Fungal Pectinase Production Optimization and its Application in Buffaloe’s Diets Degradation
Pectinase production for improving buffalo’s diets digestion is the main objective of this work. Effects of fungal strains and different cultivation conditions on pectinase production have been studied. In vitro batch culture technique was used for investigate impact of the produced pectinase compared with commercial pectinase (SMIZYME®) on rumen fermentation parameters and diet degradation. Penicillium chrysogenum exhibited the highest pectinase activity at 3 days of incubation period, initial pH 4 of the growth medium, yeast extract as a sole nitrogen source and pomegranate peel as a carbon source at a concentration of 15 % (W/V). Three (g/kg) of the both enzymes supplementation significantly increased treated diet’s dry matter (DM), neutral detergent fiber (NDF), acid detergent fiber (ADF) degradability with increase total gas production (TGP) and short chain fatty acids (SCFA) concentration. The enlargement of pectinase production locally will lead to animal production improvement, encourage self-reliance and reduce the cost of enzymes importation.
Introduction
Pectinase is a generic name group of enzymes that catalyze break down of the glycosidic bonds of the galacturonic acid long chain residues in the pectin rich plants [1]. Pectinases make up almost 25% of the global food enzyme market because of its wide range of applications in food, feed and pharmaceutical industries Int J Zoo Animal Biol
[2]. It is expected that value of food enzymes market will increase up to $ 41.4 billion by 2020, with a compounded annual growth rate of 6.7% [3]. The major constraint for commercialization of new sources of enzymes is higher cost of the production [4]. It is thought that define the optimal microbial cultivation conditions for capable microbial strains and cheap raw substrate may reduce the cost of enzymes production [5]. A large number of bacteria, yeasts and many filamentous fungi are potential pectinase producers [6]. Fungi (ex: T. viride, A. flavus, A. niger, F. oxysporum, A. terreus, and P. chrysogenum) have attracted the most attention as enzymes producers because of its prolific yield and its long history in fermentation industries [1, 5, 7, 8, 9]. Submerged fermentation technique is still the most favorable system for microbial enzymes production in large-scale, despite a lot of research papers recommended solid state fermentation for microbial enzymes (ex: pectinase) production [1, 7]. Utilization of agricultural residues as carbon sources in pectinase production media has yielded good enzyme activity with reduction of the production cost [6, 7].
Utilization of microbial pectinases in agriculture waste treatments or as livestock feed supplements was expanded in the last two decades [6]. Many studies reported good impact of pectinase as a fibrolytic enzyme on ruminant’s diets degradation in vitro and in vivo [4, 10, 11, 12]. Addition of pectinase to ruminant's feeds can reduces the feed viscosity, which increases absorption of nutrients, liberates nutrients, either by hydrolysis of non- biodegradable fibers or by liberating nutrients blocked by these fibers, and reduces the amount of feces [7]. It is well known that pectin constitute the main component of cell wall of orange peel, sugar beet pulp, and pomegranate peel, that become essential source for livestock feeding in Egypt. Therefore, this study was carried out to optimize production of fungal pectinase, and evaluates impact of the produced pectinase for improving degradation of buffaloe’s diets rich in pectin.
Material and Methods
Fungi and Inoculum Preparation
Fusarium avenaceum, Asperigillus fugimatus, Cephalosporium acremonium, Trichoderma viride, Aspergillus niger, Fusarium oxysporum, Aspergillus terreus, and Penicillium chrysogenum were obtained from Dairy Microbiology LAB - National Research Centre, Giza, Egypt. These fungi were cultivated and maintained on potato dextrose agar medium (PDA). Malt medium containing malt extract (30 g/l); yeast extract (5 g/l) was used for preparing the activated fungal inocula; beet pulp powder medium (BPPM) was used for growth and pectinase production. The medium has the following composition (g/l) NaCl, 6.0; (NH4)2SO4, 1.0; K2HPO4, 1.0; MgSO4, 7H2O, 0.05; CaCl2, 0.1; yeast extract, 0.5; peptone, 0.5; glucose, 4.0; beet pulp powder, 10.0 and medium pH was adjusted to pH 6.0. Spores of fungi were transferred from surface of the actively growing slants of (PDA) medium to 250 ml conical flasks each containing 50 ml of malt medium. After incubation on a rotary shaker (120 rpm) at 29 + 1oC for 48 h, the grown culture was employed as inocula for experimental 1000 ml conical flasks containing 100 ml (BPPM) at rate of 5 % (V/V) inoculum size.
Culture Conditions Affecting Pectinase Production
Static cultures were used for studying pectinase production by Penicillium chrysogenum under variable condition including substrate (pomegranate peel) concentration effect, incubation period, initial pH and nitrogen source. The general procedure included use of triplicate of 1000 ml conical flasks each containing 100 ml of pomegranate peel powder medium ( PPPM).
Effect of different substrate sources includes pea pods, pomegranate peel, sugar beet pulp and orange peel on pectinase production by Penicillium chrysogenum was investigated.
Effect of substrate (pomegranate peel) concentration ranged from 2.5 % to 20% (W/V) on pectinase production by Penicillium chrysogenum was investigated. The fermented pomegranate peel for each flask was mixed with 100ml of 0.02M acetate buffer (pH 5.0) by shaking in a rotary shaker (120 rpm) for one hour at room temperature to extract the enzyme and the extracted mixture was filtered and collected for pectinase activity assay.
The influence of incubation period was studied through determination of pectinase activities after 24, 48, 72, 96, 120, 144 and 168h. Effect of the initial pH of growth medium was studied through adjusting the initial pH values in a range between 3 and 8 using either 0.1N NaOH or HCl.
Effect of nitrogen source included the use of two inorganic salts (ammonium sulphate, and ammonium chloride; and three organic sources (Peptone, yeast extract and urea) were studied. Various nitrogen sources were used separately at an equivalent concentration of 0.33 g N/l media as recommended by Murad, et al. These
nitrogen sources replaced the original nitrogen source present in the PPPM. The level of a parameter optimized in an experiment was maintained in the subsequent studies.
Enzymes Sources
The produced enzyme: Laboratory produced fibrolytic enzymes from Penicillium chrysogenum. Each gram contains 200 units of pectinase and 98.82 unit of cellulase. It worth to mention that, the pectinase units of each gram of the produced enzyme was adjusted to be equivalent to that found in the commercial enzyme product (SMIZYME®). SMIZYME: A commercial enzymes source from AGRI-VET Company, Egypt. Each gram contains 200 unit of pectinase, 100 unit of cellulase, 2500 unit of α- amylase, 10000 unit of protease, 5000 unit of xylanase, 300 unit of mannanase, and 3000 unit of beta-glucannase. Enzymes assay: The pectinase and carboxymethyl- cellulase activities (CMC) for resultant crude enzyme extract (the produced enzyme) and commercial enzyme source (SMIZYME®) were determined according to Buga, et al. [13] Mandels, et al. [14] respectively. One unit of pectinase activity was defined as the amount of enzyme that produced one μmole of D-galacturonic acid per minute at 40°C and pH 5.0 [15], while one cellulase unit was defined as the amount of enzyme that liberates reducing sugar at the rate of one µmol/ ml/min under assay condition [16]. In vitro trial of the tested rations: Batch fermentation culture experiment was conducted according to Ismail, et al. [17] to evaluate impact of the produced and commercial fibrolytic enzymes sources addition to ruminant’s diets on rumen fermentation characteristics. A total mixed ration consisted of of 50% concentrates feed mixture, 20% Egyptian clover, 20 % Sugar beet pulp and 10% dried orange pulp was used as a substrate. The concentrate feed mixture consisted of 55% corn, 20 % soybean meal, 21.5 % wheat bran, 1.5 % limestone, 0.5 % di calcium phosphate, 0.2 % yeast, 0.3 % bicarbonate, 0.5 % premix and 0.5 % NaCl. For obtaining of the rumen microorganisms (inoculum), rumen fluid was collected from rumen of slaughtered rams fed clover hay ration. The produced and the commercial fibrolytic enzymes were added to control ration at the following levels: 0, 1, 2, 3, 4 and 5g/kg on DM basis. Each treatment was tested in 3 replicates accompanied by 3 blank vessels (no substrate). The tested rations (400 mg) were added separately to the 125 ml incubation vessels. Each vessel was filled with 40 ml of mixture of 1:3 (v/v) rumen fluids: buffer solution. All vessels were sealed and incubated at 39° C for 24 h. After 24 h of incubation, all vessels were filtered in fiber filter bags 25 micron porosity (ANKOM- USA). The residues in the bags were dried at 70° C in oven for 48 h to analyse dry matter (DM), neutral detergent fiber (NDF) and acid detergent fiber (ADF) digestibility. Rumen fluid pH was measured using (pH-meter). Overall volume of the produced gases was determined using Hohenheim Syringes (100 ml) as described by Navarro- Villa, et al. [18]. Quantitative analysis of ammonia concentration was carried out by a modified Nessler’s method [19]. The short chain fatty acids (SCFA) concentration was calculated according to equation of Makkar, et al. [20].
SCFA (mmol) = 0.0222 Gas–0.00425 Where, Gas: is gas production at 24 hours incubation (ml/200 mg DM).
Statistical Analysis
Statistical analyses were conducted by the general linear model procedure adapted by IBM Corp [21]. Released according to the following model:
Y =µ+T +e ij i ij Where Yij, is the dependent variable, µ is the overall mean, Ti, is the effect of treatment and eij, is the residual error. Duncan’s multiple test Duncan, et al. [22] was carried out to separate among means.
Results and Discussion
Impact of Fungal Cultures on Pectinase Production
The capability of Fusarium avenaceum, Trichoderma viride, Asperigillus flavus NRRL 5522, Cephalosporium acremonium, Fusarium oxysporum, Aspergillus niger, Asperigillus fugimatus, Penicillium chrysogenum and Aspergillus terreus on pectinase production on beet pulp powder medium (BPPM) was shown in Table 1. Penicillium chrysogenum gave significantly (P<0.05) the highest pectinase activity (10.87 µmole/ml/min) followed by Aspergillus terreus (9.37 µmole/ml/min) while the other fungal cultures gave low activity especially Fusarium avenaceum which gave the lowest pectinase activity (8.09 µmole/ml/min). Pectinases produce by many of microorganisms including fungi, yeasts and bacteria [23, 24, 25]. However, almost all the commercial preparations of pectinases are produced from fungal sources [26, 27]. Pectinase production by filamentous fungi varies according to the type of strain, cultivation conditions and the growth medium composition [1]. On
- the light of these results, Penicillium chrysogenum was chosen for further studies for pectinase production.
Table 1: Effect of fungal cultures on the produced pectinase activity (µmole/ml/min).
| Fungal cultures | Pectinase activity (µmole/ml/min) | ± SE | ||||||
|---|---|---|---|---|---|---|---|---|
| Trichoderma viride | 8.24d | 0.05 | ||||||
| Asperigillus flavus NRRL 5522 | 8.54cd | 0.08 | ||||||
| Aspergillus niger | 8.77c | 0.28 | ||||||
| Fusarium oxysporum | 8.86c | 0.13 | ||||||
| Aspergillus terreus | 9.37b | 0.08 | ||||||
| Penicillium chrysogenum | 10.87a | 0.12 | ||||||
| Fusarium avenaceum | 8.09d | 0.15 | ||||||
| Cephalosporium acremonium | 8.19d | 0.09 | ||||||
| Asperigillus fugimatus | 8.12d | 0.06 |
Table 2: Effect of different carbon sources, pH value, concentration of pomegranate peel, incubation period and nitrogen sources
Average in the same column having different superscripts are differ significantly (P<0.05) for a, b, c and d. Table 2: Effect of different carbon sources, pH value, concentration of pomegranate peel, incubation period and nitrogen sources on produced pectinase enzyme (µmole/ml/min) from Penicillium chrysogenum.
| Different carbon sources | ± SE | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pea pods | pomegranate peel | Sugar beet pulp | Orange peel | 0.27 | ||||||||
| 8.79c | 10.53a | 8.29c | 9.79b | |||||||||
| pH value | 0.36 | |||||||||||
| 3 | 4 | 5 | 6 | 7 | 8 | |||||||
| 27.01b | 28.44a | 26.18c | 25.77c | 25.72c | 24.7d | |||||||
| Concentration of pomegranate peel (%) | 1.002 | |||||||||||
| 2.5 | 5 | 7.5 | 10 | 12.5 | 15 | 17.5 | 20 | |||||
| 15.49e | 17e | 20.9d | 22.33d | 25.4c | 29.53a | 27.57ab | 26.36bc | |||||
| incubation period (day) | 0.442 | |||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||||||
| 26.6d | 29.53b | 31.55a | 29.57b | 29.18bc | 28.52c | 26.94d | ||||||
| Nitrogen sources | 0.47 | |||||||||||
| (NH4) SO 2 4 | NH Cl 4 | Yeast extract | Peptone | Urea | ||||||||
| 29.82c | 31.71b | 33.97a | 31.21bc | 31.82b |
Average in the same row having different superscripts are differ significantly (P<0.05) for a, b, c, d and e.
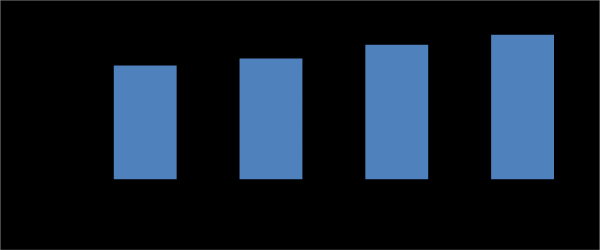
µmole/ml/min) was obtained at 2.5% of pomegranate peel powder concentration of Penicillium chrysogenum growth medium. This result is in line with that obtained by Castilho, et al. [31] who found that pectinase formation by A. niger decreases at high moisture levels. Moisture content is a critical factor on enzymes production processes because this variable has influence on growth and biosynthesis and secretion of different metabolites [32, 33]. Higher moisture levels (as 2.5% pomegranate peel powder concentration can cause a reduction in enzyme yield due to steric hindrance of the growth of the producer strain by reduction in porosity (interparticle spaces) of the solid matrix, thus interfering oxygen transfer [34]. On the other hand, Mamma, et al. [35] reported that lower moisture content causes reduction in solubility of nutrients of the substrate, low degree of swelling and high water tension. In addition, Acuna Arguelles, et al. [36] reported that, in media with low water-availability fungi suffer modifications in their cell membrane, leading to transport limitations and affecting microbial metabolism. This may be the reason for reduction of pectinase enzyme activity at 17.5 and 20% of pomegranate peel powder concentration of fungal growth medium. Based on these data, pomegranate peel powder concentration at 15% (w/v) was chosen for further studies.
Effect of Different Carbon Sources on Pectinase Production
It is generally agreed that the optimum medium for the enhanced production of pectinase is that containing pectic materials as an inducer [28, 29, 30]. The selection of a suitable substrate for the fermentation process is a critical factor. As shown in Table 2 and Figure 1 Pomegranate peel as pectolytic substrate increased significantly (P<0.05) pectinase activity by Penicillium chrysogenum reached (10.53 µmole/ml/min), while sugar beet pulp gave the lowest pectinase activity (8.29 µmole/ml/min). From the previous data, pomegranate peel selected as a sole carbon source for conducting further studies on pectinase production by Penicillium chrysogenum.
Effect of Carbon Source Concentration on Pectinase Production
Data illustrated in Table 2 and Figure 2 showed effect of different concentration of pomegranate peel powder ranged from 2.5% to 20% (W/V) on pectinase production by Penicillium chrysogenum. Maximum pectinase activity (P<0.05) reached (29.53 µmole/ml/min) was obtained at 15% (w/v) pomegranate peel powder concentration, while the minimum activity reached (15.49
Effect of Initial pH of Fungal Growth Medium on Pectinase Production
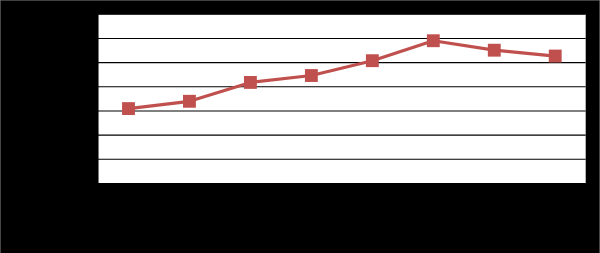
As shown in Table 2 and Figure 3 initial pH of the medium has profound effect on pectinase production. Pectinase production by Penicillium chrysogenum grown on deferent pH values of pomegranate peel powder showed its highest values (P<0.05) reached (28.44 µmole/ml/min), at pH 4.0. According to Dalagnol, et al.
[37] extreme pH values may change the protein structure, leading to a partial or irreversible denaturation, resulting in a loss of activity. Moreover when the pH level increased, the enzyme production decreased. The initial pH of the medium has a great effect on the growth of the organism, permeability membrane, as well as on the biosynthesis and stability of the enzymes [38, 39]. Optimum production of pectic enzymes from many molds within the acidic pH range has been reported by previous studies [37, 40, 41, 42]. While, Debing, et al. [43] found that the pH 6.5 was the optimal pH for pectinase production from A. niger by solid state fermentation. Also, Rasheedha, et al. [44] found that P. chrysogenum exhibited maximum polygalacturonase production at initial pH of 6.5. Based on the obtained results, the initial pH of the medium was adjusted to pH 4.0 in subsequent experiments.
Effect of Incubation Period on Pectinase Production
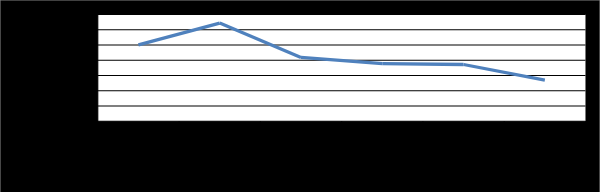
Production of pectinase was monitored for a period of seven days as shown in Table 2 and Figure 4. The highest pectinase activity (P<0.05) reached (31.55 µmole/ml/min) was recorded after three days of incubation with Penicillium chrysogenum. In addition, when the incubation period increased, the enzyme production decreased. The time of incubation depends on the growth rate of the microorganism and its enzyme production pattern. Ghildyal, et al. [45] investigated that, maximum production of pectic enzyme from different molds varies from 1 to 6 days. Castilho, et al_. [31] reported that, highest pectinase activity was obtained by _A. niger, for fermentation time 22 h while, Rangarajan, et al. [46] found that, pectinase show maximum activity after 40 h of fermentation by A. niger grow on orange peel. In addition, Leda, et al. [47] reported that, the highest polygalacturonase activities were obtained by A. niger after 70 h of fermentation period. Moreover, Sarvamangala, et al. [48] observed a gradual increase in the production of pectinase from deseeded sunflower head by A. niger after 72 h of fermentation period in submerged and up to 96 h in solid-state conditions. From the previous data, 3 days incubation period was selected for conducting further studies on modified pomegranate peel powder medium.
![Figure 4: The highest pectinase activity (P<0.05) reached (31.55 µmole/ml/min) was recorded after three days of incubation with _Penicillium chrysogenum_**_._** In addition, when the incubation period increased, the enzyme production decreased. The time of incubation depends on the growth rate of the microorganism and its enzyme production pattern. Ghildyal, et al. [45] investigated that, maximum production of pectic enzyme from different molds varies from 1 to 6 days. Castilho, et al_._ [31] reported that, highest pectinase activity was obtained by _A. niger_, for fermentation time 22 h while, Rangarajan, et al. [46] found that, pectinase show maximum activity after 40 h of fermentation by _A. niger_ grow on orange peel. In addition**,** Leda, et al. [47] reported that, the highest polygalacturonase activities were obtained by _A. niger_ after 70 h of fermentation period. Moreover, Sarvamangala, et al. [48] observed a gradual increase in the production of pectinase from deseeded sunflower head by _A. niger_ after 72 h of fermentation period in submerged and up to 96 h in solid-state conditions. From the previous data, 3 days incubation period was selected for conducting further studies on modified pomegranate peel powder medium.](/fulltextimages/4879/fig_4.png)
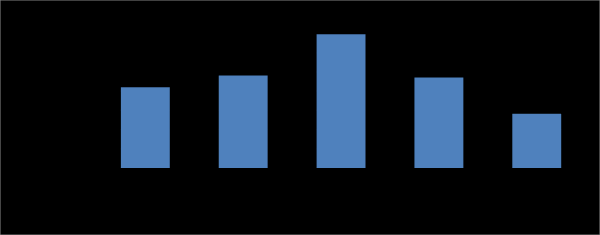
Effect of Nitrogen Sources on Pectinase Production
(3.78 U/ml) on beet pulp powder media. Also, Aguilar, et al. [49] who reported that yeast extract the best inducer of pectinases by Aspergillus sp. Moreover, Kashyap, et al. [50] found that, yeast extract, peptone and ammonium chloride were found to enhance pectinase production up to 24% and addition of glycine, urea and ammonium nitrate inhibited pectinase production. In addition, Vivek, et al. [51] found that, organic nitrogen sources showed higher endo, exo pectinases activities than inorganic nitrogen sources. Also the increasing trend in the enzymes activity with the increase in nitrogen source content was observed in the case of organic nitrogen sources while decreasing trend observed for inorganic nitrogen sources.
- Figure 5: Effect of nitrogen sources on pectinase production. the tested rations compared to control one. Increasing the produced enzyme and SMIZYME® supplementation levels up to 3g/kg DM gave the highest values of in vitro DMD degradation.
- In Vitro DMD, NDF and ADF Degradation of the
- Tested Rations
- Results of Table 3 showed that, all the produced enzyme and SMIZYME® supplementation levels increased
- (P<0.05) the in vitro DMD, NDF and ADF degradation for
Table 4: Fibrolytic enzymes effect on DMD, NDF and ADF degradation of the tested rations (in vitro).
| Enzymes Source | Enzyme level g/Kg DM | IVDMD% | ±SE | IVNDF % | ±SE | IVADF% | ±SE | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 | 45.16e | 0.58 | 26.74d | 0.6 | 20.22f | 0.19 | ||||||||||||||||
| The produced enzyme | 1 | 47.45d | 0.55 | 30.39cd | 2.68 | 23.19d | 0.97 | ||||||||||||||||
| 2 | 48.8bc | 0.03 | 36.22a | 0.38 | 28.72ab | 0.61 | |||||||||||||||||
| 3 | 51.68a | 0.56 | 37.39a | 0.32 | 29.46ab | 0.15 | |||||||||||||||||
| 4 | 49.76b | 0.59 | 35.51a | 0.43 | 28.00bc | 0.68 | |||||||||||||||||
| 5 | 49.10bc | 0.09 | 34.45ab | 0.37 | 26.97c | 0.15 | |||||||||||||||||
| SMIZYME® | 1 | 48.22cd | 0.3 | 28.68cd | 2.07 | 20.84ef | 0.13 | ||||||||||||||||
| 2 | 48.31cd | 0.24 | 31.40bc | 1.45 | 22.11de | 0.01 | |||||||||||||||||
| 3 | 52.19a | 0.35 | 34.42ab | 0.08 | 29.90a | 0.09 | |||||||||||||||||
| 4 | 51.59a | 0.12 | 38.19a | 1.03 | 29.71a | 0.29 | |||||||||||||||||
| 5 | 49.95b | 0.2 | 38.12a | 0.04 | 29.06ab | 0.53 |
Table 5: Fibrolytic enzymes effect on DMD, NDF and ADF degradation of the tested rations (in vitro).
Average in the same column having different superscripts are differ significantly (P<0.05) for a, b, c d, e and f.
Colombatto, et al. [52] mentioned that, using commercial enzyme product supplemented to alfalfa stems at six levels: 0, 0.51, 1.02, 2.55, 5.1 and 25.5 g/kg DM, increased in vitro DM, NDF, ADF, and hemicellulose degradation compared to control. Also Pinos, et al. [53] showed that, enzymes improved ruminal disappearance rates of DM and NDF in situ of total mixed ration (TMR) with three different forage: concentrate ratios (400:600, 500:500, 600:400 g/g) and two (0 or 2 g) levels of enzymes/kg DM. Moreover Azzaz, et al_. [54] investigated that Asperozym and Bacillozym® addition to banana wastes increased IVDMD and IVOMD significantly compared with the untreated banana wastes (Control). From the result of the current study, increasing of DM, NDF, ADF, and hemicellulose degradation (_in vitro) because of supplemented rations with fibrolytic enzymes may be increased numbers of total bacteria and Fibrobacter succinogens in the incubation medium with improving in vitro fermentation as mentioned by Mao, et al. [55]. Moreover Kholif, et al. [56] found that, the treatment with commercial cellulolytic enzymes (Veta- Zyme Plus®) increased DM digestibility (in vitro) further Aboul Fotouh, et al. [4] reported that, addition lactating goats with Asperozym and Phytabex plus® at levels up to 1000 unit of cellulolytic enzymes/kg DM increase (P≤0.05) values of IVDMD compared to control.
In Vitro PH Value, TGP, NH3-N and SCFA of the Tested Rations
Data of Table 4 showed that, all the produced enzyme and SMIZYME® supplemented levels decreased (P<0.05) in vitro pH value and increased (P<0.05) total gas production (TGP) compared to control ration. The highest values (P<0.05) of in vitro pH (6.7) was recorded by control ration, while the ration supplemented with SMIZYME® recorded the lowest values of in vitro pH (5.69) at levels up to 3g/kg DM .On the other hand, the highest values of in vitro TGP was recorded by the produced enzyme ration reached (295.26) at levels up to 4g enzyme /kg DM., while control ration recorded the lowest values of in vitro TGP reached (283.66).
| Enzymes Source | Enzyme level g/Kg DM | pH value | ±SE | TGP/1g | ±SE | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 | 6.70a | 0.14 | 283.66d | 0.6 | ||||||||||||
| The produced enzyme | 1 | 6.47b | 0.03 | 290.05bc | 0.93 | ||||||||||||
| 2 | 6.37bc | 0.05 | 291.62b | 0.44 | |||||||||||||
| 3 | 6.15d | 0.06 | 294.99a | 0.08 | |||||||||||||
| 4 | 6.24cd | 0.01 | 295.26a | 0.11 | |||||||||||||
| 5 | 6.38bc | 0.06 | 294.45a | 1.01 | |||||||||||||
| SMIZYME® | 1 | 6.13d | 0.12 | 291.37b | 1.32 | ||||||||||||
| 2 | 5.89e | 0.01 | 295.02a | 0.73 | |||||||||||||
| 3 | 5.69e | 0.1 | 294.44a | 1.04 | |||||||||||||
| 4 | 5.74e | 0.07 | 288.07c | 0.18 | |||||||||||||
| 5 | 5.87e | 0.01 | 287.66c | 1.3 |
Table 6: Fibrolytic enzymes effect on pH value andTGP of the tested rations (in vitro).
Average in the same column having different superscripts are differ significantly (P<0.05) for a, b, c, d and e.
Moreover data of Table 5 showed that, all the produced enzyme and SMIZYME® supplementation levels increased (P<0.05) the in vitro short chain fatty acid (SCFA). Control ration recorded the lowest value of in vitro short chain fatty acid (SCFA) being (1.255 mmol/dl), while the highest value in vitro SCFA, being (1.310
vitro fermentation by increasing substrate fiber degradation, Volatile Fatty Acids (VFA) production, and ruminal microbial growth. Moreover Elghandour, et al. [60] obtained decreased ruminal pH values and increased in vitro SCFA production when incubated 4 fibrous feeds, including corn stover, with different levels of exogenous fibrolytic enzyme. The effectiveness of enzymes depends upon substrate, enzyme specificity and enzyme dose causing variable responses with different enzyme preparations and doses [61]. Increased TGP indicated the increased fermentable material with enzyme addition [62]. Also Vallejo, et al. [63] investigate that, addition of cellulase and xylanase linearly decreased (P≤0.05) pH compared to control, this may be due to greater enzymatic hydrolysis of feeds into readily fermentable substrates that depress pH when fermented. Further Handique, et al. [64] reported that, the in vitro study by using complete feed containing bajra straw supplemented with exogenous fibrolytic enzyme increase in total volatile fatty acid and an unaltered rumen pH, total nitrogen and ammonia N compared to control. These support the hypothesis that a suitable enzyme dose could improve fermentation efficiency [65, 66].
| Enzymes Source | Enzyme level g/Kg DM | NH3-N µmol/ml | ±SE | SCFA (mmol/dl) | ±SE | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 0 | 2.78 | 0.09 | 1.255d | 0.005 | ||||||||||||
| The produced enzyme | 1 | 2.75 | 0.2 | 1.284c | 0.005 | ||||||||||||
| 2 | 2.59 | 0.12 | 1.291bc | 0.001 | |||||||||||||
| 3 | 2.4 | 0.02 | 1.306a | 0.005 | |||||||||||||
| 4 | 2.66 | 0.07 | 1.307a | 0.003 | |||||||||||||
| 5 | 2.56 | 0.23 | 1.303ab | 0.004 | |||||||||||||
| SMIZYME® | 1 | 2.03 | 0.21 | 1.289bc | 0.01 | ||||||||||||
| 2 | 2.6 | 0.32 | 1.306ab | 0.006 | |||||||||||||
| 3 | 2.29 | 0.51 | 1.303ab | 0.005 | |||||||||||||
| 4 | 2.33 | 0.14 | 1.275c | 0.004 | |||||||||||||
| 5 | 2.77 | 0.08 | 1.273c | 0.0039 |
Table 7: Fibrolytic enzymes effect on NH3-N and SCFA of the tested rations (in vitro).
Conclusion
Fungal pectinase enzyme was locally produced (the produced enzyme) under the optimum conditions. This would contribute for reducing the cost of importation. The evaluated fibrolytic enzymes (the produced enzyme and SMIZYME®) were increased DM, NDF, ADF, TGP and SCFA (in vitro).
References
-
Murad HA, Azzaz HH (2011) Microbial pectinases and ruminant nutrition. Research journal of microbiology 6(3): 246-269.
-
Ruiz HA, Rodrigue Jasso R, Rodriguez R, Contreras Esquivel JC, Aguilar CN (2012) Pectinase production from lemon peel pomace as support and carbon source in solid-state fermentation column-tray bioreactor. Biochem Eng J 65: 90-95.
-
El Enshasy HA, Elsayed EA, Suhaimi N, Abd Malek R, Esawy M (2018) Bioprocess optimization for pectinase production using _Aspergillus niger_ in a submerged cultivation system. BMC Biotechnology 18: 71.
-
Aboul Fotouh GE, El Garhy GM, Azzaz HH, Abd El Mola AM, Mousa G A (2016) Fungal cellulase production optimization and its utilization in goat’s rations degradation. Asian Journal of Animal and Veterinary Advances Journal 11(12): 824-831.
-
Khattab MS, Azzaz HH, Abd El Tawab AM, Murad H A ( 2019) Production optimization of fungal cellulase and its impact on ruminal degradability and fermentation of diet. Int J Dairy Sci 14(2): 61-68.
-
Azzaz HH, Murad HA, Kholif AM, Morsy TA, Mansour AM, et al. (2013a) Pectinase production optimization and its application in panama fiber degradation. Egyptian J Nutrition and Feeds 16(2): 117-125.
-
Murad HA, Azzaz HH (2010) Cellulase and dairy animal feeding. Biotechnology 9(3): 238-256.
-
Azzaz HH, Murad HA, Kholif AM, Hanfy MA, Abdel Gawad MH (2012) Optimization of Culture Conditions Affecting Fungal Cellulase Production. Research Journal of Microbiology 7(1): 23-31.
-
Azzaz HH, Morsy TA, Murad HA (2016) Microbial Feed Supplements for Ruminant’s Performance Enhancement. Asian Journal of Agricultural Research 10(1): 1-14.
-
Azzaz HH, Murad HA, Kholif AM, Morsy TA, Mansour AM, et al. (2013) Increasing nutrients bioavailability by using fibrolytic enzymes in dairy buffaloes feeding. Journal of Biological Sciences 13(4): 234-241.
-
Azzaz HH, Eman SA Farahat, Ebeid HM (2017) Effect of partial replacement of corn grains by date seeds on rahmani ram’s nutrients digestibility and Nubian goat’s milk production. Int J Dairy Sci 12(4): 266-274.
-
Kholif AE, Kassab AY, Azzaz HH, Matloup OH, Hamdon HA, et al. (2018) Essential oils blend with a newly developed enzyme cocktail works synergistically to enhance feed utilization and milk production of Farafra ewes in the subtropics. Small Ruminant Research 161: 43-50.
-
Buga ML, Ibrahim S, Nok AJ (2010) Partially purified polygalacturonase from _Aspergillus niger_ (SA6). African Journal of Biotechnology 9(52): 8944-8954.
-
Mandels M, Hontz L, Nystrom J (1974) Enzymatic hydrolysis of waste cellulose. Biotech Bioeng 16: 1471.
-
Soares MN, Silva R, Gomes E (1999) Screening of Bacterial Strains for Pectinolytic Activity Characterization of the pgase Produced by _Bacillus_ _species_. Rev Microbiol 30: 229-303.
-
Miller GL (1972) Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Biotechnol Bioeng Symp 5: 193-219.
-
Ismail SA, Abdel Fattah AM, Emran MA, Azzaz HH, El Gamal MS, et al. (2018) Effect of Partial Substitution of Ration’s Soybean Meal by Biologically Treated Feathers on Rumen Fermentation Characteristics (_in_ _vitro_). Pakistan Journal of Biological Sciences 21: 110- 118.
-
Navarro Villa A, M O’brien, Lopez S, Boland TM, O’kiely P (2011) Modifications of a gas production technique for assessing in vitro rumen methane production from feedstuffs. Anim Feed Sci Technol 166-167: 163-174.
-
Szczechowiak J, Szumacher Strabel M, El Sherbiny ME, Pers Kamczyc P, Pawlak, et al. (2016) Rumen fermentation, methane concentration and fatty acid proportion in the rumen and milk of dairy cows fed condensed tannin and/or fish-soybean oils blend. Anim Feed Sci Technol 216: 93-107.
-
Makkar HP (2005) _In vitro_ gas methods for evaluation of feeds containing phytochemicals. Anim Feed Sci Technol 123: 291-302.
-
IBM Corp Released (2011) IBM SPSS Statistics for Windows. Version 20.0. Armonk, NY: IBM Corp.
-
Duncan DB (1955) Multiple range and multiple F-test. Biometrics 11: 1-42**.**
-
Cao J, Zheng L, Chen S (1992) Screening of pectinase producer from alkalophilic bacteria and study on its potential application in degumming of ramie. Enzyme and Microbial Technology 14(12): 1013-1016.
-
Blanco P, Sieiro C, Villa TG (1999) Production of pectic enzymes in yeast. FEMS Microbiology Letters 175: 1-9.
-
Huang LK, Mahoney RR (1999) Purification and characterization of an endo-polygalacturonase from Verticillum alboatrum. Journal of Applied Microbiology 86(1): 145-146.
-
Godfrey T, West S (1996) Industrial Enzymology. 2nd (Edn.), London: Macmillan Press.
-
Uhlig H (1998) Industrial enzymes and their applications, New York: John Wiley & Sons, Inc pp: 435.
-
Solis Pereira S, Favela Torres E, Viniegra Gonzales G, Gutierrz-Rojas M (1993) Effect of different carbons sources on the synthesis of pectinase by Aspergillus niger in submerged and solid state fermentation. Appl Microbiol Biotechnol 39: 36-41.
-
Hang YD, Woodanms EE (1994) Production of fungal polygalacturonase from apple pomace. Lebensm. Wiss U Technol 27: 194-196.
-
Naidu GSN, Pandam T (1998) Production of pectolytic enzymes a review Process Eng 19(5): 355-361.
-
Castilho LR, Ricardo AM, Tito LMA (2000) Production and extraction of pectinases obtained by solid state fermentation of agroindustrial residues with _Aspergillus niger_. Bioresource Technol 71: 45-50.
-
Krishna C, Chandrasekaran M (1996) Banana waste as substrate for aamylase production by _Bacillus_ _subtilis_ (CBTK 106) under solid state fermentation. Appl Microbiol Biotechnol 46(2): 106-111.
-
Ellaiah PK, Adinarayana Y, Bhavani P, Padmaja B, Srinivasulu (2002) Optimization of process parameters for glucoamylase production under solid state fermentation by a newly isolated Aspergillus species. Process Biochem 38(4): 615-620.
-
Lonsane BK, Ghildyal NP, Budiatman S, Ramakrishna SV (1985) Engineering aspects of solid state fermentation. Enzyme Microbiol Technol 7(6): 258- 265.
-
Mamma D, Kourtoglou E, Christakopoulos P (2008) Fungal multienzyme production on industrial by- products of the citrus-processing industry. Bioresource Technolo 99(7): 2373-2383.
-
Acuna Arguelles ME, Guti_errez Rojas M, Viniegra Gonz_alez G, Favela-Torres E (1995) Production and properties of three pectinolytic activities produced by _A. niger_ in submerged and solid state fermentations. Appl Microbio, Biotechnol 43(5): 808-814.
-
Dalagnola LMG, Silveiraa VCC, Silvaa HBD, Manfroib V, Rodrigues RC (2017) Improvement of pectinase, xylanase and cellulase activities by ultrasound: Effects on enzymes and substrates, kinetics and thermodynamic parameters. Process Biochemistry 61: 80-87.
-
Shoichi T, Xoighi K, Hiroshi S (1985) Cellulase production by _P. purpurogenum_. J Ferment Technol 62: 127.
-
Poorna AC, Prema P (2006) Production of cellulase free endoxylanase from novel alkalophilic thermotolerent _Bacillus pumillus_ by solid state fermentation and its application in wastepaper recycling. Bio Reso Tech 98(3): 485-490.
-
Zetelaki Horvath K (1980) Factors affecting pectinase activity. Acta Alimentaria 10: 371-378.
-
Shin I, Dowmez S, Kilic O (1983) Study on pectolytic enzyme production from some agricultural wastes by fungi. Chemical Microbiologie Technologie der Lebensm 8: 87-90.
-
Ma X , Wang D , Yin M , Lucente J, Wang W, et al. (2017) Characteristics of pectinase treated with ultrasound both during and after the immobilization process, Ultrason Sonochem 36: 1-10.
-
Debing J, Peizun L, Stagnitti F, Xianzhe X, Li L (2006) Pectinase production by solid fermentation from _Aspergillus niger_ by a new prescription experiment. Ectoxicology and Environ 64(2): 244-250.
-
Rasheedha AB, Kalpana MD, Gnanaprabhal GR, Pradeep BV, Palaniswamy M (2010) Production and characterization of pectinase enzyme from Penicillium chrysogenum 3(4): 377-381.
-
Ghildyal NP, Ramakrishna SV, Nirmala P, Devi BK, Asthana HA (1981) Large scale production of pectolytic enzyme by solid state fermentation. Journal of Food Science and Technology 18(6): 243-251.
-
Rangarajan V, Rajasekharan M, Ravichandran R, Sriganesh K, Vaitheeswaran V (2010) Pectinase Production from Orange Peel Extract and Dried Orange Peel Solid as Substrates Using A_spergillus_ _niger._ International Journal of Biotechnology and Biochemistry 6: 445-453.
-
Leda RC, Ricardo AM, Tito LM (2000) Production and extraction of pectinases obtained by solid state fermentation of agroindustrial residues with _Aspergillus niger_. Bioresource Technology 71: 45-50.
-
Sarvamangala RP, Dayanand A (2006) Production of pectinase from deseeded sunflower head by _Aspergillus niger_ in submerged and solid-state conditions. Bioresource Technology 97(16): 2054- 2058.
-
Aguilar G, Trejo B, Garcia J Huitron G (1991) Influence of pH on endo and exo- pectinase production by _Aspergillus_ species CH-Y-1043. Can J Microbiol 37: 912-917.
-
Kashyap DR, Soni SK, Tewari R (2003) Enhanced production of pectinase by _Bacillus_ sp. DT7 using solid state fermentation. Bioresou. Technol 88(3): 251-254.
-
Vivek R, Rajasekharan M, Ravichandran R, Sriganesh K, Vaitheeswaran V (2010) Pectinase production from orange peel extract and dried orange peel solid as substrates using A_spergillus niger_. International Journal of Biotechnology and Biochemistry 6: 445- 453.
-
Colombatto D, Mould FL, Bhat MK, Owen E (2007) Influence of exogenous fibrolytic enzyme level and incubation pH on the _in vitro_ ruminal fermentation of alfalfa stems. Anim Feed Sci Technol 137(1-2): 150- 162.
-
Pinos RJ, Moreno R, Gonzalez SS, Robinson PH, Mendoza G Alvarez G (2008) Effects of exogenous fibrolytic enzymes on ruminal fermentation and digestibility of total mixed rations fed to lambs. Anim Feed Sci Technol 142(3-4): 201-219.
-
Azzaz HH , Murad HA, Kholif AM, Hanafy MA, Abdel Gawad M H (2012b) Utilization of cellulolytic enzymes to improve the nutritive value of banana wastes and performance of lactating goats. Asian J Anim Vet Adv 7(8): 664-673.
-
Mao HL, Wu CH, Wang JK, Liu JX (2013) Synergistic effect of cellulase and xylanase on in vitro rumen fermentation and microbial population with rice straw as substrate. Animal Nutrition and Feed Technology 13: 477-487.
-
Kholif AM, Farahat ESA, Hanafy MA, Kholif SM, El Sayed RR (2015) Utilization of cellulolytic enzymes to improve the nutritive value of date kernels and the investigation of the impact of adding these enzymes to lactating goat’s diets on rumen fermentation and nutrients digestibility. Asian J Anim Sci 9: 441-447.
-
Nsereko VL, Beauchemin KA, Morgavi DP, Rode LM , Furtado AF, et al. (2002) Effect of a fibrolytic enzyme preparation from Trichoderma longibrachiatum on the rumen microbial population of dairy cows. Can J Microbiol 48(1): 14-20.
-
Mohamed AM, El Saidy BE, Ibrahim K, Tejido ML, Carro MD (2005) Effect of exogenous enzymes on _in_ _vitro_ ruminal fermentation of a high-forage diet and productive response of lactating ewes. Egyptian J Nutr feeds 8: 591-602.
-
Giraldo LA, Tejido ML, Ranilla MJ, Carro MD (2008) Effects of exogenous fibrolytic enzymes on _in vitro_ ruminal fermentation of substrates with different forage: concentrate ratios. Anim Feed Sci Technol 141: 306-325.
-
Elghandour MMY, Salem AZM, Gonzalez Ronquillo M, Brquez JL, Gado HM, et al. (2013) Effects of exogenous enzymes on in vitro gas production kinetics and ruminal fermentation of four fibrous feeds. Animal Feed Science and Technology 179(1-4): 46-53.
-
Salem AZM, Ammar H, Kholif AE, Elghandour MMY, Ortiz LB (2015) Effect of glucoamylase enzyme extract on in vitro gas production and degradability of two diets with 25% of corn or sorghum grains. Indian Journal of Animal Sciences 85: 183-188.
-
Elghandour MMY, Salem AZM, Martinez Castaneda JS, Camacho LM, Kholif AE, et al. (2015) Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. Journal of Integrative Agriculture 14(3): 526-533.
-
Vallejo LH, Salem AZM, Kholif AE, El Ghandour MMY, Fajardo RC, et al. (2016) Influence of cellulase or xylanase on the _in vitro_ rumen gas production and fermentation of corn stover. Indian Journal of Animal Sciences 86 (1): 70-74.
-
Handique B, Maurya LK, Devi YR, Pearlin VB (2018) Supplementation of exogenous fibrolytic enzyme in livestock nutrition. Journal of Entomology and Zoology Studies 6(6): 302-305.
-
Azzaz HH, Murad HA, Morsy TA (2015) Utility of Ionophores for Ruminant Animals: A Review. Asian Journal of Animal Sciences 9(6): 254-265.
-
Abd El Tawab AM, Murad HA, Khattab MS, Azzaz HH (2019) Optimizing production of tannase and in vitro evaluation on ruminal fermentation, degradability and gas production. Int J Dairy Sci 14(2): 53-60.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind