Expression and Purification of the Recombinant OmpL1 Protein for Potential Vaccine Production against Leptospirosis
Leptospirosis is a popular zoonosis caused by pathogenic strains of the Leptospira. OmpL1 is the outer membrane protein present in most pathogenic Leptospira strains and consider an important factor in the immunogenicity response. We describe here a simple procedure for generating recombinant OmpL1 protein resistant to Leptospira. OmpL1102-316 recombinant was cloned on Escherichia coli strain DH10b and expressed in strain BL21. The purification has performed by an affinity-linked Ni- Sepharose column with His-tag attached to this protein. The recombinant protein OmpL1 induced immunological memory in laboratory rats, suggesting a potential recombinant vaccine production for using in against Leptospirosis in animals.
Introduction
Leptospirosis, also known as acute jaundice, spreads from animals to human by the Leptospira pathogenic strains and is one of the most popular zoonoses in the world [1]. According the World Health Organization (WHO) and other data sources, each year between 7 and 10 million people suffers from Leptospirosis, with an estimated rate of 0.1- 1/ 100,000 in temperate climates and 10-100 / 100,000 in tropical climates such as South Asia and South East Asia [2, 3]. Leptospirosis spreads through the skin, mucous membranes or by direct or indirect contact with mucous membranes, urine, or infected organisms. In human, the disease has clinical manifestations such as latent bacterial infection, systemic toxicity syndrome, liver, kidney damage and Weil syndrome [4]. In animals, it is one of the causes of stillbirth, reduced fertility, productivity and mortality, thereby causing serious economic losses [5]. Currently, the commercial veterinary Leptospiral vaccines are large amount of inactivated vaccine from whole cells of Leptospira. However, these vaccines have few disadvantages; they give short-term immunity [6] and elicit protection only against serovar contained in preparation.
The identification of genes encoding several antigens specific to pathogenic Leptospira is the decisive step in recombinant vaccine research. Surface contact proteins, acting at the bacterial-host interface, are conserved between different serovars and considered suitable cross-protection vaccine candidates [7]. The outer membrane proteins (OMPs) are the most important and highly conserved agent in pathogenic Leptospira strains. In these, trans-membrane outer membrane protein L1 (OmpL1) is one of the most sensitive proteins for antigen-antibody immune response [8, 9]. The OmpL1 trans-membrane protein including 320
amino acids was first reported in 1993 [10]. It was shown to appear in 15 standard strains and 163 new pathogenic isolates of several Leptospira serogroups [8]. Specific immunity stimulatory abilities in the animal model are due to the OmpL1’s ability to bind to extracellular matrix (ECM) and other serum components based on laminin binding and plasma fibronectin [11]. Previous researches showed that recombinant OmpL1 protein (rOmpL1) could create protective immunity in animals [8, 10], these indicate the high potential of rOmpL1 in vaccine production. In this work, we describe the cloning and expression of a novel multi-epitope fusion protein, containing sequences of diverse previously studied OMPs of Leptospira, followed by its immunological and immune protective characterization in the hamster model of Leptospirosis.
Materials and Methods
Materials
Total DNA of five strains of Leptospira interrogans includes Leptospira interrogans serovar Bataviae (L. Bataviae), Leptospira interrogans serovar Canicola (L. Canicola), Leptospira interrogans serovar Grippotyphosa (L. Grippotyphosa), Leptospira interrogans serovar Icterohaemorrhagiae (L. Icterohaemorrhagiae) and Leptospira interrogans serovar Pomona (L. Pomona), which are currently used to produce the inactivated Leptospira vaccines in Vietnam, was extraction using GeneJET Genomic DNA Purification Kit (Thermo Scientific, USA).
The pJET1.2 / blunt vector (Thermo Fisher Scientific, USA) with size 2,974bp contains ampicillin resistance gene (Amp), capable of inserting a gene segment with size from 6bp to 10kb. The pJET1.2/ blunt vector was used to create pJET1.2-OmpL1 plasmid.
Primer Design and PCR
The OmpL1 sequence information was collected on the GeneBank/NCBI. Based on the information of OmpL1 gene with accession numbers AY622662, using Primer3 software to design a primers pair for the OmpL1 gene xTotal DNA samples were used as a template for PCR reaction, amplifying the entire OmpL1 gene segment of five Leptospira spp. Components of the reaction PCR include 10X PCR buffer (2.5µL), 2.5mM dNTP (2.5µL), 10 pmol/µL forward or reverse primer (0.5µL), 5U/µL Taq DNA polymerase (0.15µL), 200pmol/µL template (1µL), and sterile distilled water 17.85µL. PCR reaction was performed with 25 cycles: denatured at 95°C for 30s, annealed at 55°C for 30s, and extended at 72°C for 7 min. OmpL1 gene segments after purification and quality control were sequenced on ABI 3500 sequencing machine. The obtained OmpL1 gene sequences were compared analyzed by BioEdit, BLAST and MEGA 6.06 software and selected suitable gene segment for cloning into pJET1.2 vector.
Cloning OmpL1 Gene
OmpL1 gene was cloned from L.Batavie genome DNA with specifically designed primers. The PCR product was cloned to pJET1.2 vector using CloneJET PCR Cloning Kit (Thermo Fisher) [13]. Recombinant vector containing OmpL1 gene was then transformed into E. coli strain DH10b strain by heat shock method. Results of the transformation into E.coli of the recombinant vector pJET1.2-LipL21 were checked by PCR using OmpL1 gene-specific primers. At the same time, the colonies modeling positive PCR reactions were grown in liquid LB medium supplemented with 50µg/mL of Ampicillin overnight at 37°C, shaking 200rpm. Plasmid DNA extraction checked for the presence of the OmpL1 gene by cutting by restriction enzymes EcoRV and XhoI according to the manufacturer’s instructions (Thermo Fisher, USA). OmpL1 gene sequence was determined by ABI 3500 sequencing machine using pJET1.2 Forward primer. The obtained nucleotide sequence was compared with the OmpL1 gene sequence using Bio edit.
Expression of rOmpL1 Protein in E.coli
The OmpL1 gene fragment was ligated in to pET32a by T4 DNA ligase enzyme in VDNA:Vvector ratio is 2:1. Components of 20µL reaction include: 10x Buffer: 2µL, 50% PEG: 2µL, T4 ligase: 1µL, pET32a vector: 5µL and DNA fragment: 10µL. The reaction mixture was incubated overnight at 4°C in order to form pET32a-OmpL1 plasmid and transformed into E.coli BL21. The transformed E.coli BL21 was cultured in liquid LB medium containing 50µg ampicillin and grown until the optical density (OD) reached 0.6 – 0.8 at 260nm. At this time, 100µl IPTG (1mM) was added into medium and incubation continued until 4 hours later at 37°C, shaking 200rpm. The cells were collected by centrifugation and re-suspended by intermittently sonicating in 30 minutes in ice. The soluble and insoluble fractions were isolated by centrifugation at 9000rpm for 10min.
Purification of the rOmpL1 Protein
The recombinant OmpL1 protein contains His tag to facilitate purification. His tag is found only in the recombinant protein but not in the E. coli protein, makes a specific ability to adhere to the chromatographic column contained Ni2+. Recombinant OmpL1 that have specific interactions with Ni2+ will be mounted on the column, and the non-clinging or nonspecific substances are expelled. Purification process uses gel, column and AKATA prime Plus automatic chromatography system (GE Healthcare).
The soluble fraction recovered from the centrifugation step described above was applied to a column containing Ni- sepharose gel. Equilibrating the gel column with 5 times the volume of 0.02M phosphate buffer (5mM Imidazol included) and get ready for the next purification step. Protein extracts were added to the column at a rate of 0.6mL/min. Unbound proteins were cursed with supplemented 0.02M phosphate buffer with the addition of 60mM Imidazol. The protein fraction attached to the column was eluted with 0.02M phosphate buffer supplemented with 500mM Imidazol. Fractions were analyzed by 12.5% SDS-PAGE to detect the presence of rOmpL1 and determine by Bradford method at OD 595nm. Pooled proteins were dialyzed three times in phosphate buffer saline and considered to be the final production.
Production of Antiserum against rOmpL1
Five to eight-week-old mice were immunized with 10µg purified rOmpL1 and 100µg aluminum (Al(OH)3). The experiments were performed once a week in three weeks and mice’s blood was collected in week fourth. Serum was collected in supernatant by centrifugation of blood on 6000rpm at 4°C.
Western blot for the Evaluation of Antibody Response
Total protein from Leptospia were fractionated by 12.5% SDS-PAGE and electro-transferred to a PVDF membrane. Transferring membrane was performed at 60mA/gel, 4°C, for 2 hours, potential 100V. Then, the membrane was blocked with 5% skim milk-TBST for 1 hour (or overnight at 4°C). Next, incubated membrane with primary antibody (mice’s serum in previous step) mixed in blocking solution for 1.5 hours at room temperature then washed 3 times by TBST in 5 minutes. The membrane was continued to incubate with second antibodies in 5% nonfat dried milk-TBST solution (ratio 1:1000) for 2 hours, following the repetition of the TBS-T wash as described above. The second antibody is goat anti-mouse IgG peroxidase conjugate (Sigma, USA).
Results
Prediction and Selection of Vaccine Epitopes and Primer Design
Based on sequencing result, five Leptospira serovar’s OmpL1 genes divided into two groups, the first group was included 3 strains of L. Bataviae, L. Canicola, and L. Grippotyphosa with similarity about 99%, second once included strains of L. Ictero haemorrhagiae and L. Pomona also achieved similarity rates up to 98%. Using I-TASSER and TM-score tool to integrate finding common ground between two 3D structures of 5 OmpL1 proteins [14] (Figure 1), amino acid sequence from 102 to 316 residues (Table 1 & Figure 2) was selected. And PCR was used to amplify the gene fragment coding for OmpL1102-316 using DNA from L. Canicola, as template, with the forward primer (5’- CCTGCTGATATCAGATTAATCACCCTTGATAG -3’) containing EcoRV restriction endonuclease sites (underlined) and the reverse primer (5’- TTAGAGCTCGAGTTTATAACCGAATCTGTAGA -3’) containing a XhoI (underlined) restriction site.
| No. | Start | End | Peptide | Length |
|---|---|---|---|---|
| 1 | 19 | 24 | AASLSA | 6 |
| 2 | 40 | 98 | QLGGTITKDGLDAATHYGPVRSSNTCTVGPSDPACFQNPAKPTGEGNYIGVAPRKAIPA | 59 |
| 3 | 106 | 121 | DRTTGGAINARSTKGA | 16 |
| 4 | 132 | 136 | ESDFG | 5 |
| 5 | 148 | 161 | TQKISGGVTKADIA | 14 |
| 6 | 166 | 170 | VDMTW | 5 |
| 7 | 207 | 220 | SLNGSNNIKGGYDI | 14 |
| 8 | 226 | 248 | AGAVANLLSDGTDPVTTREHVRF | 23 |
| 9 | 251 | 251 | S | 1 |
| 10 | 265 | 267 | VTD | 3 |
| 11 | 285 | 300 | VGKTQSVGGATNLSPF | 16 |
| 12 | 314 | 316 | FGY | 3 |
Table 1: LipL21 Predicted epitopes.
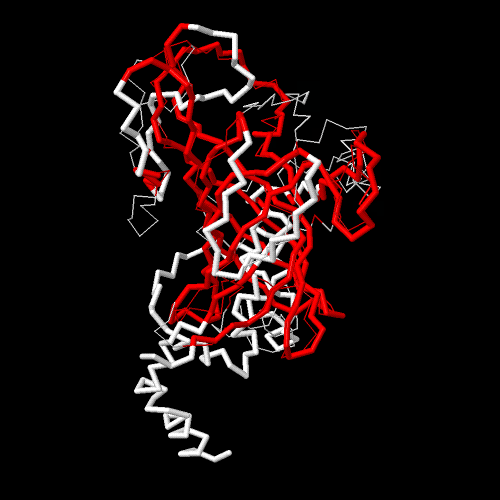
(The red is similar sequence) Figure 1: 3D construction was combined from five Leptospira serovars.
Expression and Purification of Recombinant OmpL1
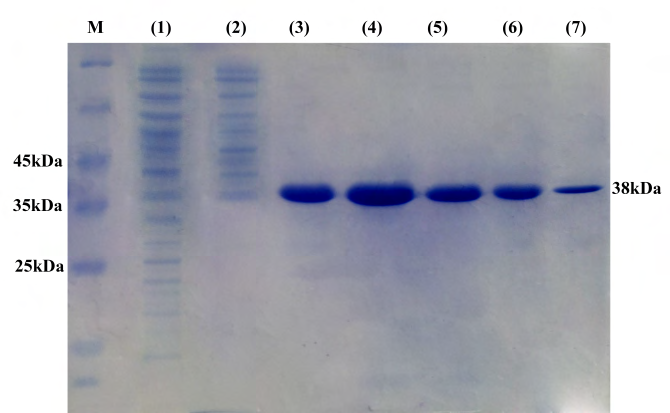
The OmpL1 gen was successfully ligated into pET32a to make pET32a-OmpL1 vector then transformed into E.coli BL21. PCRs confirmed the accuracy of recombinant plasmids extracted from five colonies after culture. Furthermore, PCR product sequences were completely similar to previous calculation (data not shown). The rOmpL1 was expressed from E.coli BL21 with pET32a-OmpL1 contained under IPTG induction. The recombinant proteins after purification showed a single band in the gel in SDS‐PAGE assay (Figure 3), indicating that the purified proteins could become antigens for subsequent experiments.
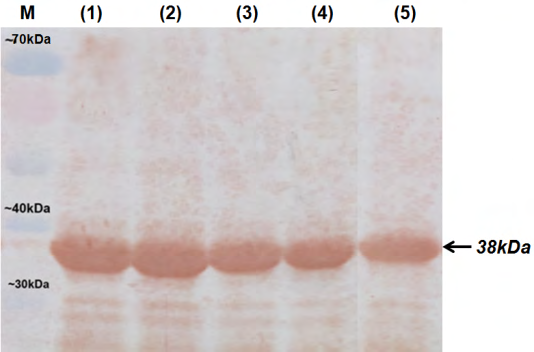
After injected with purified rOmpL1, the white mice were in good-health conditions until day twenty first. A Western blot assay was done with the total protein of five pathogenic Leptospira serovars (L. Canicola, L. Bataviae, L.
Grippotyphosa, L. Icterohaemorrhagiae and L. Pomona). In the assay, only one band at 38 kDa position was shown, thus antibody that collected from serum mice could recognize antigen from all of five Leptospira serovars (Figure 4).
Discussion
The focus of our research is producing a recombinant protein from the OmpL1 gene of pathogenic Leptospira serovars in order to make vaccine against these strains in animals. We have been trying to develop an optimal process to create the protein as our goal. Although combining two or more recombinant proteins in one vaccine has been shown to be effective [12], we found that it was still necessary to evaluate the protective ability of OmpL1 independently for a more comprehensive view about serovars in Vietnam. In this study, we have selected a potential sequence from OmpL1 sequences. The results of sequencing showed that there were 77 differences in sequence between two Leptospira groups in the study and the difference rate of 10%. Therefore, we analyzed the amino acid sequences of both groups carefully by using the 3D construction tool I-TASSER combined with TM-Score tool. The results found that the similarity from amino acid 102 residues to amino acid 316 residues. We have cloned OmpL1 fragment coded for rOmpL1 into pJET1.2 vector and transformed into E.coli DH10b. Normally, the DNA fragments are easily degraded so should be kept in the plasmids because they are more stable. Furthermore, the generation of multiple plasmid copies through culturing E. coli DH10b contained the plasmid is an effective way of gene multiplication to avoid the risk of undesirable mutations in PCR. We have also expressed rOmpL1 in E.coli BL21 expression system. The reasons of choosing these systems are E.coli to be bacteria too, so its protein expressed will be high similar to Leptospira and increase immunological efficiency.
The rOmpL1 was purified by affinity tags since they help collecting large amounts of highly target proteins [14]. In present, there are some widely affinity tag such as maltose- binding protein, elastin-like polypeptide tag, glutathione-S- transferase,... but we used his-tags because of population. Moreover, his-tag does not affect to protein activities. The fact that purified rOmpL1 made immunized mice to produce antibody against OmpL1 protein of Leptospira. Of course, we need to do more in vivo experiments to determine ability of the protein and removing his-tag is necessary for higher amount. Overall, we believe that a description of a process for expression of rOmpL1may be useful for producing a work recombinant vaccine against Leptospira.
Acknowledgement
This research was funded by the VETVACO Join Stock Company, Vietnam.
Conflict of Interest: All authors have no conflict of interest
References
-
Adler B, de la Peña Moctezuma A (2010) Leptospira and leptospirosis. Veterinary microbiology 140(3-4): 287- 296.
-
Haake D, Levett PN (2015) Leptospirosis in humans. Curr Top Microbiol Immunol 387: 65-97.
-
Verma AK, Kumar A, Dhama K, Deb R, Rahal A, et al. (2012) Leptospirosis-persistence of a dilemma: an overview with particular emphasis on trends and recent advances in vaccines and vaccination strategies. Pakistan journal of biological sciences: PJBS 15(20): 954-963.
-
Lindsay KW, Bone I, Fuller G (2010) Neurology and neurosurgery illustrated e-book. In: 5th (Edn.), Elsevier Health Sciences, pp: 612.
-
Mariya R, Chaudhary P, Kumar A, Thangapandian E, Amutha R, et al. (2006) Evaluation of a recombinant LipL41 antigen of Leptospira interrogans serovar Canicola in ELISA for serodiagnosis of bovine leptospirosis. Comparative immunology, microbiology and infectious diseases 29(5-6): 269-277.
-
Bulach DM, Kalambaheti T, de la Peña-Moctezuma A, Adler B (2000) Functional Analysis of Genes in the rfbLocus of Leptospira borgpetersenii Serovar Hardjo Subtype Hardjobovis. Infection and immunity 68(7): 3793-3798.
-
Jorge S, Kremer FS, Oliveira NRd, Navarro GdOSV, Guimarães AM, et al. (2018) Whole-genome sequencing of Leptospira interrogans from southern Brazil: genetic features of a highly virulent strain. Memórias do Instituto Oswaldo Cruz 113(2): 80-86.
-
Dong H, Hu Y, Xue F, Sun D, Ojcius DM, et al. (2008) Characterization of the ompL1 gene of pathogenic Leptospira species in China and cross-immunogenicity of the OmpL1 protein. BMC microbiology 8(1): 223.
-
Flannery B, Costa D, Carvalho FP, Guerreiro H, Matsunaga J, et al. (2001) Evaluation of recombinant Leptospira antigen-based enzyme-linked immunosorbent assays for the serodiagnosis of leptospirosis. Journal of clinical microbiology 39(9): 3303-3310.
-
Haake DA, Champion CI, Martinich C, Shang ES, Blanco DR, et al. (1993) Molecular cloning and sequence analysis of the gene encoding OmpL1, a transmembrane outer membrane protein of pathogenic Leptospira spp. J Bacteriol 175(13): 4225-4234.
-
Barnett JK, Barnett D, Bolin CA, Summers TA, Wagar EA, et al. (1999) Expression and distribution of leptospiral outer membrane components during renal infection of hamsters. Infect Immun 67(2): 853-861.
-
Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, et al. (2012) Primer3—new capabilities and interfaces. Nucleic acids research 40(15): e115-e115.
-
De Jonghe K, Morio S, Maes M (2013) First outbreak of Chrysanthemum stem necrosis virus (CSNV) on potted Chrysanthemum in Belgium. New Disease Reports 28(14): 2044-0588.
-
Arnau J, Lauritzen C, Petersen GE, Pedersen J (2006) Current strategies for the use of affinity tags and tag removal for the purification of recombinant proteins. Protein expression and purification 48(1): 1-13.
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System