33 | 0,01 | sm |
| Y | 0,18 | 0,00 | t | 0,00 | 0,00 | t |
Table 1: ** Biometric analysis of Nasua nasua and Procyon cancrivorus according to the nomenclature of Guerra, 1988 [8].
CI=centromeric index; SE=standard error; CM=chromosomal morphology; m=metacentric; sm=submetacentric; st=subtelocentric; t=telocentric. Table 1: Biometric analysis of Nasua nasua and Procyon cancrivorus according to the nomenclature of Guerra, 1988 [8].
NOR-banding
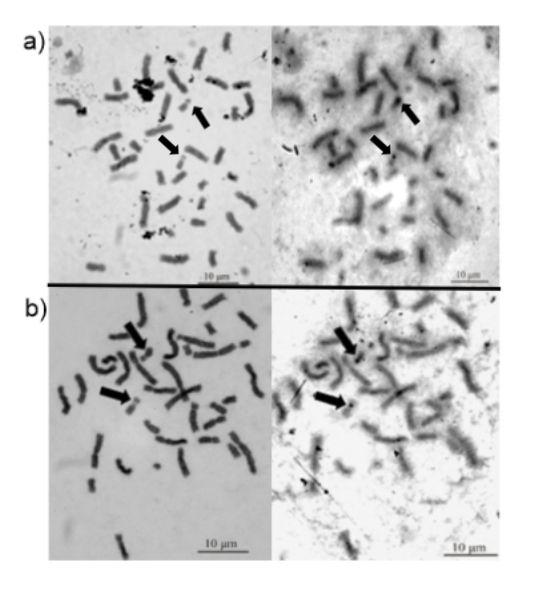
In order to identify the organizing region of the nucleolus and its position in chromosomes, the silver nitrate technique was used. The analysis obtained in the bands of N. nasua and P. cancrivorus allowed us to observe that the secondary constrictions of pair 15 coincide with the organizing regions of the nucleolus (Figure 3).
C-banding
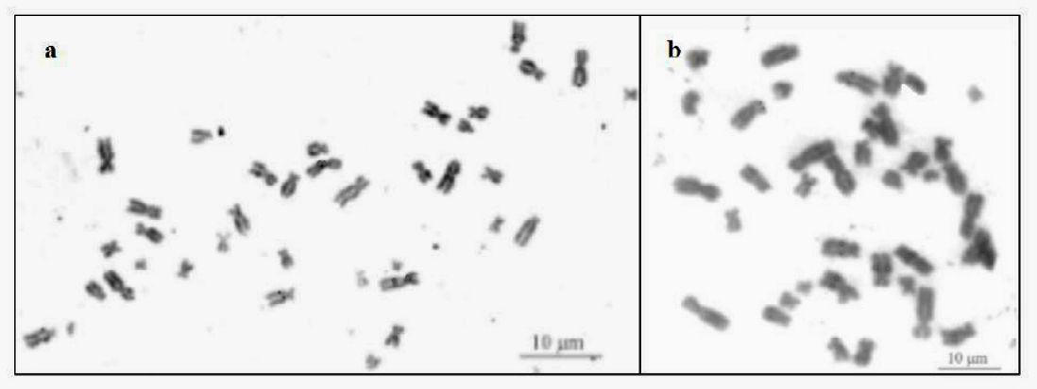
The C-banding technique demonstrated regions rich in heterochromatin in the interstitial regions of chromosomes 6, 7 and 9 in both species, in addition to chromosome 5 and 10 in N. nasua. The chromosome 5 of P. cancrivorus presents regions rich in heterochromatin in its pericentromeric. The X chromosome of N. nasua showed a positive C mark in the centromeric region. However, in P. cancrivorus, no positive C blocks were observed on sex chromosomes (Figure 4).
G-banding
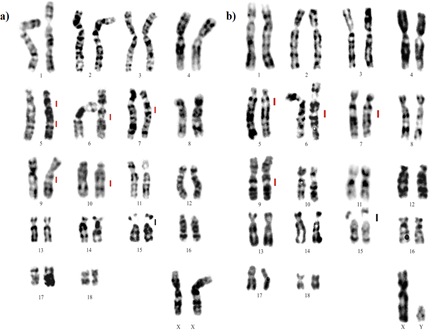
The G-band pattern was much conserved in most chromosomes, compared to others belonging to the family. Each species had an exception; N. nasua suffered a paracentric inversion of the short arm of the par 9, while P. cancrivorus underwent a pericentric inversion, modifying the shape of the par 16. (Figure 5).
Discussion
The chromosomal structure and organization of two representative mammals species (Procyonidae) were studied, with a focus on their karyotype description and classical cytogenetic. Mammal’s’ evolutionary history has produced an amazing group with diverse adaptations and a vast number of species. Chromosomes of numerous species of Carnivores have been analyzed extensively with comparative conventional banding [16, 17].
The ACK proposed by Pinthong K, et al. [11] based on the karyotype and banding pattern of four different families has been used as the closest model to the ancestral karyotype of the order Carnivora. All comparisons between these taxa will be made on the basis of this karyotype (Figure 6).
![Figure 6: ACK proposed by Pinthong K, et al. [11], comparing with Nasua nasua (NNA) and Procyon cancrivorus (PCA). The ACK is a composite karyotype, whose chromosomes were selected from those of Felis catus (Felidae) and Basariscus astutus (Procyonidae). Chromosomes with an inversion relative to the ACK are indicated by arrows (indicating the breakpoint). The red vertical line indicates heterochromatin-rich regions and the black lines the location of nucleolus-organizing regions.](/fulltextimages/9725/fig_6.png)
Figure 6: ACK proposed by Pinthong K, et al. [11], comparing with Nasua nasua (NNA) and Procyon cancrivorus (PCA). The ACK is a composite karyotype, whose chromosomes were selected from those of Felis catus (Felidae) and Basariscus astutus (Procyonidae). Chromosomes with an inversion relative to the ACK are indicated by arrows (indicating the breakpoint). The red vertical line indicates heterochromatin-rich regions and the black lines the location of nucleolus-organizing regions.
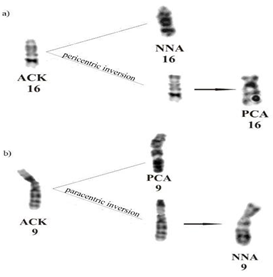
The karyotype of P. cancrivorus (PCA) is the one that has undergone a pericentric inversion on chromosome 16 thus modifying the NF of its karyotype (Figure 7a). Although NNA maintains the standard karyotype in relation to ACK, by G-banding it can be observed that it has undergone a paracentric inversion on chromosome 9 (Figure 7b). These two simple inversions distinguish the N. nasua (NNA) and the P. cancrivorus (PCA) from the ACK. Both species showed the same chromosome number characteristic of the order, but differ from each other in the fundamental number; this difference is based on the change in the morphology of one of their chromosomes caused by a pericentric inversion. N. nasua (NNA) maintains the same number, morphology and arrangement as the ACK proposed by Pinthong K, et al. [11].
No differences were observed regarding the position of the nucleolus organizer region, in both species the NOR band was located on chromosome pair 15 just at the secondary constriction of the short arm. This chromosome is considered ancestral as it is repeated in most carnivore families, with subtle variations in some species. Within the suborder Caniformia the morphology of this chromosome is quite diverse; however in the procyonids this chromosome characteristic of the order presents greater stability. Wurster- Hill and Bernirshke [3] defined this chromosome present in all procyonids as a characteristic marker of carnivores, identical to the E1 pair of felids.
Both species of procyonids analyzed in this work were carriers of C bands in chromosome pairs 5, 6, 7 and 9 in interstitial position, except in P. cancrivorus which on par 5 had a single C-band in the pericentromeric region and N. nasua which had an extra mark on pair 12 and sex chromosome X. As in the species analyzed by Pathak S, et al. [18], none presented positive C marks on the Y chromosome.
In most mammalian species the C-bands are centromeric, but other terminal, interstitial or whole-arm C-banding patterns are also frequent in the different orders. From the carnivore species analyzed so far, two general conclusions can be formulated in this regard: (1) compared to other mammalian orders, carnivores possess much less constitutive heterochromatin in their genomes; (2) contrary to most mammals, some carnivores possess very little or no centromeric heterochromatin [18, 19]. The only carnivore species that exhibit large blocks of heterochromatin in the centromeres and pericentromeric regions of submetacentric chromosomes are the canids and ursids. This distinctive feature of these two families is a consequence of their great karyotypic diversity resulting from different types of structural rearrangements. Probably the most important source of karyotypic variability in canids and ursids is related to the amount of heterochromatin and its distribution pattern detectable by C-banding techniques [20].
Only two species of the family Procyonidae have been analyzed by C-banding; B. astutus and P. lotor both showed insignificant amounts of centromeric heterochromatin and a large positive C-block on the short arm of the small subtelocentric pair [18]. With so little constitutive heterochromatin, however, it is conceivable that karyotypic changes, such as Robertsonian fusions, may be more difficult to achieve. This may partly explain the remarkable stability observed in procyonid chromosomes, as well as their conserved G-band pattern with the other carnivore families.
Conclusions
The species Procyon cancrivorus, described for the first time in this study, corroborates the results obtained in the other species analyzed, showing a highly conserved karyotype and identical G, C and NOR band patterns. Although the Family Felidae is not closely related to the Procyonidae family, both show the most conserved karyotype of the order Carnivora. Finally, N. nasua and P. cancrivorus, focus species of the study, present almost complete homology compared to the ACK proposed in the literature, 17 of the 19 chromosomal pairs of both species share the same pattern of G bands between them and the ACK, evidencing once again how conserved is the karyotype of this family.
Acknowledgments
We are grateful to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for supporting this research. The authors are grateful to all our colleagues at the Laboratório de Diversidade Genética Animal from the Universidade Federal do Pampa (RS, Brazil) and Parque Ecológico El Puma (Subsecretaría de Ecología y Desarrollo Sustentable), Argentina. We are also grateful to Guilherme Castro Franco de Lima for the illustration of the Nasua nasua and Procyon cancrivorus used in Figures 1 and 2.
References
-
Cubas ZS, Silva JCR, Catão-Dias JL (2014) Treatise on Wild Animals: Veterinary Medicine. Rock, (2nd Edn.) Sao Paulo.
-
Cheida CC, Nakano-Oliveira E, Fusco-Costa R et al. (2006) Ordem Carnivora. In: Reis N, Peracchi A L, Pedro WA & Lima IP (Eds), Mamíferos do Brasil. Edifurb, Londrina, pp: 261-265.
-
Wurster DH, Benirschke K (1968) Comparative cytogenetic studies in the order Carnivora. Chromosoma 24(3): 336-382.
-
O’brien SJ, Menninger JC, Nash WG (2006) Atlas of mammalian chromosomes. John Wiley & Sons. 446-509.
-
Wurster DH, Gray CW (1975) The interrelationships of chromosome banding patterns in procyonids, viverrids, and felids. Cytog Gen Res 15(5): 306-331.
-
Stanyon R, Bigoni F, Wienberg J, Hadidian J (1993) A standardized G‐banded karyotype for the raccoon (Procyon lotor) compared with the domestic cat. Ital J of Zool 60(1): 41-45.
-
Burgos M, Jiménez R, De La Guardia RD (1986) A Rapid Simple and Reliable Combined Method for G-Banding Mammalian and Human Chromosomes. Stain Technol 61( 5): 257-260.
-
Fredga K, Narain Y (2000) The complex hybrid zone between the Abisko and Sidensjö chromosome races of Sorex araneus in Sweden. Biol J Linn Soc 70(2): 285-307.
-
Supanuam P, Tanomtong A, Khunsook S, Sangpakdee W, Pinthong K, et al. (2012) Localization of Nucleolar Organizer Regions (NORs) of 4 Gibbon Species in Thailand by Ag-NOR Banding Technique. Cytologia 77(2): 141-148.
-
Pinthong K, Tanomtong A, Khongcharoensuk H, Chaiphech S, Rattanayuvakorn S, et al. (2017) Karyotype and idiogram of Indian hog deer (Hyelaphus porcinus) by conventional staining, GTG-, high-resolution and Ag- NOR banding techniques. Cytologia 82: 227-233.
-
Nash WG, Menninger JC, Padilla-Nash HM, Gary Stone, Polina L. Perelman, et al. (2008) The Ancestral Carnivore Karyotype (2n= 38) Lives Today in Ringtails. Journal of heredity 99(3): 241-253.
-
Moorhead PS, Nowell PC, Mellman WJ, Battips DM, Hungerford DA (1960) Chromosome preparations of leukocytes cultured from human peripheral blood. Exp. cell Res 20: 613-616.
-
Seabright Marina (1971) A rapid banding technique for human chromosomes. Lancet 2(7731): 971-972.
-
Howell WM, Black DA (1980) Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 36(8): 1014– 1015.
-
Levan A, Fredga K, Sandberg AA (1964) Nomenclature for centromeric position on chromosomes. Hereditas 52(2): 201-220.
-
Perelman PL, Graphodatsky AS, Serdukova NA, Nie W, Alkalaeva EZ, et al. (2005) Karyotypic conservatism in the suborder Feliformia (Order Carnivora). Cytogen and gen res 108(4): 348-354.
-
Ruiz-García M, Jaramillo MF, López JB, Rivillas Y, Aurita Bello, et al. (2021) Mitochondrial and karyotypic evidence reveals a lack of support for the genus Nasuella (Procyonidae, Carnivora). J Vert Biol 71(21040): 21040- 21041.
-
Pathak S, Wurster-Hill DH (1977) Distribution of constitutive heterochromatin in carnivores. Cytog and cell gen 18(5): 245-254.
-
Ledesma M, Ledesma C, Schiaffino K, Miguel A Rinas, Ricardo J Gunski (2004) Cytogenetic analysis of Panthera Onca (Felidae: Pantetherinae) from the province of Misiones, Argentina. Mastozool. Neotrop 11(1): 85-90.
-
Hatanaka T, Galetti JRPM (1999) Chromosome Banding of Chrysocyon brachyurus (Mammalia, Canidae). Cytologia 64: 159-163.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review