Molecular Evidence for Subclinical Infection of Chlamydia pecorum in Captive Koalas (Phascolarctos Cinereus) of Japanese Zoos
Chlamydial infections pose a significant threat to the health and long-term survival of koalas. However, the pathogenesis of this disease is not well understood. Chlamydia pecorum (C. pecorum) is an important pathogen affecting wild koalas’ health and their survival. However, the infection status of C. pecorum remains largely unknown in captive koalas across Japanese zoos. Therefore, in this study, we aimed to determine the infection status of C. pecorum in koalas from seven Japanese zoos. We tested swab samples obtained from koalas of these zoos for C. pecorum using qPCR and conventional PCR. We found a 21.74% overall prevalence of C. pecorum in these koalas. Female koalas showed a higher prevalence (24.24%) than male koalas (15.38%). However, no direct association was observed between koala health and C. pecorum loads. Notably, among C. pecorum positive koalas, some showed higher chlamydial loads without any classic signs associated with the progression of chlamydial diseases, such as ocular inflammation and wet bottom. We found higher C. pecorum load in adult koala than other groups. These results shed light on the subclinical infection of C. pecorum in captive koalas, and also may act as a guide for future control measures.
Introduction
The koala (Phascolarctos cinereus) faces many threats to its survival as a species, and is experiencing population declines ranging from severe to catastrophic [1]. This iconic marsupial species has been placed on the International Union for Conservation of Nature (IUCN)’s Red List of Threatened Species [2]. Anthropogenic factors—such as habitat loss due to urbanization—and climate change are clearly major contributors to the decline in koala populations, but infectious disease may also play a role. Specifically, chlamydiosis has attracted much attention from conservationists as a disease with a potentially adverse impact [3, 4].
In koalas, chlamydiosis can develop following infection with either of two chlamydiae species, Chlamydia pecorum (C. pecorum) or Chlamydia pneumoniae (C. pneumoniae) [5]. Of these species, C. pecorum is regarded as the more pathogenic and to be the causative agent for more severe disease [6]. C. pecorum infection in koalas may cause conjunctivitis, leading to blindness [7, 8]. Other manifestations include urogenital changes such as urethritis, ureteritis, nephritis, cystitis, prostatitis, orchitis and epididymitis [8, 9]. Moreover, C. pecorum infection has been reported to cause infertility in female koalas, resulting females lacking pouch youngs [10, 11].
Despite the reports of asymptomatic chlamydial infection or limited overt diseases in chlamydial infection [12, 13], the influence of chlamydial infection on long-term koala survival cannot be overlooked [12, 13, 14]. Certain C. pecorum genotypes have been associated with disease progression, while others have been associated with the absence of clinical disease [15]. The prevalence of C. pecorum infection in free range koala populations varies from region to region. Reported prevalence in populations and subpopulations across Australia include figures of 4% to 71% [4], 0%-85% [6, 16, 17], and up to 88% [18, 19, 20]. Some island populations are regarded as chlamydia-free, but the disease is generally regarded as endemic in mainland Australia, although there are differences in the manifestation of symptoms among and between populations with a high prevalence of C. pecorum [20].
In addition to the wild populations in Australia, zoos around the world are home to captive koala populations. These captive populations represent an invaluable source of genetic diversity and a bulwark against the threat to this species, as well as providing many opportunities for research on koala chlamydiosis and strategies to deal with it. Furthermore, the challenge of maintaining sustainable captive populations requires the diagnosis and treatment of the disease in zoo-reared koalas [21, 22]. However, chlamydial disease in captive koalas remains less studied. For a better understanding of the influence of any disease on its host, the prevalence study is critical. Moreover, knowledge of prevalence and clinical severity could inform strategies to combat chlamydiosis. Therefore, it is important to know the prevalence of chlamydial infection status such as C. pecorum infection in the koalas both in the wild and captivity.
The koala population in Japanese zoos represents a promising target for research in this field. This population is maintained across 7 zoos in the country [23], with social housing at varying degrees of population density. These koalas receive regular medical examinations, and the zoos are affiliated with laboratories capable of PCR testing for the pathogens that cause chlamydial disease. In this study, we determined the prevalence of C. pecorum infection in koalas in Japanese zoos, through qPCR and conventional PCR analyses of ocular swab samples. Furthermore, we investigated any associations between C. pecorum infection and demographic factors such as age and sex and health parameters, including body condition and presence or absence of abnormal clinical signs.
Materials and Methods
Ethics approval and Consent to Participate
This study was performed in accordance with the protocols of the Institutional Animal Care and Use Committee of the Joint Faculty of Veterinary Medicine, Kagoshima University, Japan for scientific purposes (Approval Number: 19K001).
Sample Collection and Processing
We targeted the captive koala population in Japan for analysis. This population is housed in seven zoos across the country: Kanazawa Zoo, Tama Zoological Park, Kobe Oji Zoo, Awaji Farm England Hill Zoo, Hirakawa Zoological Park, Saitama Children’s Zoo and Nagoya Higashiyama Zoo. All koalas’ origin is Australia and most of them are from Queensland, except Awaji Farm England Hill Zoo in which koalas are originally from Victoria, Australia.
At each zoo, koalas were housed in environments maintained within a constant temperature range (23~25°C) using an air conditioning system. Each habitat is designed, as far as possible, to simulate the natural habitat in order to avoid any general stress, and provided the koalas with an ad libitum supply of eucalyptus leaves.
Between June and October 2021, ocular swab samples (n=46) were collected from the conjunctivas of both eyes of each koala by zoo veterinarians in accordance with the relevant institution’s standard operating procedure. The samples were then shipped to Transboundary Animal Diseases (TAD) Center (Kagoshima University, Japan) on ice, and stored at -30°C until analysis.
Demographic and Clinical Assessments
The sex and age of koalas were recorded from the zoo’s data at the time of sampling. The koalas were subdivided into five age groups such as joeys (age <1 year), juvenile (age = 1-2 years), young adult (age = 2-4 years), adult (age = 4-8 years), old (age>8 years) koalas [24]. At the same time, each koala was examined by a zoo veterinarian to determine body condition score on a scale of 1 to 5. This score was determined by palpating muscle in the scapular region. The koala’s condition was regarded as ‘poor’ based on a score ≤2, ‘good’ based on a score of 3, and ‘very good” based on a score 5 [24]. Each koala was also observed for any visible clinical signs and subjected to blood sampling for determination of white blood cells (WBCs) count. Based on the results for body condition score, observed clinical signs, and WBC data, the zoo veterinarian made a comment on overall health status of individual koala.
Extraction of DNA
DNA was extracted from each ocular swab sample (400µL) using a QIAmp DNA Mini Kit, in accordance with the manufacturer’s instructions (QIAGEN), and eluted in 100 μl of the AE Elution buffer provided with the kit. The DNA concentration and purity of the extracted DNA were measured using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc., Waltham, MA USA), and DNA samples were kept at 30 °C until analysis.
Determination of C. pecorum by Real Time PCR
Quantitative real-time PCR (qPCR) was performed to test for the presence of C. pecorum and determine the chlamydial load in each koala, using extracted DNA as a template. The forward primer was 5’ ATCGGGACCTTCTCATCGAG 3’ and the reverse primer was 5’ GACTAACAGTATAAGCAGTG 3’, and the primers targeted a 108 bp fragment of the K7 gene for hypothetical protein gene partial coding sequence (LC681833.1) or partial genome sequence (CP088917.1, CP085634.1, CP080403.1 etc.) of C. pecorum. The qPCR assays were carried out with a total volume of 20µL, consisting of 10 µL Brilliant-III Ultra-Fast SYBR Green Q-PCR Master Mix (Agilent Technologies, Santa Clara, CA, USA), 0.2 µL of 10 pM forward and reverse primers and 5 µL DNA sample as a template, following the manufacturer’s instructions using a CFX Connect Real-Time PCR Detection System (Bio-Rad, USA). PCR conditions were initial denaturation at 95°C for 3 min, and 40 cycles of 95°C for 5 s and primers annealing at 50°C for 10 s. The specificity of the PCR was confirmed by melt curve analysis. Samples were run in duplicate, and a positive and negative control, as well as a standard curve for the C. pecorum gene was included in all qPCR assays. All samples that showed ≥ 50 copies/μL DNA were considered as positive [17].
Determination of C. pecorum By Conventional PCR
A Phusion PCR was performed on a sub-set (10/46) of C. pecorum positive koalas for the validation of qPCR positive samples. C. pecorum positive DNA, as determined by qPCR was used as the template for conventional PCR amplification. The primers and their concentration used were the same as of qPCR (mentioned above). The cyclic conditions for the PCR were as follows: initial denaturation at 98°C for 2 min, denaturation at 98°C for 30 s, annealing at 50°C for 30 s, extension at 72°C for 1 min, followed by 40 cycles, and final extension at 72°C for 5 min. Next, the resulting PCR fragments were subcloned into the Zero Blunt TOPO vector and sequenced. The sequence data of C. pecorum were submitted to DDBJ (accession no. LC681833- LC681840).
Statistical Analysis
All statistical analyses were performed using the STATA software package, and p-values of ≤0.05 were regarded as significant. The Kruskal-Wallis test was performed to determine associations between C. pecorum load and risk factors based on demographic or health characteristics, age and body condition (BC). A Mann-Whitney test was also performed comparing between sex.
Results
Population, Clinical Signs and Health Status of Koala
Across the seven Japanese zoos, we obtained a total of 46 captive koalas for evaluation in this study. The age, sex, and body condition score of the study population are summarized in Table 1. This population comprised of 13 male (age range: 11 months to 12 years) and 33 female (age range: 10 months to 24 years) koalas. Results of clinical signs and WBC counts are shown in Table 2. Five of the 46 koalas showed some symptoms at sampling; specifically, loose stools in two koalas, alopecia in two koalas, and abdominal bloating in one koala. WBC counts in all koalas were normal [25]. In each case, the examining veterinarian made an overall comment on koala health as ‘satisfactory’ (Table 2).
| Variables | Name of the Zoos | ||||||
|---|---|---|---|---|---|---|---|
| Kanazawa Zoo (n=4) | Tama Zoological Park (n=3) | Kobe Oji Zoo (n=8) | Awaji Farm England Hill Zoo (n=4) | Hirakawa Zoological Park (n=12) | Saitama Children’s Zoo (n=5) | Nagoya Higashiyama Zoo (n=10) | |
| Sex | |||||||
| Male | 1 (25%) | 2 (66.67%) | 2 (25%) | 2 (50%) | 2 (16.67%) | 1 (20%) | 3 (30%) |
| Female | 3 (75%) | 1 (33.33%) | 6 (75%) | 2 (50%) | 10 (83.33%) | 4 (80%) | 7 (70%) |
| Age group | |||||||
| Joey | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 2 (20%) |
| Juvenile | 1 (25%) | 0 (0%) | 0 (0%) | 0 (0%) | 5 (41.67%) | 2 (40%) | 1 (10%) |
| Young Adult | 0 (0%) | 1 (33.33%) | 4 (50%) | 0 (0%) | 2 (16.67%) | 2 (40%) | 3 (30%) |
| Adult | 3 (75%) | 2 (66.67%) | 3 (37.5%) | 0 (0%) | 4 (33.33%) | 1 (20%) | 1 (10%) |
| Old | 0 (0%) | 0 (0%) | 1 (12.5%) | 4 (100%) | 1 (8.33%) | 0 (0%) | 3 (30%) |
| BC | |||||||
| Poor | 0 (0%) | 0 (0%) | 0 (0%) | 1 (25%) | 1 (8.33%) | 0 (0%) | 0 (0%) |
| Good | 3 (75%) | 3 (100%) | 0 (0%) | 3 (75%) | 4 (33.33%) | 5 (100%) | 10 (100%) |
| Very good | 1 (25%) | 0 (0%) | 8 (100%) | 0 (0%) | 7 (58.33%) | 0 (0%) | 0 (0%) |
Table 1: Study population and characteristics (sex, age and body condition) of koalas from different zoos.
| Koala ID | Sex | Age | Group | BC | C. pecorum copies/μl DNA | WBC (10²/μl) | Clinical signs |
|---|---|---|---|---|---|---|---|
| KU_TZ_07 | Female | 3 Y | Young adult | Good | 65.46 | - | - |
| KU_HZ_19 | Female | 1 Y | Juvenile | Very good | 3.3e+07 | 62 | - |
| KU_HZ_20 | Female | 6 Y | Adult | Poor | 79.48 | 69 | Alopecia |
| KU_HZ_23 | Female | 4 Y 2 M | Adult | Very good | 2684279 | 89 | - |
| KU_HZ_25 | Female | 3 Y | Young adult | Good | 64.08 | 81 | Loose stool |
| KU_HZ_27 | Female | 1 Y | Juvenile | Very good | 120.71 | 82 | - |
| KU_HZ_28 | Female | 1 Y | Juvenile | Very good | 1547.72 | 88 | - |
| KU_NZ_37 | Male | 4 Y | Young adult | Good | 69.25 | 85 | - |
| KU_NZ_43 | Female | 1 Y | Juvenile | Good | 443.32 | 69 | - |
| KU_NZ_44 | Male | 11 M | Joey | Good | 2417.08 | 69 | - |
Table 2: A Summary of _C. pecorum_ infected koalas in this study.
Prevalence of C. pecorum
A quantitative PCR assay was performed using a C. pecorum-specific primer sets to determine infection status of captive koalas. The specificity (E=96.6%, R2=0.996, slope= 3.407) and melt curve of qPCR are shown in a supplementary figure (Fig S1). A total of 10 koalas were positive for C. pecorum across the seven zoos, with varied amount of DNA copies (Table 2). The overall prevalence of C. pecorum in this population was 21.74% (10/46). The highest prevalence of C. pecorum in koalas was in the Hirakawa Zoological Park (50%, 6/12), followed by Tama Zoological Park (33%, 1/3) and Nagoya Higashiyama Zoo (30%, 3/10), and variations between zoos were statistically insignificant (Figure S1).
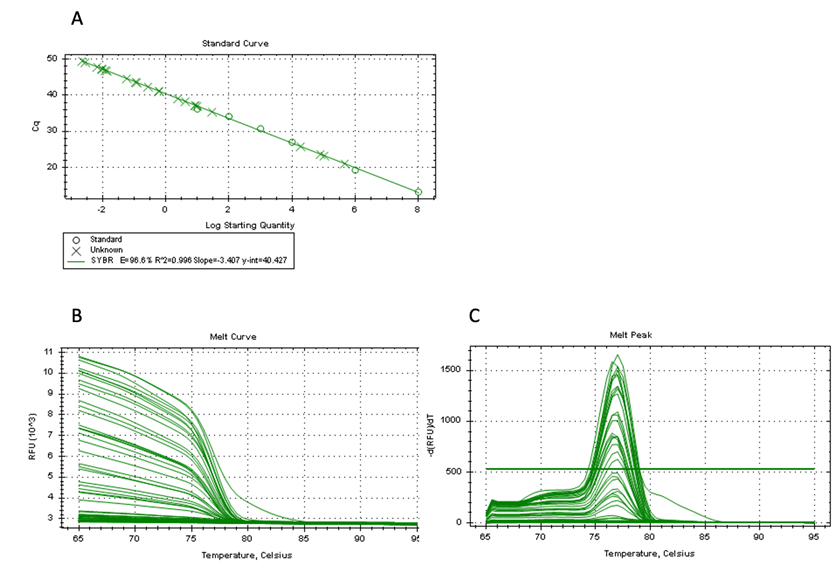
Figure-S1: The specificity of qPCR reaction. (A) A standard curve analysis, (B) A melt curve analysis, and (C) a melt peak are shown.
| Variables | Prevalence (%) of C. pecorum | P value (Fisher’s exact test) |
|---|---|---|
| Management unit | ||
| Kanazawa Zoo | 0/4 (0%) | 0.065 |
| Tama Zoological Park | 1/3 (33%) | |
| Kobe Oji Zoo | 0/8 (0%) | |
| Awaji Farm England Hill Zoo | 0/4 (0%) | |
| Hirakawa Zoological Park | 6/12 (50%) | |
| Saitama Children’s Zoo | 0/5 (0%) | |
| Nagoya Higashiyama Zoo | 3/10 (30%) | |
| Sex | ||
| Male | 2/13 (15.38%) | 0.411 |
| Female | 8/33 (24.24%) | |
| BC | ||
| Poor | 1/2 (50%) | 0.109 |
| Good | 5/28 (17.86)% | |
| Very good | 4/16 (25%) | |
| Age group | ||
| Joey | 1/2 (50%) | 0.115 |
| Juvenile | 4/9 (44.44%) | |
| Young Adult | 3/12 (25%) | |
| Adult | 2/14 (14.29%) | |
| Old | 0/9 (0%) |
Table 3: Prevalence of C. pecorum infection regarding zoo, sex, body condition, and age.
The C. pecorum load ranged between 69.25 and 2417.08 copies/μL DNA (median 1243.165) in infected male koalas, and 65.46 to 32807365.86 copies/μL DNA (median 282.015) in infected female koalas (Table 2). Female koalas also showed a higher prevalence than males (24.24%; Table 3). In addition, koalas with poor BC showed the highest prevalence of C. pecorum (50%; Table 3). By age, the highest prevalence of C. pecorum was found in joey (50%), followed by the juvenile (44.44%), adolescent (25%), and fully adult (14.29%) (Table 3).
Association between C. pecorum Loads And Population Characteristics
Mann-Whitney U test was performed for the detection of association between C. pecorum loads and population characteristics sex. The Kruskal-Wallis test also was performed to determine associations between C. pecorum load and risk factors based on age and BC. C. pecorum loads did not differ significantly with respect to sex (Figure 1A); however, the highest C. pecorum loads were observed in female koalas. Similarly, risk analysis revealed that BC and age cohorts were not significantly associated with C. pecorum load (Figures 1B-C).
However, we found higher C. pecorum load in very good BC koala followed by good and poor body condition koala. We also found higher C. pecorum load in adult koala, followed by juvenile and young adults.
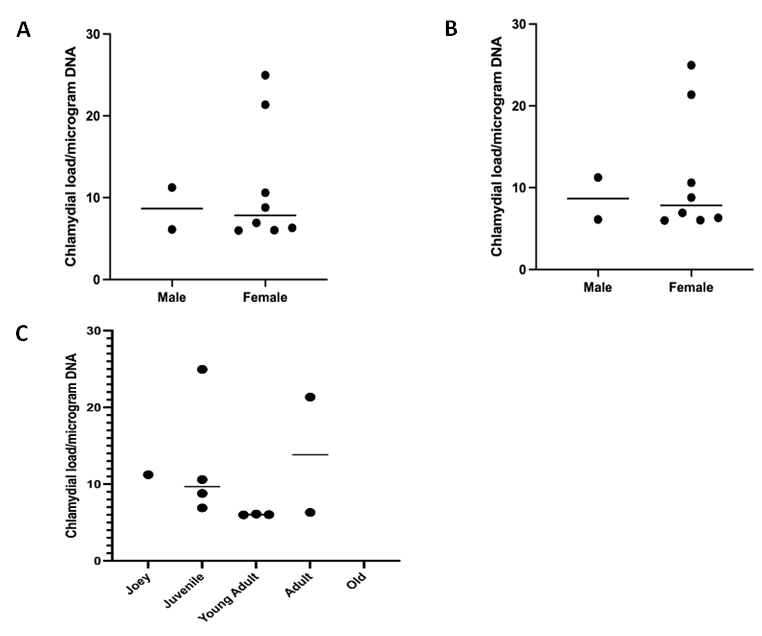
Figure 1: Associations of C. pecorum load with sex, BC, and age. C. pecorum load is indicated for sex (A) body condition, (B), and age (C). All resultant copy numbers have been log2 transformed. Each dot denotes an individual’s C. pecorum load with median line displayed. Y-axis shows individual koala values (log transformed). For Statistical significance was evaluated using the Mann-Whitney U test and Kruskal-Wallis test. Data are presented as median bar.
Validation of C. pecorum Positivity by Conventional PCR
Ten koalas were found positive for C. pecorum by qPCR. Among 10 koalas 5 showed lower copy numbers than the lowest standard (10 copies) in the qPCR. Therefore, a conventional PCR was performed for further confirmation of C. pecorum gene, resulted 8 koalas positive for C. pecorum. After sub-cloning and sequencing, we found the target size (108 bp) of the DNA fragment (Figure 2), and was 100% homogenous with NCBI reference sequences (CP088917.1, CP085634.1, CP080403.1, CP080401.1, CP004035.1, CP004034.1, CP004033.1, and CP002608.1).
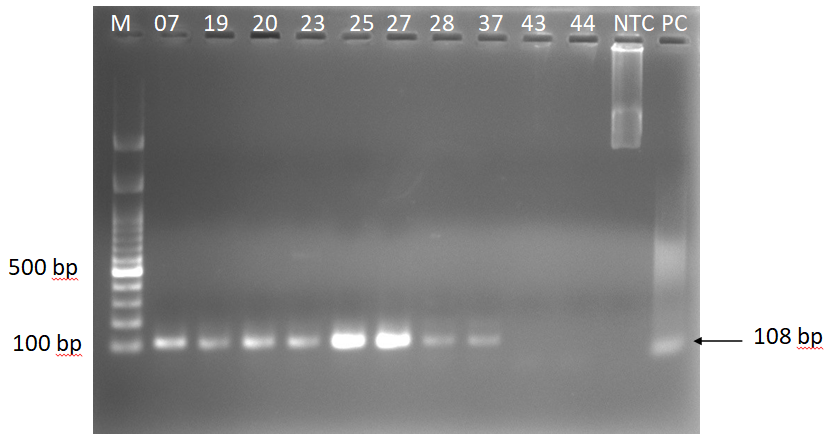
Figure 2: Amplification of C. pecorum gene using genomic DNA from the conjunctival swab samples of koalas. Lane M, 100 bp DNA marker (GeneDireX, Inc.); lane (07=KU_TZ_07, 19=KU_HZ_19, 20=KU_HZ_20, 23=KU_HZ_23, 25=KU_HZ_25, 27=KU_ HZ_27, 28=KU_HZ_28, 37=KU_NZ_37, 43=KU_NZ_43, 44=KU_NZ_44) Samples; Lane NTC, no-template control; Lane PC, positive control.
Discussion
To the authors’ knowledge, this is the first report on C. pecorum (one of the important causative agents for chlamydiosis) prevalence in the captive koala population throughout the Japan. We determined the C. pecorum prevalence based on qPCR and conventional PCR analyses of conjunctival swab samples collected from the koalas at each of seven Japanese zoos. We also determined the chlamydial load in each koala, and evaluated associations with demographic and health parameters to provide useful information on chlamydia infection and disease in captive koalas.
As a major novel finding in this study regarding prevalence, we found 21.74% of the captive koalas in Japanese zoos had subclinical infection with C. pecorum. This figure partially reflects the prevalence reported in wild koala populations [16, 26, 27], in line with the figures of 25% and 41% reported for wild koalas in New South Wales and Queensland, respectively [18], but lower than the figure of 88% for South Australia [20]. However, this prevalence was not uniform across the Japanese zoos, and ranged from 0% to 50%. Thus, further monitoring of the Japanese captive koala population may generate useful data to formulate decision on how to best conserve wild koala populations in Australia. The highest prevalence among Japanese zoos was noted at the zoo with the largest koala population (Hirakawa Zoological Park, Kagoshima), and this result could be consistent with poorer health of koala. Also, relatively high C. pecorum positive ratio was observed in female (Table 1) possibly due to the group rearing of female koalas in Japanese zoo. No significant association was observed between chlamydial load, age, sex, and zoo. Moreover, the lack of any association between chlamydial load and overt signs of disease in the study population provides some interesting evidence for the debate over the severity of chlamydiosis as a threat to wild koalas, warranting further investigation on the pathogenicity of C. pecorum in the wild koala populations [16, 26, 27].
Researcher has argued that C. pecorum of itself may not be the major driver of population decline for the species. Indeed, KoRV-B has been reported to be a significant contributing factor to the development of chlamydial disease [28]. The clinical onset of chlamydiosis may be triggered by factors such as environmental stress, such as anthropogenic habitat destruction [29]. This might explain associations between C. pecorum infection and clinical disease in the wild, but the infection would not be the causative factor [13]. In our study, the higher prevalence was observed in joey, followed by juveniles, and the low prevalence was seen in older koalas. In contrast, a previous study [30] reported the higher prevalence in juvenile koala. These age- related findings are also consistent with those reported in a previous epidemiolocal investigation in wild koalas [31]. The Japanese captive koala population is too small to allow a definitive conclusion, but the absence of overt disease among C. pecorum-positive koalas in this study may support the argument that susceptibility to chlamydial disease may be linked to environmental stress, in a key finding from this study. However, further investigations of the pathogenesis of chlamydial disease are clearly warranted.
We found no significant association between sex and chlamydial load in C. pecorum-positive koalas, as stated above. However, prevalence appeared to be markedly higher in females than males (24.24% vs. 15.38%). Further studies in larger populations are required, but these findings tend to support the possibility that female koalas may act as subclinical carriers of C. pecorum infection. Couple with the high prevalence in juvenile koalas, our study findings may suggest that chlamydia can be transmitted from dams to their off-spring through close contact and pap feeding. Sex and fighting have been suggested as routes of transmission [26, 32], but our findings suggest these routes may not have played a major role for infections in this study population.
Due to some limitations of handling of zoo koala, the study, chlamydial detection was based on ocular swab samples taken from the bilateral conjunctivas of each animal. In the majority of previous relevant studies, samples were obtained by swabbing of the urogenital tract or conjunctivas of free-ranging koalas. However, Canfield et al. reported 16% C. psittaci positivity in conjunctival swab samples obtained from captive koalas [33].
Non-endogenized KoRV subtypes have been suggested to comprise the health of koalas [34]. There may be other factors/agents that might contribute to C. pecorum infection in koalas, such as the presence of other chlamydial species, KoRV infection and/or other viral infections [30]. We observed that all koalas used in this study were infected with KoRV-A and some of them were co-infected with other KoRV subtypes [23]. The effect of KoRV subtype co-infection should be addressed in the future study. Overall, this study shows a subclinical chlamydial infection in these koala populations, and therefore it might be interesting to investigate further how/if chlamydial strain, koala genetics, stress levels and immunophenotype, co-infections by KoRV or herpesviruses, or stress responses might affect koala health.
Competing Interests
The authors declare that they have no competing financial interests.
Acknowledgments
We would like to thank Kobe Oji zoo for supplying the koala sample. This study was supported by the Ministry of Education, Culture, Sports, Science and Technology, Japan. We would like to thank Mr. Smith Henry Ivan of the Joint Faculty of Veterinary Medicine, Kagoshima University, Japan for his enthusiastic English edition of this manuscript.
References
-
Gonzalez AV, Allavena R, McKinnon A, Larkin R, Henning J (2017) Decline causes of Koalas in South East Queensland, Australia: a 17-year retrospective study of mortality and morbidity. Scientific Reports 7: 42587.
-
Woinarski J, Burbidge AA (2016) _Phascolarctos_ _cinereus_. The IUCN Red List of Threatened species. T16892A21960344.
-
Hemming V, Hoffmann M, Jarrad F, Rumpff L (2018) NSW Koala research plan: Expert elicitation of knowledge gaps. In: Research CoEaE, (Edn.), Office of Environment and Heritage. Melbourne, Victoria, Austriala: The University of Melbourne, pp: 122.
-
Quigley BL, Timms P (2020) Helping koalas battle disease - Recent advances in Chlamydia and koala retrovirus (KoRV) disease understanding and treatment in koalas. FEMS Microbiology Reviews 44(5): 583-605.
-
Glassicki T, Giffard P, Timms P (1996) Outer Membrane Protein 2 Gene Sequences Indicate that _Chlamydia_ _pecorum_ and _Chlamydia pneumoniae_ Cause Infections in Koalas. Systematic and Applied Microbiology 19(3): 457-464.
-
Polkinghorne A, Hanger J, Timms P (2013) Recent advances in understanding the biology, epidemiology and control of chlamydial infections in koalas. Veterinary Microbiology 165(3-4): 214-23.
-
Blanshard W, Bodley K (2008) In ‘Medicine of Australian Mammals’. LV and RW (Eds.), Collingwood: CSIRO.
-
Wan C, Loader J, Hanger J, Beagley K, Timms P, et al. (2011) Using quantitative polymerase chain reaction to correlate _Chlamydia pecorum_ infectious load with ocular, urinary and reproductive tract disease in the koala (_Phascolarctos cinereus_). Australian Veterinary Journal 89(10): 409-412.
-
Johnston SD, Deif HH, McKinnon A, Theilemann P, Griffith JE, et al. (2015) Orchitis and Epididymitis in Koalas (_Phascolarctos cinereus_) Infected with _Chlamydia_ _pecorum_. Veterinary Pathology 52(6): 1254-1257.
-
McColl KA, Martin RW, Gleeson LJ, Handasyde KA, Lee AK (1984) Chlamydia infection and infertility in the female koala (_Phascolarctos cinereus_). Veterinary Rehabilitation & Exercise Clinic of the Carolinas 115(25-26): 655.
-
Fabijan J, Caraguel C, Jelocnik M, Polkinghorne A, Boardman WSJ, et al. (2019) _Chlamydia pecorum_ prevalence in South Australian koala (_Phascolarctos_ _cinereus_) populations: Identification and modelling of a population free from infection. Scientific Reports 9: 6261.
-
Madden D, Whaite A, Jones E, Belov K, Timms P, et al. (2018) Koala immunology and infectious diseases: How much can the koala bear? Developmental & Comparative Immunology 82: 177-185.
-
McCallum H, Kerlin DH, Ellis W, Carrick F (2018) Assessing the significance of endemic disease in conservation-koalas, chlamydia, and koala retrovirus as a case study. Conservation Letters 11: e12425 e12425.
-
Jackson S (2003) Australian mammals: biology and captive management, pp: 524.
-
Robbins A, Hanger J, Jelocnik M, Quigley BL, Timms P (2020) Koala immunogenetics and chlamydial strain type are more directly involved in chlamydial disease progression in koalas from two south east Queensland koala populations than koala retrovirus subtypes. Scientific Reports 10(1): 15013.
-
Devereaux LN, Polkinghorne A, Meijer A, Timms P (2003) Molecular evidence for novel chlamydial infections in the koala (_Phascolarctos cinereus_). Systematic and Applied Microbiology 26(2): 245-253.
-
Nyari S, Waugh CA, Dong J, Quigley BL, Hanger J, et al. (2017) Epidemiology of chlamydial infection and disease in a free-ranging koala (_Phascolarctos cinereus_) population. PLoS One 12(12): e0190114.
-
Patterson JL, Lynch M, Anderson GA, Noormohammadi AH, Legione A, et al. (2015) The prevalence and clinical significance of Chlamydia infection in island and mainland populations of Victorian koalas (Phascolarctos cinereus). Journal of Wildlife Diseases 51(2): 309-317.
-
Legione AR, Patterson JLS, Whiteley PL, Amery-Gale J, Lynch M, et al. (2016) Identification of unusual _Chlamydia_ _pecorum_ genotypes in Victorian koalas (_Phascolarctos_ _cinereus_) and clinical variables associated with infection. Journal of Medical Microbiology 65(5): 420-428.
-
Speight KN, Polkinghorne A, Penn R, Boardman W, Timms P, et al. (2016) Prevalence and pathologic features of _Chlamydia pecorum_ infections in South Australian koalas (_Phascolarctos cinereus_). Journal of Wildlife Diseases 52(2): 301-306.
-
Phillips S, Quigley BL, Olagoke O, Booth R, Pyne M (2020) Vaccination of koalas during antibiotic treatment for Chlamydia-induced cystitis induces an improved antibody response to _Chlamydia pecorum_. Scientific Reports 10(1): 10152.
-
Chen CJ, Gillett A, Booth R, Kimble B, Govendir M (2022) Pharmacokinetic Profile of Doxycycline in Koala Plasma after Weekly Subcutaneous Injections for the Treatment of Chlamydiosis. Animals (Basel). 12(3): 250.
-
Hashem MA, Kayesh MEH, Maetani F, Goto A, Nagata N, et al. (2022) Subtype distribution and expression of the koala retrovirus in the Japanese zoo koala population. Infection, Genetics and Evolution 102: 105297.
-
Sarker N, Fabijan J, Owen H, Seddon J, Simmons G, et al. (2020) Koala retrovirus viral load and disease burden in distinct northern and southern koala populations. Scientific Reports 10: 263.
-
Canfield PM, O’Neill ME, Smith EF (1989) Haematological and biochemical reference values for the koala (_Phascolarctos cinereus_). Australian Veterinary Journal 66(10): 324-326.
-
Jackson M, White N, Giffard P, Timms P (1999) Epizootiology of Chlamydia infections in two free-range koala populations. Veterinary Microbiology 65(4): 255- 264.
-
Mackie JT, Gillett AK, Palmieri C, Feng T, Higgins DP (2016) Pneumonia due to _Chlamydia pecorum_ in a Koala (_Phascolarctos cinereus_). Journal of Comparative Pathology 155(4): 356-360.
-
Waugh CA, Hanger J, Loader J, King A, Hobbs M, et al. (2017) Infection with koala retrovirus subgroup B (KoRV-B), but not KoRV-A, is associated with chlamydial disease in free-ranging koalas (_Phascolarctos_ _cinereus_). Scientific Reports 7: 134.
-
Melzer A, Carrick F, Menkhorst P, Lunney D, St John B (2000) Overview, critical assessment, and conservation implications of Koala distribution and abundance. Conservation biology 14(3): 619- 628.
-
Russell I, Timms P, Hanger J, Loader J, Gillett A, et al. (2018) Prevalence of _Chlamydia pecorum_ in Juvenile Koalas (_Phascolarctos cinereus_) and Evidence for Protection from Infection via Maternal Immunization. Journal of Wildlife Diseases 54(4): 863-865.
-
Hulse LS, Beagley K, Ellis W, Fitzgibbon S, Gillett A, et al. (2020) Epidemiology of chlamydia-induced reproductive disease in male koalas (_Phascolarctos cinereus_) from southeast Queensland, Australia as assessed from penile urethral swabs and semen. Journal of Wildlife Diseases 56(1): 82-92.
-
Whittington R (2001) Chlamydiosis in koalas. In: Infectious diseases of wild mammals, Williams ES, Barker IK (Eds.). Iowa State University Press, pp: 423- 434.
-
Canfield PJ, Love DN, Mearns G, Farram E (1991) Chlamydial infection in a colony of captive koalas. Australian Veterinary Journal 68(5): 167-169.
-
Quigley BL, Phillips S, Olagoke O, Robbins A, Hanger J, et al. (2019) Changes in Endogenous and Exogenous Koala Retrovirus Subtype Expression over Time Reflect Koala Health Outcomes. Journal of Virology 93(18): e00849-e00819.
- The Digital Stethoscope: Harnessing AI in Veterinary Medicine Without Losing Our Healing Touch
- Meningoencephalomyelitis of Unknown Etiology: Short-Term Effect of Two Treatment Protocols on Cerebrospinal Fluid
- Safety and Efficacy of the HomeoPet Cough in Domestic Pets –A Clinical and Correction Analysis Based Upon User Response Survey
- Non Human Animals Responses to Social Loss
- Owner Reported Clinical Outcomes of a Homeopathic Proprietary Preparation for the Treatment of Upper Respiratory and Nasal Disorders in Companion Animals
- Effects and Diagnostic Approach of Ultrasound in Veterinary Practice: A Systematic Review