For an Effective and Sustainable Management of Non-Tydal Lagoon Environments to Counteract the Eutrophication Effects
Coastal lagoon environmental picture with the tendency to accumulate organic detritus, structural eutrophication, risks of dystrophy, with loss of biodiversity and fish products, is briefly described. Mitigation solutions are analyzed, focusing on the removal of macroalgal masses, artificial water renewal, sediment oxidation and more, highlighting the limits and benefits of each solution. With regard to the sediment oxidation, on which more emphasis is placed here, a review of the results obtained on two Tyrrhenian lagoons is made, through field experiences conducted between 2005 and 2021. These results showed an overall mitigation of the eutrophication effects, with reduction of nutrient load and sediment labile organic matter, reduction of macroalgal development, phanerogam return and increase in zoobenthonic biodiversity. Some possible effects of sediment resuspension on contaminants are reported.
Environmental Framework of The Lagoons
The lagoons have typically shallow depths, from a few decimetres to a few meters, resulting essentially flat flooded, in which the water is poorly renewed (non-tidal lagoons and the most continental parts of microtidal lagoons) and subject to quickly warm up or to cool down. Many marine organisms do not tolerate seasonal lowering or raisings in temperature, and even less they can tolerate high variations in temperature in the space of a single day, as a result of sudden meteorological events.
The laminar characteristic of the lagoons is the interpretive key to many other features, along with the low water renewal that is in good measure a consequence of the laminar condition. Salinity variations can take place in a short time, because of the high surface of the basin, compared to the volume; salinity can decrease due to floods and a heavy rain, while evaporation produces an increase in values, up to greatly exceed the neighboring sea values. In this context, it is understandable how very important are matter exchanges with the atmosphere.
Lagoon environments, for morphology, water renewal dynamics and exchanges at the interfaces (exchanges of matter between sediment and the water column, water and dry land, lagoon and sea waters, water surface and atmosphere), constitute real traps for nutrients, a condition that involves a eutrophic drift [1]. Though the water renewal of tidal and microtidal lagoons may have a high frequency in large areas, from 1 to a few days [2], it is equally true that the inputs of dissolved nutrients can be quickly snapped up by the submerged vegetation, so they tend to pile up. This condition is even more dramatic for non-tidal lagoon [3]. Therefore, vegetation plays an important role in nutrients cycling: they are stored, used for the growth, and returned when vegetation dies and decays to the bottom [4]. The vegetable mass can be so abundant to fill the water column completely up to surface, and when this occurs, it profoundly impacts the physical and chemical variables [5].
When the chemical energy level in the lagoon sediment, constituted by organic matter (OM) stored, exceeds the oxygenic self-depurative capacity of the ecosystem, intense anaerobic bacterial processes can occur. They are able to dissipate the energy through the release of chemical bond energy and dispersion of matter into volatile gaseous forms. These processes, for the consequences produced in the environment, are called as a whole dystrophy and can result in large die-offs of aquatic fauna. All these mechanisms are accelerated by hypertrophy of anthropogenic nature: OM storage becomes massive and dystrophic processes occur with increasingly high frequency. In such conditions, the biological communities tend to be simplified, as in extreme environments, and the reconstitution of the typical community structure becomes increasingly difficult [6].
In lagoon environments, vegetation status substantially reflects the chemical energy accumulated in the sediment top layer, a few centimetres, the ones that conduct biogeochemical exchange with the water column. Such accumulation causes instability and changes in plant assemblages, setting up vegetation cycle with extremes that go from low sediment energy levels (e.g., after a dystrophic crisis), to high levels (just before the subsequent crisis) [7, 8]. This view provides an explanation of the great sudden variations in aquatic vegetation of eutrophic, low water renewal basins, in line with the broader picture of succession proposed for the transition from oligotrophy to eutrophy in marine environments [9].
According to Figure 1, the cycle involves an increasing nutrient release from sediment, changing from nitrates to ammonium, and a seriation of more and more tolerant and opportunistic species, as sediment enriches in OM. In transitional water environments, vegetable dominance changes from seagrasses, which is the best condition, to Chlorophyceae, or, in even more extreme conditions, to microalgae and cyanobacteria [10, 11], via a series of intermediate vegetation stages, each of which prefers particular conditions of sediment OM, dissolved nutrients and N:P atomic ratio [7].
![Figure 1: Nutrient release and vegetation dominance cicles are altered by the progressive enrichment of the sediment organic matter, OM (in brown: OM increase from left to right). Arrows pointing downwards: organic matter sedimentation; arrows pointing upwards: nutrient release from the bottom (from left to right, from nitrate to ammonium dominances); hb, high biodiversity; Rh, Rhodophyceae dominance; Ch, Chlorophyceae dominance from Lenzi, et al. [12].](/fulltextimages/7060/fig_1.png)
Figure 1: Nutrient release and vegetation dominance cicles are altered by the progressive enrichment of the sediment organic matter, OM (in brown: OM increase from left to right). Arrows pointing downwards: organic matter sedimentation; arrows pointing upwards: nutrient release from the bottom (from left to right, from nitrate to ammonium dominances); hb, high biodiversity; Rh, Rhodophyceae dominance; Ch, Chlorophyceae dominance from Lenzi, et al. [12].
Mitigation Solutions
In the last forty years, eutrophication severely affected coastal lagoons, and since they are mostly heavily anthropized, public authorities have intervened in seeking solutions to improve the environmental conditions, because of the ecological, touristic and fishing importance of these ecosystems. Many solutions and examples of lagoon remediation and environmental management are available through the specialist literature. Heavy interventions to modify the basin hydrodynamics through construction of canals communicating with the sea and underwater canals, changes in internal shoreline morphology, water flow control of the hydrographic basin and even massive removal of sediments are emphasized [13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23].
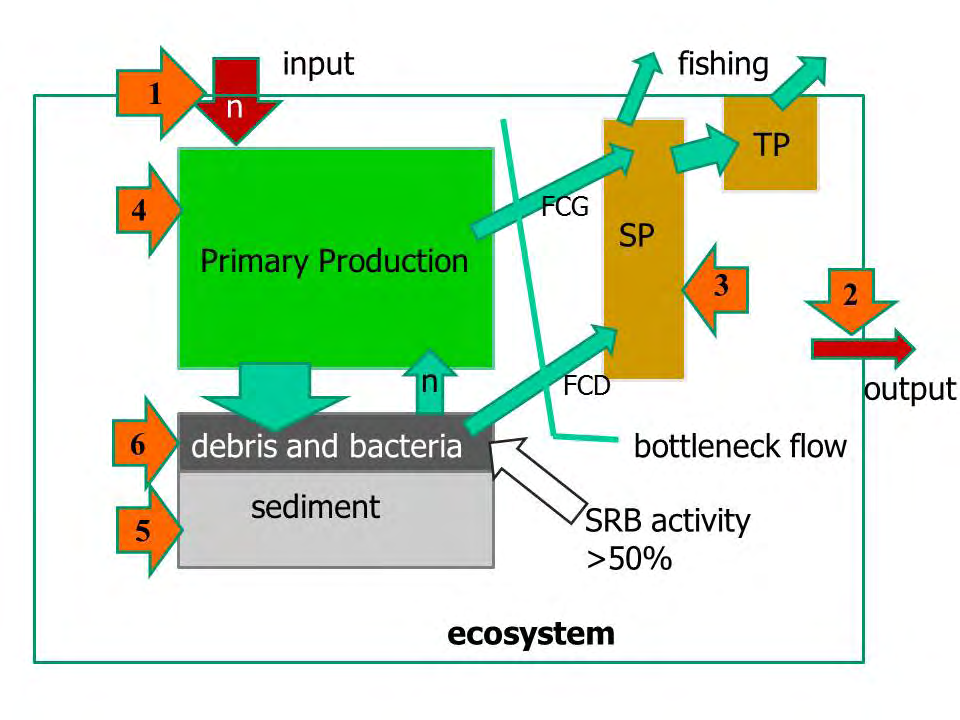
Figure 2 shows a lagoon ecosystem model that considers: a) input and output of the nutrients; b) primary, secondary and tertiary productions, considering the energy flows of the grazing and detritus chains as not effective to control vegetation and detritus productions in eutrophic systems; c) the detritus enrichment of the bottom, subject to the decay process mainly by the sulphate-reducing bacteria (SRB) action, d) the withdrawals by fisheries. According to this model, we can list 6 different modalities of intervention at different ecosystem levels to counteract dystrophic crises: 1) reduction of nutrient inputs; 2) increase in nutrient output; 3) increase of grazer and filter-feeders; 4) harvesting and removing of the macroalgal masses; 5) sediment removal to create submerged canals and to increase the depth of some critical areas; 6) actions on the anoxic top sediment layer. Applying a simple logic, to counteract the eutrophication effects, the first action to be taken would be to remove nutrient inputs (Figure 2, point 1). This can be addressed by a) treating civil wastewater with activated sludge plants, b) conveying treated municipal wastewater and shipment to other locations, c) deviating into the sea the tributary waterway of the lagoon basin that collects municipal wastewater and agricultural runoff, d) using constructed wetlands as phytotreatment and sedimentation basins of urban effluents or other sources before releasing them into the lagoon [24, 25]. The removal of nutrient inputs is not always the easiest to adopt, as the territory is very often heavily anthropized, full of urban constraints, private properties, craft and industrial activities, factory farms, and so on.
The pumping of marine or fresh waters can be a solution to counter OM accumulation in sediment and manage the summer critical issues (Figure 2, point 2). Water-flow increase should enhance nutrient output, reducing the ecosystem organic and nutrient load. This environmental management criterion was used in the Orbetello lagoon (Tuscany, Italy), a non tidal lagoon of 25 km2 1 m deep, pumping 12 m3 s-1 sea-water for 3-5 months per year. In an annual balance, it was estimated a water mass output of 110*106 m3 (30 *106 m3, by tidal flow, and 80 *106 m3, by pumping) which pushed away from the system 66.58% of N and 27.88% of P of the annual input of the anthropic sources; nevertheless, in the first 3 cm of the lagoon sediment about 108 tons of N and 11 tons of P were still estimates [3]. The pumping solution did not stop the macroalgal developments. In fact, the laminar nature of the basin never allowed a homogeneous water exchange, neither to prevent stagnation of large areas - the water pumped tended to flow along paths in a low energy, with a speed that did not exceed 1-1.5 cm s-1 – nor to obtain a remarkable improvement of the environmental conditions [26], the hydrodynamism of the basin remaining almost exclusively under the control of the wind [27, 11].
The increase of consumers at the lower levels of the food chain is another possible solution to reduce the tendency to accumulate chemical energy in the lagoon sediment (Figure 2, point 3). This solution removes from the system an usable biomass which would allow to obtain an economic return in support of management costs. However, it should be studied time after time, in relation to the climatic location of the basin, the type of fish or other herbivores that can be used commercially.
Acting directly on the macroalgal standing crops, the harvesting is a measure on which it is possible to leverage for triggering a virtuous circle in the lagoon dynamics (Figure 2, point 4). Macroalgal mass removal, which takes place through specific boats, is a “downstream” management practice to counteract the eutrophication process, common to many eutrophic transition areas around the world. In Italy, for the Venice lagoon (Adriatic Sea) and the Orbetello lagoon (Tyrrhenian Sea), the harvesting activities began almost simultaneously in the early 80s [28, 29], for the Sacca di Goro lagoon (Adriatic Sea) they began in the second half of the ‘90s, especially to safeguard the farms of the clam Ruditapes philippinarum [30]. In the ‘90s, macroalgal harvesting was undertaken in a bay on the coast of New York (USA), in the Prevost lagoon (Provence, France) and in the Tuggerah Lakes System and Peel-Harvey Estuary, Eastern and Western Australia, respectively [32, 33, 34]. Macroalgal harvesting involves a substantial financial commitment in investment and management: relatively large and properly equipped boats, staff for the operation and maintenance, fuel consumption, ground staff for the reception and transfer of the landed algal masses, trucks for mass transferring, personnel for the treatment of the material at storage sites, areas set up for reception of the masses and equipment for their handling, etc. This activity could be practiced with good results only if it were industrially possible to use the harvested matter. This is not always possible, especially in the Western world, when the algal blooms are due to opportunistic Chlorophyceae, which currently doesn’t allow their viable use [35, 36, 37]. This is a painful aspect of the macroalgal harvesting activities, for which it is necessary to transfer the masses to landfills for wastes, the whole operation entailing only costs. Macroalgal harvesting could be an effective tool in terms of mass removal and, therefore, of energy-detritus destined to accumulate in the sediments, but only if it can remove relatively high amounts compared to standing crops. To determine which is the threshold of the biomass to be removed for obtaining a concrete direct benefit from this activity, focused studies should be conducted, even because this threshold presumably changes from one environment to another, and from a macroalgal species to another. In addition, macroalgal harvesting should be managed to avoid seagrass meadows disturbance and damage.
High expectations are usually placed on beneficial effects that should result in the excavation of submerged canals (Figure 2, point 5), in the belief, mostly preconceived because rarely supported by specific studies, that through them an increase of the lagoon hydrodynamics can be obtained. Canals can be functional for tidal and microtidal lagoons (even if, in this case, natural channels, already exist), but this is not so obvious for the non-tidal lagoons, whose hydrodynamic is dominated only by the wind. In non- tidal environments, a submerged artificial canal tends to accumulate clay, silt, organic detritus and floating vegetation masses, resulting a depression, an area of low energy of the bottom. The filling process of the canal is inexorable and can determine prohibitive environmental conditions near the bottom, which is finally an area of nutrient and hydrogen sulphide releases. This process can be also very fast, in relation to the canal exposure to the wind, but it is certainly faster than the times it takes to start the canal maintenance. Similarly, thinking to change the morphology of a non-tidal basin by trying to improve the hydrodynamics, is equivalent to believe in the existence of important tidal flows and real currents in these ecosystems.
Many works of heavy environmental transformation have been conducted in the past to remedy conditions of frequent dystrophic crisis and for heavy industrial contamination problems [16], the result is always a deep denaturation of the original environment, with losses of biodiversity and habitats of important naturalistic value [38].
Finally, an action can be carried out on the anoxic, high organic load sediments, wholly downstream of the chain of events caused by eutrophication (Figure 2, point 6). Sediments constitute the substrate that recirculates nutrients and nourishes the anaerobic bacteria, especially SRB, for which the dystrophic processes and die-offs of fish can occur. Action on the sediment can be preventative or delayed. It is preventive when the intervention try to avoid anaerobic processes by oxidizing the sediment and reducing the organic load, it is delayed when the intervention try to block the SRB action on the border line of the environmental crisis.
Actions on the Sediment Anoxic Top Layer
Resuspension of the normally steady soft sediments (SR) of a non tidal lagoon can be caused by a lot of natural and artificial events. E.g., among natural events, storms and strong wind, stormwater runoff and floods, bioturbation and fish disturbance are listed, while among anthropogenic events, boat traffic, fishing, dredging and positioning of pipelines are the most common. Here I intend to focus on those events which disturb the soft sediment recently settled, maximum of 1-5 years, based on their variable sedimentation rate. These are the first centimeters of the sediment layer, at most 4-6 cm. Therefore, there is less interest in the dredging, which impacts on different thicknesses of sediment and can move historical storage, well-consolidated, that might never have affected otherwise the water column. Instead, the effects of wind, fish schools moving, bioturbation and bioirrigation by infauna, and boat traffic can be very important in shallow water.
It was hypothesized that in eutrophic/ipertrophic lagoons, seagrasses are confined to areas subject to high bottom shear velocity by wind, and that from there they can resume repopulating areas with low values of this variable when the dystrophic stress abates; essentially, the wind influences the distribution of these meadows through sediment oxidation, leading to sediment resuspension and OM transport [27]. Furthemore, it was shown that frequent strong winds can reduce organic load in deeper parts of the lagoons [11]. Experimental studies suggest decomposition of sediment labile organic matter (LOM) is independent of redox values, but periodic alternation of exposures to oxygen leads to a more rapid decomposition of the organic matter, than in stable condition [39]. SR can have a significant impact on the water body by lifting to the surface particulate organic matter (POM) and dissolved organic matter (DOM) [40], activating bacterial oxidative mineralization and increasing the remineralization rate [41, 42]. This phenomenon can show contradictory results, much depending on the nature of the sediments and bottom conditions, the height of the water column, the quality of the settled OM, the environmental conditions and seasons under which disturbance occurs [43, 44]. When SR occurs with a relatively high frequency, the final effects may be quite different from those of occasional events. A SR event that occurs in a eutrophic environment with low frequency (e.g., 20 times per year) could allow the release in the water column of pore-water nutrients, favoring the sudden development of opportunistic micro or macroalgal species. In addition, the sudden organic load (detritus and dissolved molecules) that reaches the water column could produce a DO decrease, especially in the summer period when it is lower and the temperature does not allow to maintain high values.
A different situation is created in the case of frequent resuspension, which, after a first “negative” effect, does not allow that it can be repeated because, resuspension by resuspension, the sediment quality changes. A laboratory study [45] proved frequent SR increased the mineralization rate with respect to undisturbed sediment by a factor between 2 and 5. This study showed that the frequent turbulence affecting surface sediment (e.g.: 24-48 h) produces an increase in the mineralization rate, without showing any significant effect on the water column.
An experience conduced in field [46] demonstrated that repeated passages of boats on a limited area of a shallow non- tidal lagoon to resuspend sediment soft top layer, increased the sediment oxidative status and decreased the organic content, without any significant increase in nutrients or oxygen consumption in the water column. The same results were confirmed in two areas (20 hectares each) of another non-tidal lagoon after a frequently repeated disturbance of sediments by a fitted boat; moreover, by trend of dissolved sulphides, it was highlighted the disturbance interrupted sulphate reduction processes [47].
In a study that examined the interaction between physical disturbance and organic enrichment in relation to the effects on diversity and macroinfaunal assemblages, Widdicombe and Austen [48] observed that diversity was low when physical disturbance was low and sediment organic quantity was high, and when the frequency of disturbance was high and sediment organic quantity was low. On the contrary, diversity was high when both disturbance and organic enrichment were high or low. In essence, a low- water-renewal – low-hydrodynamic-environment, such as a not-tidal lagoon, tends to an increasingly low biodiversity as sediment OM increases. Martelloni, et al. [49] confronted high organic rich sediment of a lagoon between undisturbed condition and artificial sediment disturbance (frequency=60 times in 12 months). They confirmed a close relationship between the number of zoobenthic species in the sediment and organic load, showing a relative increase in the number of species in disturbed sediment.
The idea of using frequent SR to counteract the harmful effects of eutrophication in shallow water environments, found initial support in the reduction of the sediment organic load. But this is not the only possible effect (Figure 3).
![Figure 3: A conceptual model for sediment resuspension consequences from Lenzi and Porrello [55].](/fulltextimages/7060/fig_3.png)
In sediments with a prevailing oxidative status, orthophosphates are bound to ferric oxides-hydroxides and to carbonates and clays, becoming increasingly rare in interstitial waters and less and less present into the water column, and finally, unavailable to algae, creating P-limitation conditions. Moreover, oxidation accelerates nitrification and nitrates become predominant over the reduced forms [50]. An increase in nitrate concentrations produces an increase in denitrification [51], which occurs in anoxic microhabitats (e.g.: inside microparticles) [52]. Thereby, part of the nitrogen load is lost as N2 or N2O and the eutrophy of the system can decline [53], unlike what happens in totally anaerobic processes, mostly nutrient retentive, in which N is not lost from the system [54]. Summing up, a frequent top layer SR can produce: increase of sediment redox, reduction of LOM, P limitation and partial loss of N. In the conceptual model shown in Figure 3, frequent SR establishes a virtuous circle, despite the eutrophic potential of the basin remains, in a sort of downstream eutrophication management.
All this helps to contain macroalgal developments, even if much depends on the commitment and operating frequency, while the development of sea grasses is favored, since these last can find the nutrients directly into the substrate through the root system, and no longer suffer from suffocation by macroalgal floating masses.
SR can be practiced in several ways. In the Orbetello lagoon, SR has been conducted with the same macroalgal harvesting boats (13 m x 4.5 m; 13 tons capacity). The boats, while sailing, determine a thrust on the water mass resuspending the soft sediment (3-5 cm), which consists in the direct effect on the sediment [46], moreover they produce an indirect effect equally important, that is the resuspended sediment fall out on the surrounding bottom [46, 47, 48, 49, 50, 51, 52, 53, 54, 55]. The first effect is related to boat size, basin depth, sediment type and boat thrust. Snorkel divers measured the trace left in sediment by boats of the type used for harvesting, establishing that each boat displaces the top sediment in a belt about 6 m wide [46]. It was estimated that more than 30% of the matter was still suspended at an orthogonal distance of 50 m from the boat path, the total fall-out of resuspended sediment happened within a hundred meters. The total resuspended sediment, considering the two boards of the boat, amounted to 4.026 kg for meter of boat path. In total, in six months of algal harvesting activity (6,000 tonnes year-1 landed), 16,589 tons of sediment top-layer were handled [55]. Compared to macroalgae harvested, this figure suggests that sediment resuspension was the major effect, in terms of mass, of the harvesting boats.
A specially boat was used to disturb and resuspend the sediment top layer in the Burano lagoon, 120 ha 1 m deep (Tuscany, Italy). The boat (4.70 m x 1.60 m) was equipped with an outboard motor and a generator to power two 0.75 kW motors having shafts terminating in propellers, usually used to oxygenate aquaculture ponds. The motors were mounted on steel frames on either side of the hull so that the shafts could be inclined at different angles. When the motors were in action they produced a jet of water and air directed towards the bottom, and could be inclined by varying the angle of the motor shaft in relation to the speed of the vessel and depth. Disturbance was carried out for 3 months in two areas for a total of 44 ha. The impact of daily SR on the water column was monitored monthly in the disturbed areas and in two undisturbed control areas 20 ha each. Results showed a stable OM content in disturbed areas and an significant increase in undisturbed areas, as well as an increase in macroalgae in these latter areas. Seagrass meadows increased greatly in disturbed areas, where seedlings of Ruppia cirrhosa were observed for the first time in many years. No significant N–NH4, SRP, DO or pH changes on the water column were observed due to SR [47]. In the same field experience, Lenzi and Renzi [57] showed that 1) the higher abatement of LOM was recorded in the disturbed area; 2) C:N ratio of sediment was much lower in the undisturbed areas than in the disturbed areas; 3) sediment and macroalgal tissue C:N ratios did not significantly change in response to disturbance; 4) initial sediment OM enrichment from phytoplankton was greater in disturbed (water clarification) than undisturbed areas ; 5) chlorophyll-a, phaeopigments and the pigment diversity index did not reflect any decline in photosynthesis in the disturbed areas; 6) treatment effects increased with increasing frequency of disturbance. These findings indicated less availability of LOM and therefore lower risk of extended anoxic events in disturbed areas, and supported the hypothesis that artificial disturbance of sediment can be used to manage lagoon environments and prevent eutrophication consequences. Finally, this study showed sediment LOM load and macroalgal mats decreased significantly when sediment resuspension and high-density macroalgal mat disturbance occur for a long period with high frequency.
Possible Effects of the Sediment Resuspension on Bio-Availability of Contaminants
Resuspension of contaminated sediments can determine contaminants transportation downstream and affect the water column for a variable time. Contaminants can be essentially heavy metals, polycyclic hydrocarbons (PAHs), and polychlorinated biphenyls (PCBs).
In anoxic sediments, heavy metals are normally bound as sulphides, with very low solubility [58], while in the aerobic sediment layer, heavy metals can be bound to OM [59] and iron and manganese oxi-idroxides [60]. Resuspension of anoxic sediments could result in the oxidation of sulphides and heavy metal cations releases. Some sulphides can rapidly oxidize in a well-aerated water column [61], however, when this occurs, some conditions of the water column and the sediment top layer could favor metal adsorption, thus, as a result of sedimentary suspension, it could paradoxically be obtained a concentration of heavy metal cations lower than before [62]. In numerous studies, the contact between heavy metal sulphides and dissolved oxygen in the water column did not show significant Cu, Zn, Cd, Pb cation releases, despite significant Fe and Mn releases due to sulphides oxidation; according to Simpson, et al. [61], dissolved heavy metal cations can coprecipitate with Fe and Mn oxy-hydroxides or be complexed by OM. Moreover, during resuspension trace metal sulphides may remain unoxidized because of buffering by FeS and MnS oxydation, that are present in large excess with respect to other metal sulphides [61].
Mercury (Hg) is also chemically bound in the oxidized sediment to OM and Fe and Mn oxy-hydroxide, while in anoxic sediment it is found mostly as highly insoluble sulphide. Microbial degradation of OM and chemical dissolution of sulphide can release Hg. Moreover, SRB activity can release Hg by methylation process from HgS, in OM rich sediment [63] (Figure 4) producing methylmercury (MeHg), volatile, which can easily enter the food chain and be retained by lipid.
![Figure 4: Diagram (concentration to the ordinates, and varying environmental conditions to the abscissas) shows hypothetical sediment conditions from the oxidized state (left), to an increasingly reduced anoxic state, proceeding to the right on the abscissas. The trend of redox potential (Eh in mV; black line) goes towards increasingly negative values (from left to right) with the gradual decrease of dissolved oxygen, in conjunction with the increase in the load of organic matter in sediments (OM; brown line). As this situation proceeds towards anoxia/dystrophy, sulphate-reducing bacteria activity increases, therefore acid-volatile sulphides (AVS) concentration increases (yellow line). Sulfate-reducing bacteria operate the methylation of Hg. This last procedure increases with increasing bacterial activity, until the concentration of the AVS becomes high enough to inhibit the process (trend of the gray line). Therefore, two moments are identified in which the concentration of MeHg is the lowest or zero (blue circles), and an intermediate phase in which this concentration is at the highest values (red circle), which could coincide mainly with the intermediate seasons from Lenzi, et al. [64].](/fulltextimages/7060/fig_4.png)
Figure 4: Diagram (concentration to the ordinates, and varying environmental conditions to the abscissas) shows hypothetical sediment conditions from the oxidized state (left), to an increasingly reduced anoxic state, proceeding to the right on the abscissas. The trend of redox potential (Eh in mV; black line) goes towards increasingly negative values (from left to right) with the gradual decrease of dissolved oxygen, in conjunction with the increase in the load of organic matter in sediments (OM; brown line). As this situation proceeds towards anoxia/dystrophy, sulphate-reducing bacteria activity increases, therefore acid-volatile sulphides (AVS) concentration increases (yellow line). Sulfate-reducing bacteria operate the methylation of Hg. This last procedure increases with increasing bacterial activity, until the concentration of the AVS becomes high enough to inhibit the process (trend of the gray line). Therefore, two moments are identified in which the concentration of MeHg is the lowest or zero (blue circles), and an intermediate phase in which this concentration is at the highest values (red circle), which could coincide mainly with the intermediate seasons from Lenzi, et al. [64].
SRBs require low or no presence of oxygen in the sediments, and sufficient OM load to start methylation process. Sediment oxygenation in low OM load decreases methylation [65]. When the SRB activity becomes predominant, as in the full-blown dystrophic conditions, it can lead to enhanced porewater dissolved sulphide, which has been shown to inhibit Hg methylation [66, 65] (Figure 4). According to Kim, et al. [67], MeHg resulted significantly and negatively correlated with acid-volatile sulphides (AVS) concentration, suggesting increased Hg methylation was coupled with AVS oxidation. It is therefore possible to hypothesize a range of environmental conditions that goes from one extreme, where sediments are partially oxidized and the sulphate reducing activity is low or zero and so is methylation, at the opposite extreme, that is an anoxic/dystrophic state with very intense SRB activity, enrichment in AVS and low or no methylation. In an increasing SRB activity gradient between the two extremes, we can imagine an increase in methylation, up to a peak more or less halfway (Figure 4). Because extreme dystrophic conditions are not desirable to avoid methylation, the choice of the environmental manager should therefore be oriented towards achieving a sediment low OM load, relatively high Eh values, low SRB activity, consequently, low or no methylation.
SR can induce a change in sediment redox status, and can have an impact by changing the association of Hg with binding phases, both effects can be important factors determining the rate of Hg methylation in sediments [67]. Ultimately, if it is true SR could enhance methylation by decreasing AVS levels, it is even more decisive that SR may limit methylation by sediment oxidation, limiting SRB activity [67].
In the Orbetello lagoon, there is an excessive accumulation of Hg in commercial fish species, in particular sea bass and sea bream. There is a high presence of cinnabar in the sea of the South Tuscany due to the washing away of debris from Amiata Month, and the values are even higher in the lagoon than in the nearby sea [68]. Lenzi, et al. [55] observed that, paradoxically, smaller fish were more likely to accumulate Hg than larger ones. They hypothesized the metal accumulation in fish depended more on contact with the contaminant released by the anoxic sediment, than on the food chain. In this lagoon, in fact, SRB activity can be very intense in the hot season, with abundant release of hydrogen sulphide, but subtly can release greater quantities of MeHg in the intermediate seasons, when bacterial activity and the presence of AVS are relatively lower. Il was showed that, using diffusive gradient in thin films technique (DGT), resuspension of sediments did not increase the availability of HgII in disturbed areas, compared to non-disturbed areas [55].
PAHs are present in marine environment sometimes in
relevant concentrations, both of pyrogenic and petrogenic origin. The pyrogenic PAHs are composed mainly of larger ring systems then the petrogenic. PCBs (C12H10-xClx) are artificial molecules. They have been replaced long ago because they have been recognized carcinogens, but they are still widely spread in the world.
PCBs and PAHs tend to be associated with particulate organic matter. SR can involve the release of these contaminants. Desorption is very dependent on the molecules, the nature of the binder particle and the environmental conditions. SR, however, exposes the organic contaminants to oxidation and UV action, which are more effective the higher is the frequency of SR. Le Blanc, et al. [69] showed that degradation of phenanthrene resulted greater in resuspension conditions that in undisturbed sediments. DO, UV and the specific bacterial population that is produced in such conditions contribute directly. Regarding PCB, the water column redox did not have a significant effect on partitioning from organic particles [70]. Nevertheless, by exposing anoxic sediment to DO, partitioning of a PCB between pore water DOM and freely dissolved DOM decreased for pore water sediment of two estuarine sites out of three [71].
References
-
McLusky DS, Elliott M (2004) The Estuarine Ecosystem. Ecology, Threats and management. Oxford Uni Press, pp: 214.
-
Viero D, Defina A (2016) Renewal time scale in tidal basin: Climbing the Tower of Babel. Sustainable Hydraulics in the Era of Global Change-Erpicum, Taylor & Francis Group, London.
-
Palmieri R, Porrello S (2003) Restoration of the eutrophic Orbetello lagoon (Tyrrhenian Sea, Italy): water quality management. Mar Pollut Bull 46(12): 1540-1548.
-
Sfriso A, Marcomini A, Pavoni B (1987) Relationships Between Macroalgal Biomass and Nutrient Concentrations in a Hypertrophic Area of the Venice Lagoon. Marine Environmental Resources 22(4): 297- 312.
-
Lenzi M, Gennaro P, Mercatali I, Persia E, Solari D, et al. (2013) Physico-chemical and nutrient variable stratifications in the water column and in macroalgal thalli as a result of high biomass mats in a non-tidal shallow-water lagoon. Mar Pollut Bull 75(1-2): 98-104.
-
Raffaelli DG, Raven JA, Poole LJ (1998) Ecological impact of green macroalgal blooms. Oceanography and Marine Biology: an Annual Review 36: 97-125.
-
Lenzi M, Gennaro P, Renzi M, Persia E, Porrello S (2012) Spread of Alsidium corallinum C. Ag. in a Tyrrhenian eutrophic lagoon dominated by opportunistic macroalgae. Marine Pollution Bulettin 64(12): 2699- 2707.
-
Lenzi M (2014) Vegetation Structure of Lagoon Environments As Result of Sediment Biogeochemical Dynamics and Organic Matter Load: A Review. In: Macrophytes. Biodiversity, Role in Aquatic Ecosystems and Management Strategies, pp: 45-58.
-
Duarte CM (1995) Submerged aquatic vegetation in relation to different nutrient regimes. Ophelia 41(1): 87- 112.
-
Sorokin IY, Sorokin PY, Gnes A (1996) Structure and functioning of the anthropogenically transformed Comacchio lagoonal ecosystem (Ferrara, Italy). Mar Ecol Prog Ser 133: 57-71.
-
Lenzi M, Persiano M, Gennaro P, Rubegni F (2016) Wind Mitigatin Action on Effects of Eutrophication in Coastal Eutrophic Water Bodies. International Journal of Marine Science and Ocean Technology 3(2): 14-20.
-
Lenzi M, Gennaro P, Renzi M, Persia E, Porrello S (2012) Spread of Alsidium corallinum C. Ag. in a Tyrrhenian eutrophic lagoon dominated by opportunistic macroalgae. Marine Pollution Bulettin 64(12): 2699- 2707.
-
Murakami K (1984) Dredging for controlling eutrophication of lake Kasumigaura, Japan. Lake and Reservoir Management 1(1): 592-598.
-
Van Berk AH, Oostinga H (1992) North lake of Tunis and its shores: restoration and development. Terra Aqua 49: 23-32.
-
Bettinetti A, Pypaert P, Sweerts JP (1996) Application of an Integrated management Approach to the Restoration Project of the Lagoon of Venice. Journal of Environmental management 46(3): 207-227.
-
Vandenbroeck J, Charrada BR (2001) Restoration and Development Project of South Lake of Tunis and its Shores. Terra et Aqua 85: 11-20.
-
Jin W, Webb C (2003) Experience of creek and lagoon restoration feasibility study. International Conference on Estuaries and Coasts. Hangzhou, China, pp: 708-711 .
-
SCE (2005) San Dieguito Wetlands Restoration Project. Final Restoration Plan. Southern California Edison Company, pp: 260 .
-
Newton A, Mudge SM (2005) Lagoon-sea exchange, nutrient dynamics and water quality management of the Ria Formosa (Portugal). Coastal and Shelf Science 62(3): 405-414.
-
Deheyn DD, Shaffer LR (2007) Saving Venice: Engineering and ecology in the Venice lagoon. Technology in Society 29(2): 205-213.
-
Tsihirintzis VA, Sylaios GK, Sidiropoulou M, Koutrakis ET (2007) Hydrodynamic modelling and management alternatives in a Mediterranean, fishery exploited, coastal lagoon. Aquacultural Engineering 36(3): 310- 324.
-
Ponti M, Pasteris A, Guerra R, Abbiati M (2009) Impacts of maintenance channel dredging in a northern Adriatic coastal lagoon. II: Effects on macrobenthic assemblages in channels and ponds. Estuarine, Coastal and Shelf Science 85(1): 143-150.
-
Wijnhoven S, Escaravage V, Daemen E, Hummel H (2009) The Decline and Restoration of a Coastal Lagoon (Lake Veere) in the Dutch Delta. Estuaries and Coasts 33: 1261-1278.
-
Lenzi M, Mattei N, Solimeno P ( 1998) L a g o o n i n g of depurated civil effluents in a specifically enclosed, marginal area of the Orbetello Lagoon (Tuscany, Italy). Biologi Italiani 28(3): 23-28.
-
Gennaro P, Funari F, Guidotti M, Mercatali I, Porrello S, et al. (2006) Reduction of land based fish-farming impact by phytotreatment pond system in marginal lagoon area. Aquaculture 256(1-4): 246-254.
-
Innamorati M, Melillo C (2004) Study of the Orbetello lagoon. Ecology and economic aspects. Dept of Ecol Vegetale, University of Florence, pp: 199.
-
Rubegni F, Franchi E, Lenzi M (2013) Relationship between wind and seagrass meadows in a non-tidal eutrophic lagoon studied by a wave exposure model (WEMo). Mar Pollut Bull 70(1-2): 54-63.
-
Lenzi M (1992) Experiences for management of Orbetello Lagoon: eutrophication and fishing. Marine Coastal Eutrophication, pp: 1189-1198.
-
Runca E, Bernstein A, Postma L, Di Silvio G (1996) Control of macroalgae blooms in the Lagoon of Venice. Ocean Coast Manag 30(2-3): 235-257.
-
De Leo GA, Batoli M, Naldi M, Viaroli P (2002) A First Generation Stochastic Bioeconomic Analysis of Algal Bloom Control in a Coastal Lagoon (Sacca di Goro, Po River Delta). Marine Ecology 23 (1): 92-100.
-
Cotroneo C, Yozzo D, Doss T, Will B, McLaughlin J (2010) Pilot Project to Harvest Sea Lettuce Removal of Ulva lactuca from Jamaica Bay Restore America’s Estuaries.
-
Barre N (2005) Un moyen de lutte contre l’eutrophisation d’une lagune: Agir sur le stock d’algues. CEN LR Pôle Lagunes. Réseau de Suivi Lagunaire-Séminaire eutrophisation Montpellier, pp: 12.
-
Guyoneaud R, De Wit R, Matheron R, Caumette P (1998) Impact of macroalgal dredging on dystrophic crises and phototrophic bacterial blooms (red waters) in a brackish coastal lagoon. Oceanologica Acta 21(4): 551-561.
-
King RJ, Hodgson BR (1995) Tuggerah Lakes system, New South Wales, Australia. In: McComb AJ (Ed.), Eutrophic Shallow Estuary and Lagoons. CRC Press, Boca Raton, Florida, pp: 19-29.
-
Bastianoni S, Tiezzi E, ColacevichA, Borghini F, Coppola F, et al. (2008) Biofuel potential production from the Orbetello lagoon macroalgae: a comparison with sunflower feedstock. Biomass Bioenerg 32(7): 619-628.
-
Migliore G, Alisi C, Sprocati AR, Massi E, Ciccoli R, et al. (2012) Anaerobic digestion of macroalgal biomass and sediments sources from the Orbetello lagoon, Italy. Biomass and Bioenergy 42: 69-77.
-
Schultz-Jensen N, Thygesen A, Leipold F, Thomsen ST, Roslander C, et al. (2013) Pretreatment of the macroalgae Chaetomorpha linum for the production of bioethanol- Comparison of five pretreatment technologies. Bioresource Technology 140: 36-42.
-
Zacharias I, Gianni A (2008) Hydrodynamic and dispersion modeling as a tool for restoration of coastal ecosystems. Application to a re-flooded lagoon. Environmental Modelling & Software 23(6): 751-767.
-
Kristensen E, Holmer M (2001) Decomposition of plant materials in marine sediment exposed to different electron acceptors (O2, NO3 -, and SO4 2-), with emphasis on substrate origin, degradation kinetics, and the role of bioturbation. Geochimica et Cosmochimica Acta 65(3): 419-433.
-
Hopkinson CS (1985) Shallow-water benthic and pelagic metabolism: evidence of heterotrophy in the nearshore Georgia bight. Mar Biol 87: 19-32.
-
Wainright SC (1987) Stimulation of heterotrophic microplankton production by resuspended marine sediments. Science 238(4834): 1710-1712.
-
Wainright SC (1990) Sediment-to-water fluxes of particulate material and microbes by resuspension and their contribution to the planktonic food web. Mar Ecol Prog Ser 62: 271-281.
-
Tengberg A, Almroth E, Hall P (2003) Resuspension and its effects on organic carbon recycling and nutrient exchange in coastal sediments: in situ measurements using new experimental technology. Journal of Experimental marine Biology and Ecology 285-286: 119-142.
-
Arnosti C, Holmer M (2003) Carbon cycling in a continental margin sediment: contrasts between organic matter characteristics and remineralisation rates and pathways. Estuarine, coastal and shelf science 58(1): 197-208.
-
Stahlberg C, Bastviken D, Svensson BH, Rahm L (2006) Mineralisation of organic matter in coastal sediment at different frequency and duration of resuspension. Estuarine, Coastal and Shelf Science 70(1-2): 317-325.
-
Lenzi M, Finoia MG, Persia E, Comandi S, Gargiulo V, et al. (2005) Biogeochemical effects of disturbance in shallow water sediment by macroalgae harvesting boats. Marine Pollution Bulletin 50 (5): 512-519.
-
Lenzi M, Birardi F, Calzolai R, Finoia MG, Marcone F, et al. (2010) Hypertrophic lagoon management by sediment disturbance. Mar Pollut Bull 61(4-6): 189-197.
-
Widdicombe S, Austen MC (2001) The interaction between physical disturbance and organic enrichment: An important element in structuring benthic communities. Limnol Oceanogr 46(7): 1720-1733.
-
Martelloni T, Tomassetti P, Gennaro P, Vani D, Persia E, et al. (2016) Artificial soft sediment resuspension and high density opportunistic macroalgal mat fragmentation as method for increasing sediment zoobenthic assemblage diversity in a eutrophic lagoon. Marine Pollution Bulletin 110(1): 212-220.
-
Revsbech NP, Sorensen J, Blackburn TH, Lomholt JP (1980) Distribution of oxygen in marine sediments measured with microelectrodes. Limnol Oceanogr 25(3): 403-411.
-
Herbert RA, Nedwell DB (1990) Role of environmental factors in regulating nitrate respiration in intertidal sediments. In: Denitrification in Soil and Sediment, pp: 56: 77-90.
-
Fenchel T (1992) Aspects of decomposer food chains in marine benthos. Ver Deutsh Zool Ges 65: 14-22.
-
Novicki BL, Requintina E, Kueren VD, Kelly JR (1997) Nitrogen losses through sediment denitrification in Boston Harbour and Massachusetts Bay. Estuaries 20: 626-639.
-
Moeller K, Mueller T (2012) Effects of anaerobic digestion on digestate nutrient availability and crop growth: A review. Eng Life Sci 12(3): 242-257.
-
Lenzi M, Porrello S (2017) Non-tidal lagoon structure, dynamics and eutrophication management. Lambert Academic Publishing.
-
Lenzi M, Finoia MG, Gennaro P, Mercatali I, Persia E, et al. (2013) Assessment of resuspended matter and redistribution of macronutrient elements produced by boat disturbance in a eutrophic lagoon. J Environ Manage 123: 8-13.
-
Lenzi M, Renzi M (2011) Effects of artificial disturbance on quantity and biochemical composition of organic matter in sediments of a coastal lagoon. Knowl and Manag of Aquatic Ecosystems 8: 17.
-
Simpson SL, Apte SC, Batley GE (2000) Effect of Short- Term Resuspension Events on the Oxidation of Cadmium, Lead, and Zinc Sulfide Phases in Anoxic Estuarine Sediments. Environ Sci Technol 34(21): 4533-4537.
-
Chapman MP, Wang F, Janssen C, Persoone G, Allen EH (1998) Ecotoxicology of metals in aquatic sediments: binding and release, bioavailability, risk assessment, and remediation. Canadian Journal of Fisheries and Aquatic Sciences 55(10): 2221-2243.
-
Davrance M, Bollinger JC (2000) Heavy Metals Desorption from Synthesized and Natural Iron and Manganese Oxyhydroxides: Effect of Reductive Conditions. J Colloid Interface Sci 227(2): 531-539.
-
Simpson SL, Apte SC, Batley GE (1998) Effect of Short- Term Resuspension Events on Trace Metal Speciation in Polluted Anoxic Sediments. Environmental Science & Technology 32(5): 620-625.
-
Forstner U (1995) Non-linear release of metal from aquatic sediments. Salomones W. And Stigliani W.M. Eds. Biogeodynamics of pollutants in soil and sediments, pp: 247-307.
-
Benoit J, Gilmour CC, Heyes A, Mason R, Miller C (2003) Geochemical and biological controls over methylmercury production and degradation in aquatic systems. Biochemistry of Environmental Important Trace Elements 19: 262-297.
-
Lenzi M, Franchi E, Leporatti-Persiano M, D’Agostino A, Gennaro P, et al. (2021) Assessment of the causes of Hg bioaccumulation in the fish of a Mediterranean lagoon subject to environmental management interventions. Mar Pol Bull 162: 111907.
-
Benoit JM, Gilmour CC, Mason RP, Heyes A (1999) Sulfide controls on mercury speciation and bioavailability to methylating bacteria in sediment pore waters. Environmental Science & Technology 33(6): 951-957.
-
Gilmour CC, Riedel GS, Ederington MC, Bell JT, Gill GA, et al. (1998) Methylmercury concentrations and production rates across a trophic gradient in the northern Everglades. Biogeochemistry 40(2-3): 327-345.
-
Kim EH, Mason RP, Porter ET, Soulen HL (2006) The impact of resuspension on sediment mercury dynamics, and methylmercury production and fate: a mesocosm study. Marine Chemistry 102(3-4): 300-315.
-
Miniero R, Beccaloni E, Carere M, Ubaldi A, Mancini L (2011) Merchry (Hg) and methyl mercury (MeHg) concentration in fish from the coasta lagoon of Orbetello central Italy. Marine Pollution Bulletin 76(1-2): 365-369.
-
LeBlanc LA, Gulnick JD, Brownawell BJ, Taylor GT (2006) The influence of sediment resuspension on the degradation of phenanthrene in flow-through microcosms. Mar Environ Res 61(2): 202-223.
-
Friedman LC, Lohmann R, Burgess RM, Perron MM, Cantwell MG (2011) Resuspension of polychlorinated biphenyl-contaminated field sediment: release to the water column and determination of site-specific KDOC. Environmental Toxicology and Chemistry 30(2): 377- 384.
-
Pedersen JA, Gabelich CJ, Lin CH, Suffet IH (1999) Areation effects on the partitioning of a PCB to anoxic estuarine sediment pre water dissolved organic matter. Environ Sci Technol 33(9): 1388-1397.
- Genetic Improvement of Nile Tilapia (Oreochromis niloticus): Advances in Selective Breeding and Genomic Approaches for Sustainable Aquaculture
- Microplastics, Contaminants, and Waste Hotspots: Divergences and Faults in Prioritizing Control Efforts
- Creating a Healthier, More Vibrant Open and Closed Aquatic Environment. A Submersible, Centrifugal Magnetically Affixed Current Changing Aquarium Pump
- An Attempt to Assess Alpha Diversity and Sample Size: Using the Ostracod Assemblages off Kumamoto Port, Japan
- Assessment of the Efficiency of Common Fishing Gears and Crafts Used at Mohananda River of Chapai Nawabganj, Bangladesh
- Fish Productivity and Biodiversity Status of Sundarban Mangrove in Bangladesh