On the Correspondence between Stable Carbon Isotopes in Planktic Foraminiferal Species Globigerina bulloides and the Nutrients along a North-South Transect in South-Western Indian Ocean
During the pilot expedition to the Southern Indian Ocean onboard ORV Sagar Kanya a total of twenty one surface sediment samples were collected along a north-south transect between 9.50 N to 55.010 S latitude and 73.560 E and 44.860 E longitudes. These samples were collected to study the response of carbon isotope (δ13C values) fractionation in calcareous shells of planktic foraminiferal species Globigerina bulloides towards the nutrient contents of the ambient sea waters along a northsouth transect in the south-western Indian Ocean. It may be inferred that isotopic values (δ13C) in general increase as the nutrient content of ambient water masses decreases in the regions south of 300S latitude. On the other hand δ13C values are relatively low in the region north of 30oS having higher nutrients in the ambient waters. The results of present study demonstrate the potential of foraminiferal δ13C values to trace the signature of paleo-nutrients changes in this region. The inferences drawn here further necessitates to take up similar studies along many transect in and around study area to arrive at a comprehensive picture.
Introduction
Nutrients are necessary for the growth and survival of animals, plants and other organisms. However, industrial, agricultural and urban development has dramatically increased nutrient levels in river systems, including nitrogen and phosphorus containing substances, degrading water quality, causing acidification and eutrophication and affecting aquatic ecosystems. Nutrient assessment and management in river systems has been an important part of water resource management for the past few decades, but the provision of appropriate and effective nutrient assessment and management continues to be a challenge for water resource managers and policy makers. Difficulties in assessment and management are due in part to the fact that nutrients in rivers may originate from a variety of sources, take numerous pathways and transform into other substances.
The isotopic compositions are a valuable tool to follow and trace the source and cycling of organic matter in the marine systems. The stable isotope composition of marine organisms is a fundamental property that reflects their physiology and status and are good tracers of their contribution in both supplying carbon to the marine food web and depositing carbon to the sediment. These Stable isotopes have also emerged as the powerful tools because they offer record of past environmental conditions such as temperature, salinity, nutrients and pH as well as humidity, biodiversity, vertical migration of foraminifera during their life cycle, changes in symbiont photosynthetic and respiratory rates [1, 2, 3, 4, 5, 6, 7]. The distribution of d13C present in the ocean is closely related to the circulation pattern and the oxygen and nutrient contents of water masses [8]. Carbon isotopes are also a useful and underused indicator of net ecosystem production as elevated carbon isotope (δ13C) values are linked with areas and periods of high productivity. Accordingly carbon isotopic ratios of planktic foraminifera are being used to provide information about the changes in the oceanic nutrient concentration, ventilation and circulation pattern of the ocean waters [9, 10, 11, 12, 13]. The d13C of foraminifera are used to infer the influence of different water masses, exchange of CO2 between sea water and atmosphere and biological productivity and nutrient cycling waters [14, 15] and to understand the processes responsible for the variability in atmospheric pCO2 from glacial to interglacial times [4, 16, 17, 18].
Globigerina bulloides can be found in a wide range of thermal environments from subpolar to tropical ocean waters, although it is generally more abundant in cool subtropical/transitional waters [19]. In the study area G. bulloides dominates planktic foraminiferal assemblage. Its abundance increases considerably during periods of high phytoplankton productivity which may result from spring bloom conditions or upwelling [20]. Previous studies used the d18O data of Globigerina bulloides for inferring ambient environmental conditions both in space and time [20, 21, 22, 23]. Although stable isotope data abounds in the literature, relatively limited data on d18C values of Globigerina bulloides is available for the Indian sector of the Southern Ocean.
In this context, we present here a study of variation of the d13C values in the calcareous shells of planktic foraminiferal species Globigerina bulloides in surface sediment samples collected over the Southern Indian Ocean along a North- South transect. Our analysis throws new lights on the observed d13C and environmental changes associated with different nutrient regimes.
The Study Area
The Antarctic Circumpolar Current (ACC) which provides pathways between the major ocean basins, plays a vital role in the global redistribution of salt, nutrient, heat, etc [24, 25] and provides the nutrient input to the Southern Ocean. According to Wyrtki [26], the ACC extends from the sea surface to depths of 2000-4000 m and can be spatially spread to the extent of 1000-2000 km. The water column circulation or mixing is also due to the rise of relatively warmer deep ocean waters to the colder surface of the ocean just south of the current which compensates for the sinking of surface water along the edge of icy Antarctica and further north. Ultimately, carbon sinks into the deep ocean as particulate organic matter. As the particulate matter decays, the carbon in the organic matter gets oxidised into carbon dioxide. Because of this deep ocean “biological pump” of carbon, the atmospheric carbon dioxide is lower than it otherwise would be [27].
The study area is an integral part of the South Indian Ocean where circulation is characterized by a subtropical anticyclonic gyre [26]. The poleward Agulhas Current lies to its west, the eastward flowing Antarctic Circumpolar Current (ACC) on its south and equator ward flowing West Australian Current on its east. The Subtropical Front (STF) is located at approximately 42-43˚S latitude in central south Indian Ocean.
Southern Ocean, the study area is an important part of the global climate system and can be divided into three zones based on the dominant dynamics: The Western Boundary Current (WBC) zone (35°-45° S), the Antarctic Circumpolar Current (ACC) zone (45°-60° S) and the Seasonal Sea Ice (SSI) zone (60°-75° S). The WBC zone contains a number of energetic western boundary currents, such as the Agulhas Current, the Brazil/Malvinas Current and the East Australia Current. Hydrographic conditions in Southern Ocean (SO) are modulated by an eastward flowing Antarctic Circumpolar Current (ACC) which is embedded with numerous circumpolar fronts [28, 29].
The barotropic Antarctic Circumpolar Current (ACC) reaches the ocean floor and is able to mix efficiently the North Atlantic Deep Water (NADW) and deep waters from Indian and Pacific Oceans. The mixture of these deep waters, the Circumpolar Deep Waters (CDW), then spreads back in to other oceans basins.
It is well known that Antarctica Bottom Water and Circumpolar Deep Water (CDW) enter the Indian Ocean in the west off Madagascar and East Africa, and in the east along the Ninety East Ridge [30, 31]. Furthermore, deep upwelling across the area north of 18° S with an intensity of about 4 x 10-7 m/s, about three times larger than estimated for the Pacific and Atlantic Oceans has been noticed earlier [32]. A very strong, deep, meridional overturning cell, consisting of an inflow of 27±10 Sv below about 1800 m near 32º S and outflow above that depth was augmented by an Indonesian Through flow of 6.6 Sv has also been noticed [33]. The overturning circulation carries layers of warm near-surface water and cold deep water in alternate directions thereby, transporting heat along with allied properties. The Southern Ocean plays a unique role in the global scale overturning circulation as well due to the circumpolar connection in the Southern Ocean. Water found at intermediate and abyssal depths at low latitudes rises towards the surface in the Southern Ocean. Deep water that upwells closer to Antarctica is cooled by the cold air blowing off the continent and its salinity is increased by brine released during sea ice formation. The dense water produced in this way sinks near the continental margin of Antarctica and returns to the north in deep currents flowing along the sea floor.
Various frontal regions such as, STF in between 42-43°S (water temperature ~13°C), SAF (seawater T in the range of 7-9°C) identified between 44-45°S and (PF) between 52-56°S (seawater T near 5°C) have been demarcated on the basis of these physical properties of the water column. Sea surface temperature variation from 21.8°C to 19°C was reported between 35° to 40°S (salinity 34.2 to 35.5‰) [34]. The mixed layer depth was reported at 50-75 m in this region. The vertical variation of temperature indicates the possibility of thermocline depth upto 100 m. The temperature minimum was noticed around 100-200 m after the 49°S latitude [27]. The different sampling stations represent distinct latitudinal belt and can be grouped into several zoogeographic provinces which are primarily influenced by ecology and climate, namely tropical, subtropical, transitional and sub Antarctic [35, 36, 37, 38, 39].
Materials and Methods
A total of 21 recent sea surficial sediment samples (comprising Peterson grab, gravity and piston core-top samples), collected during the 199C and 200th (also known as Pilot expedition to Southern Ocean) cruises of ORV Sagar Kanya, and were used for the present study. Geographically this study area covers transect between 9.50 N to 55.010 S latitude and 73.560 E and 44.860 E longitude in the Indian Ocean Sector of the Southern Ocean (South western Indian Ocean). The locations of various stations are shown in Figure 1. The calcium carbonate compensation depth and the lysocline in and around the study area lie below 4,400-4,700 meters water depth [40]. All samples were collected well above this water depth (Table 1) to avoid any dissolution effects on the tests of planktic foraminifers. Table 1 showing details of sampling stations.
| Sample No. | Location Latitude | Longitude | Water Depth (M) | Sampling Method | |
|---|---|---|---|---|---|
| 1. | SK199C/03 | 09° 30.00'N | 73° 30.60'E | 1030 | Peterson Grab |
| 2. | SK199C/06 | 08° 08.00'N | 73° 33.86'E | 2250 | Spade Corer |
| 3. | SK199C/10 | 01° 55.38'S | 67° 52.85'E | 2597 | Spade Corer |
| 4. | SK199C/12 | 04° 41.18'S | 67° 05.75'E | 3320 | Spade Corer |
| 5. | SK199C/13 | 07° 21.89'S | 67° 10.37'E | 3305 | Spade Corer |
| 6. | SK199C/14 | 09° 10.74'S | 65° 57.33'E | 3373 | Spade Corer |
| 7. | SK199C/15 | 11° 25.46'S | 67° 24.16'E | 3513 | Gravity Corer |
| 8. | SK199C/16 | 12° 35.56'S | 67° 08.59'E | 3722 | Gravity Corer |
| 9. | SK199C/17 | 15° 16.71'S | 66° 00.77'E | 3368 | Piston Corer |
| 10. | SK199C/19 | 16° 16.06'S | 63° 27.86'E | 4003 | Piston Corer |
| 11. | SK200/05 | 28° 19.29'S | 48° 43.56'E | 2295 | Piston Corer |
| 12. | SK200/09 | 30° 54.85'S | 44° 51.37'E | 2227 | Gravity Corer |
| 13. | SK200/14 | 36° 07.30'S | 44° 53.54'E | 2805 | Piston Corer |
| 14. | SK200/15 | 37° 00.00'S | 44° 59.00'E | 2984 | Peterson Grab |
| 15. | SK200/17 | 39° 01.71'S | 44° 58.17'E | 4022 | Piston Corer |
| 16. | SK200/19 | 40° 58.88'S | 45° 03.53'E | 2532 | Piston Corer |
| 17. | SK200/21 | 43° 09.00'S | 44° 59.00'E | 3210 | Peterson Grab |
| 18. | SK200/22A | 43° 41.47'S | 45° 04.22'E | 2723 | Piston Corer |
| 19. | SK200/23 | 44° 59.82'S | 45° 00.83'E | 1423 | Piston Corer |
| 20. | SK200/27 | 49° 00.34'S | 45° 13.11'E | 4377 | Gravity Corer |
| 21. | SK200/33 | 55° 00.39'S | 45° 00.63'E | 4185 | Piston Corer |
Table 1: Details of sampling stations, depth and sampling methods.
![Figure 1: The calcium carbonate compensation depth and the lysocline in and around the study area lie below 4,400-4,700 meters water depth [40]. All samples were collected well above this water depth (Table 1) to avoid any dissolution effects on the tests of planktic foraminifers. Table 1 showing details of sampling stations.](/fulltextimages/5839/fig_1.jpeg)
Immediately after recovery, the entire sediment sample (top 1 cm of the sediment core/grab) were stained with Rose Bengal and preserved in 10% formalin to differentiate living specimen of benthic foraminifera. In the absence of exact age date for the sediment samples, the stained benthic foraminifers collected at various stations are considered to reflect modern ambient conditions at the seabed. Accordingly the hydrographic characteristics of the uppermost 200 meters of the water column would be tied to the planktic foraminiferal record of the seabed sediments.
About 5-10 g of sediments from each sample was first dried overnight at 45-50°C. The dried samples were then soaked with water, which was decanted several times, and lastly filled with distilled water. 10 ml of 10% sodium hexa-metaphosphate [Na (PO3)6] was added to each of the samples to remove the clay materials. After that, the samples were processed over 63µm sieve with distilled water. The >63µm material was dried at 45-50°C. The > 63 µm fraction was dry sieved to get >150 µm fraction. From the >150 µm fraction an aliquot was taken by coning and quartering. The representative samples were weighed and hence used for picking of minimum 10-12 specimen of the planktic indicator species Globigerina bulloides from each sample to measure the carbon isotopic composition (d13C) at the Alfred Wegner Institute for Polar and Marine Research, Germany, with a Finnigan MAT 251 Isotope Ratio Gas Mass Spectrometer coupled to an automatic carbonate preparation device (Kiel I) and calibrated via NBS 19 to the PDB scale. The values are given in d-notation versus VPDB (Vienna Pee Dee Belemnite). Precision of carbon isotope measurements based on repeated analyses of a laboratory standard over a one-year-period was better than 0.06% for carbon. To compare the present dataset with modern hydrography we used the average phosphate and nitrate data in the uppermost 200 m of the water column, extracted for selected positions along the transect from the NOAA World Ocean Atlas [41].
Results and Discussion
In the study area, the (d13C) values in calcareous shells of planktic species Globigerina bulloides has varied from -1.97‰ (at station SK200/09) to 0.673‰ (at station SK200/27). While, the phosphate and nitrate values lie within the range 1.26 µmol at station SK 200/21 and 7.66 µmol at station SK 200/19, and between 1.38 µmol at stations SK 200/5 and 8.49 µmol at station SK 199C/12 (Figure 2a-c). It is interesting to note that nitrate ranges from 1.38 to 8.49 µmol at 9°N to 40°S, and south of 40°S from 1.97 to 5.32 µmol. Phosphate concentration varies from 2.23 to 3.67 µmol at 10°N to 40°S, and thereafter from 1.26 to 7.66 µmol.
The general profile from lower latitudes (9.69oN) to higher latitudes (55.01oS) along the transect revealed that carbon isotopic (d13C) values has shown a relatively decreasing trend from northern latitudes to upto 30oS latitudes further south of 30oS the values started increasing (Figure 2a). Contrary to it, the profiles of phosphates and nitrates have shown relatively lower values south of 30oS latitudes region with exception at station SK200/19 which shows spikes in the Phosphate values (Figure 2b-c).
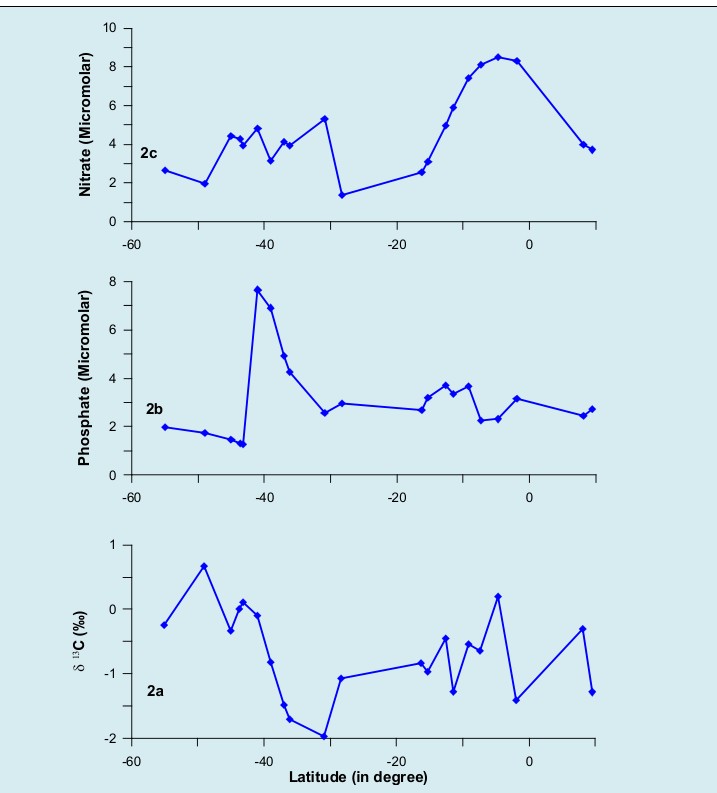
The Southern Ocean experiences unique physical processes that influence production particularly in this region. The nature and process associated with the sea ice around Antarctica have a dominant influence on water column structure, nutrient supply and primary production. Carbon isotopic values (d13C) are linked with the productivity / nutrient contents of the water masses in which they grow. It has been noticed that the nutrient content and d13C of the Dissolved Inorganic Carbon (DIC) in sea water are negatively correlated owing to the fact that in surface water masses during photosynthetic carbon fixation nutrients and preferentially the light carbon isotope are utilized. Consequently, nutrients get depleted and d13C values of DIC enriches. As deep water masses mix and gain re-mineralised carbon from surface productivity during their oceanic transit, the d13C DIC value decreases as does the d13C of foraminiferal calcite secreted from these waters [42].
The results of the present study are in agreement with the above possible explanation in southern high latitude regions, showing relatively higher values of d13C in relatively nutrient depleted waters of south of 30o S latitudes (Figure 2a-c). It seems that there is a negative correlation between d13C values and phosphate content in the region north of 30°S latitude and also in the region south of 30°S latitude (Figures 3a-b). On the contrary, though the d13C values and nitrate content are found to be negatively correlated in the region south of 30°S latitude (Figure 3d), it does not necessarily show inverse relationship between d13C values and nitrate content north of 30°S latitude (Figure 3c). By contrast, nitrate concentration is higher in the region north of 30°S latitude with an increase value of d13C. Southwards from 30°S latitude, the d13C values decreases with increasing nitrate values. Does this imply that, compared to phosphate, nitrate concentration plays a less significant correspondence with $\delta^{13}$C values in the latitudinal corridor of 9°N to 30°S? Nutrient availability plays an important but indirect role in the variation of $\delta^{13}$C values in the calcareous shells of Globigerina bulloides. Such an interesting correspondence between the $\delta^{13}$C values and nutrients looks interesting and can be explained.
The nutrient availability (nitrate and phosphate) would control primary productivity as the distribution of dissolved oxygen, nutrients and carbon in the ocean is strongly affected by production of biomass in the euphotic zone, vertical particle fluxes and demineralization of particles during sinking and post depositional processes at the ocean floor. The major nutrients (nitrate and phosphate) required for phytoplankton growth are abundant in the surface waters of the sub-Arctic, Pacific, equatorial Pacific and Southern oceans. Our inferences are in agreement with the observations of the earlier workers who notices that generally foraminiferal $\delta^{13}$C values decrease with increasing nutrient contents. The present results further support the views that the stable isotopic composition of Dissolved Inorganic Carbon (D1C) in sea water is inversely related to nutrient distribution. As stated earlier the photosynthetic fractionation in the euphotic zone and oxidation of sinking organic matter in deeper waters lead to high $\delta^{13}$C values in nutrient poor waters and low $\delta^{13}$C values in nutrient rich waters [43].
The distinct pattern of the carbon isotopic values ($\delta^{13}$C) of Globigerina bulloides along north-south transect also reflects extraneous influences on carbon isotopes fractionation as the isotope equilibration of surface seawaters with atmospheric CO2 raises $\delta^{13}$C values in cold surface waters and lowers $\delta^{13}$C values in warm surface waters [44, 45, 46, 47].
In view of the above, it may be summarized that the $\delta^{13}$C values of the calcareous shells of planktic foraminiferal species Globigerina bulloides reflect the ambient sea water $\delta^{13}$C composition and hence nutrient content of the ambient water masses. Our views are congruous with the earlier opinion that the carbon isotopic composition in Globigerina bulloides shows high correlation with the surface water phosphate values in surface sediments of N-E Atlantic and therefore may serve as a proxy for palaeo-nutrient and/or palaeo-productivity [48]. The results obtained from Southern Ocean sediment traps experiments also indicated that $\delta^{13}$C values of Globigerina bulloides is closely correlated to seasonal changes in nutrients indicating the utility of Globigerina bulloides $\delta^{13}$C values as nutrient tracer in Southern Ocean environment [49, 50].
However, the magnitude of the associated other factors such as, water chemistry and biological fractionation influencing $\delta^{13}$C values of planktic foraminiferal shells make it difficult to assess absolute paleo-nutrient content.
Conclusions and Implications
Based on the present study, it may be inferred that though the results are preliminary in nature, yet the isotopic values ($\delta^{13}$C) in general increase as the nutrient content of ambient water masses decreases in the regions south of 30°S latitude. On the other hand in the latitudinal corridor of 9°N to 30°S, though phosphate content continue to exhibit a negative correlation, to some extent implying that nitrate content plays relatively less significant role in responding to $\delta^{13}$C values at least in the above mentioned latitudinal corridor. Though the possibility of other factors influencing isotopic values (d13C) in planktic foraminiferal shell may not be ruled out completely, the nutrient content of the sea- water appears to correspond with isotopic values (d13C) of planktic foraminiferal species G. bulloides. Such an impact on stable isotopic composition of calcareous shells of planktic foraminifera looks significant from the view point of the recent phenomena of increasing acidifications of ocean at large. Impacts of such acidification on the productivity and or nutrient content of sea-water will likely to be more severe in higher latitude oceans owing to increased absorption of CO2 and the downward mixing of colder water in these regions into the deep ocean carrying carbon-di-oxide to deep waters. The results of present study underline the need for the development of quantitative determination of stable isotopic variations in response to the nutrient changes in modern marine environment prior to their application in assessing paleo-nutrients. Further, many such transects from different marine regimes of the world ocean need to be considered for such investigations to further augment the present inferences.
Acknowledgement
The authors are thankful to Dr. M.N. Rajeevan, Secretary, Ministry of Earth Sciences, Government of India. Master, officers and crew of ORV Sagar Kanya are acknowledged for providing logistics support during the collection of the samples.
References
-
Fairbanks RG, Wiebe PH, Be A WH (1980) Vertical distribution and isotopic composition of living planktonic foraminifera in the Western North Atlantic. Science 207(4426): 61-63.
-
Hemleben C, Spindler M, Anderson OR (1989) Modern Planktonic Foraminifera. Springer New York, pp: 363.
-
Spero HJ, Lerche I, Williams DF (1991) Opening the carbon isotope “vital effect’ black box, 2, quantitative model for interpreting foraminiferal carbon isotope data. Paleoceanography 6(6): 639-655.
-
Sautter LR, Thunell RC (1991) Seasonal variability in the δO and δC of planktonic foraminifera from an upwelling environment Sediment trap results from the San Pedro Basin southern California Bight. Paleoceanography 6(3): 307-334.
-
Kohfeld KE, Fairbanks RG, Smith SL, Walsh ID (1996) Neogloboquadrina pachyderma (sinistral coiling) as paleoceanographic tracers in polar oceans Evidence from Northeast Water Polynya plankton tows sediment traps and surface sediments. Paleoceanography 11(6): 679-699.
-
Bauch D, Carstens J, Wefer G (1997) Oxygen isotope composition of living Neogloboquadrina pachyderma (sin ) in the Arctic Ocean. Earth and Planetary Science Letters 146(1-2): 47-58.
-
Simstich J (1999) Die ozeanische Deckschicht des Europäischen Nordmeers im Abbild stabiler Isotope von Kalkgehäusen unterschiedlicher Planktonforaminiferenarten. Berichte Institut für Geowissenschaften University Kiel, Kiel, Germany.
-
Kroopnick PM (1985) The distribution of C of SCO in the world oceans. Deep Sea Research Part A Oceanographic Research Papers 32: 57-84.
-
Shackleton NJ (1977) The oxygen isotope stratigraphic record of the Late Pleistocene. Philo Trans R Soc London Ser B280(972): 169-182.
-
Broecker WS (1982) Ocean Chemistry During Glacial Time Geochimica Et Cosmochimica Acta. 46(10): 1689- 1705.
-
Boyle EA, Keigwin LD (1982) Deep circulation of the North Atlantic over the last 200000 Years Geochemical evidence Science. Science 218(4574): 784-787.
-
Curry WB, Lohmann GP (1982) Carbon isotopic changes in benthic foraminifera from the western South Atlantic Reconstruction of glacial abyssal circulation patterns. Quaternary Research 18(2): 218-235.
-
Duplessy JC, Shackleton NJ, Fairbanks RG, Labeyrie L, Oppo D, et al. (1988) Deepwater source variations during the last climatic cycle and their impact on the global deepwater circulation. Paleoceanography 3(3): 343-360.
-
Broecker WS, Peng TH (1982) Tracers in the Sea. New York Eldigio press.
-
Naidu PD, Niitsuma N (2004) Atypical δ 13C signature in Globigerina bulloides at the ODP site 723A (Arabian Sea) Implications of environmental changes caused by upwelling. Marine Micropaleontology 53(1-2): 1-10.
-
Shackleton NJ, Pisias NG (1985) Atmospheric carbon dioxide, orbital forcing, and climate. In the carbon cycle and atmospheric CO2: natural variations Archean to present. Chapman conference papers 32: 303-317.
-
Oppo DW, Fairbanks RG (1990) Atlantic Ocean thermohaline circulation of the last 150000 years Relationship to climate and atmospheric CO. Paleoceanography 5(3): 277-288.
-
Ahmad SM, Labeyrie LD (1994) Glacial to interglacial Holocene δ13C variations in intermediate depth water masses of the north Indian Ocean. Geo Mar Lett 14: 36- 40.
-
Toledo FAL, Costa KB, Pivel MAG, Campos EJD (2008) Tracing past circulation changes in the western south Atlantic based on planktonic foraminifera. Rev Bras Paleontol 11(3): 169-178.
-
Sautter LR, Thunell RC (1991) Planktonic foraminiferal response to upwelling and seasonal hydrographic conditions: sediment trap results from San Pedro Basin Southern California Bigh.t Journal of Foraminiferal Research 21(4): 347-363.
-
Bard E, Arnold M, Maurice P, Duprat J, Moyes J (1987) Retreat velocity of the North- Atlantic polar front during the last deglaciation determined by 14C accelerator mass spectrometry. Nature 328: 791-794.
-
Bard E, Fairbanks RG, Maurice P, Duprat J, Moyes J, et al. (1989) Sea level estimates during the last deglaciation based on d18O and accelerator mass spectrometry 14C ages measured on Globigerina bulloides. Quaternary Research 31(3): 381-391.
-
Kallel N, Labeyrie LD, Juillet Laclerc A, Duplessy JC (1988) A deep hydrological front between intermediate and deep water masses in the glacial Indian Ocean. Nature 333: 651-655.
-
Peterson RG, White WB (1998) Slow oceanic teleconnections linking the Antarctic Circumpolar Wave with the tropical El Nino-Southern Oscillation. Journal of Geophysical Research C Oceans 103: 24573-24583.
-
Jacobs GA, Mitchell JL (1996) Ocean circulation variations associated with the Antarctic Circumpolar Wavel. Geophysical Research Letters 23(21): 2947-2950.
-
Wyrtki K (1971) Oceanographic Atlas of the International Indian Ocean Expedition, 531 National Science Foundation, Washington D C.
-
Rajakumar A, Alagrsamy R, Khare N, Saraswat R, Subramaniam MM (2008) Studies on the nutrient didtribution in the Southern Ocean waters along 450 E transect. Indiaqn Jour Mar Sciences 37(4): 424-429.
-
Nowlin WD, Klinck JM (1986) The physics of the Antarctic Circumpolar. Current Rev Geophys 24(3): 469-491.
-
Anilkumar N, Luis AJ, Somayajulu YK, Ramesh Babu V, Dash M K, et al. (2006) Fronts Water Masses and Heat Content Variability in the Western Indian Sector of Southern Ocean during Austral Summer 2004. Journal of Marine Systems 63(1-2): 20-34.
-
Mantyla AW, Reid JL (1995) On the origins of deep and bottom waters of the Indian Ocean. Journal of Geophysical Research 100(2): 2417-2439.
-
Warren Bruce A (1980) Indian Ocean: Deep circulation. In: McGraw Hill Yearbook of Science and Technology, McGraw-Hill, New York, pp: 227-229.
-
Warren Bruce A (1981) Transindian hydrographic section at Lat 18°S: property distributions and circulation in the South Indian Ocean. Deep-Sea Research 28(8): 759-788.
-
Toole JM, Warren BA (1993) A hydrographic section across the subtropical South Indian Ocean. Deep-Sea Research Part 40(10): 1973-2019.
-
Anilkumar N, Dash MK, Luis AJ, Ramesh Babu V, Somayajulu YK, et al. (2005) Oceanic fronts along 45°E across Antarctic circumpolar current during austral summer 2004. Current Science 88: 1669-1673.
-
Schott G (1942) Geographie des Atlantischen Ozeans Boysen Hamburg, pp: 348.
-
Sverdrup HU, Johnson MW, Fleming RH (1942) The Oceans. Prentice-Hall Englewood Cliffs, New Jersey, pp: 1087.
-
Bradshaw JS (1959) Ecology of living planktonic foraminifera in the north and equatorial Pacific Ocean Contr Cushman Found Re. 10: 25-64.
-
Fairbridge RW (1966) The enclyclopedia of oceanography Dowen Hutchinson Ross Stroudsburg.
-
Bé AWH, Tolderlund DS (1971) Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian Oceans. In: Funnel BM and Riedel WR, (Eds.), Micropaleontology of the Oceans, Cambridge University press, London, pp: 109-149.
-
Banakar VK, Parthiban G, Pattan JN, Jauhari P (1998) Chemistry of surface sediment along a north south transect across the equator in the central Indian basin: an assessment of biogenic and detrital influences on elemental burial of the sea floor. Chemical Geology 147(3-4): 217-232.
-
Levitus S, Boyer TP (1994) World Ocean Atlas. Nat Oceanic and Atmos Admin, Washington D C.
-
Mackensen A (2004) Changing Southern Ocean palaeocirculation and effects on global climate. Antarctic Science 16(4): 369-386.
-
Matsumoto K, Lynch Stieglitz J (1999) Similar glacial and Holocene deep water circulation inferred from southeast Pacific benthic foraminiferal carbon isotope composition. Paleoceanography 14(2): 149-163.
-
Charles CD, Fairbanks RG (1990) Glacial to interglacial changes in the isotopic gradients of Southern Ocean surface water. Geological history of the polar oceans Arctic versus Antarctic, pp: 519-538.
-
Broecker WS, Maier Reimer E (1992) The influence of air and sea exchange on the carbon isotope distribution in the sea. Global Biogeochemical Cycles 6(3): 315-320.
-
Lynch Stieglitz J, Stocker TF, Broecker WS, Fairbanks RG (1995) The influence of air-sea exchange on the isotopic composition of oceanic carbon observations and modeling. Global Biogeochemical Cycles 9(4): 653-665.
-
McCorkle DC, Martin PA, Lea DW, Klinkhammer GP (1995) Evidence of a dissolution effect on benthic foraminiferal shell chemistry dC Cd/Ca Ba/Ca and Sr/Ca results from the Ontong Java Plateau. Paleoceanography 10(4): 699-714.
-
Ganssen GM, Kroon D (2000) The isotope signature of planktic foraminifera from NE Atlantic surface sediments Implications for the reconstruction of past oceanic conditions. J Geol Soc London 157: 693-699.
-
King AL, Howard WR (2004) Planktonic foraminiferal δ13C records from Southern Ocean sediment traps: New estimates of the oceanic Suess effect. Global Biogeochemical Cycles 18(2): 1-16.
-
Volkmann R, Mensch M (2001) Stable isotope composition (d18O, d13C) of living planktic foraminifers in the outer Laptev Sea and the Fram Strait. Mar Micropal 42: 163-188.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh