Activity Pattern and Time Budget of Common Moorhen (Gallinula Chloropus) at Kumbakonam Region of Tamil Nadu
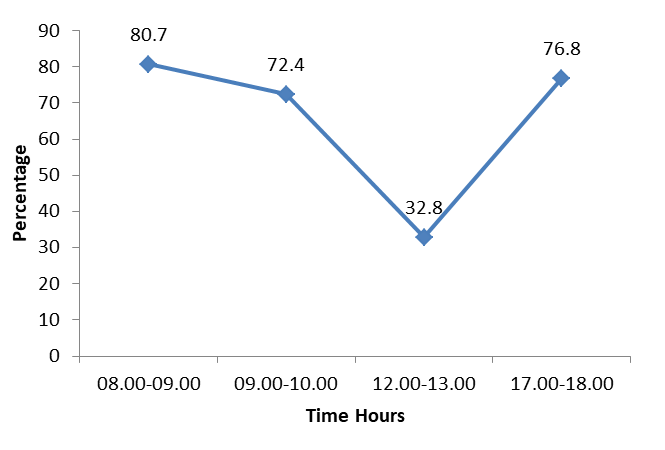
Birds inhabiting wetlands for feeding, breeding, nesting or roosting are broadly defined as water birds. They are one of the essential components of food web and nutrient cycles of the wetland ecosystems. Distribution of waterbirds in a particular wetland mainly depends on availability of food, nesting sites and predation risk. Study on behavior of birds is also believed to be its expressions in response to the internal stimuli mainly related to the physiological needs. Physical displays or posture is one of the several means of animal communications and birds effectively use visual signals. The Common Moorhen Gallinula chloropus is a sexually monomorphic waterbird of the rail family, which exploits a wide spectrum of habitats ranging from rivers and canals to lakes and ponds. This study was carried out in the Cauvery Delta region of Tamil Nadu. Observations on activity of Common moorhen were carried out using focal and scan sampling methods. There were two peaks of feeding in the morning hours; one between 08.00 and 09.00 (80.7%) and another between 09.00 and 10.00 (72.4%). The feeding activity was low in the noon hours and the least was observed between 12.00 and 13.00 (32.8%). Thereafter the feeding gradually increased till 17.00 to 18.00 when the feeding rate reached at its peak (76.8%) in the evening. The overall activities of Common Moorhen shows that the bird spend most of the time 48.96% on Feeding activities followed by 24.54% on Movement, 13.45% on maintenance activities, 5.9% on postures and display, 4.28% on Agonistic and 2.87% on call activities. Overall time spent for different activities by the common Moorhen shows majority of the time they spent feeding activities. In the morning hours they engage movement activities whereas in the noon hours they spend agonistic and maintenance activities.
Introduction
Waterbirds and wetlands are the inseparable and inevitable elements [1]. Birds inhabiting wetlands for feeding, breeding, nesting or roosting are broadly defined as water birds [2]. These waterbirds are essential component of the food web and nutrient cycles of the wetland ecosystems [3]. Abundance of waterbirds in a particular wetland mainly depends on availability of food, nesting sites and predation risk [4]. It is considered as a good bio indicators and useful models of the wetlands for studying the various environmental problems [5]. Wetlands are the main custodians of the water birds [6, 7]. Due to high nutritional value and productivity, it attracts the huge number of migratory and resident bird species [8]. Among the various habitats, wetlands are considered as one of the most threatened ecosystem in the world [9]. During the last century the world has lost over 50% of wetlands due to various human influences, and the remaining of them have been declined because of various human activities [10]. Wetlands in India face tremendous anthropogenic pressure mainly due to discharging of domestic sewage, discharging of industrial effluent, dumping of solid waste, over exploitation of their natural resources and conversion of wetlands in to barren lands. This resulted in biodiversity loss and disturbance of the wetland services [11]. A study has shown about 38% loss of inland wetlands in India during 1971 to 2001. This loss of wetlands has dangerously reduced the availability stop over sites for migrating birds and has increased the importance of remaining wetlands to migrants as well as nesting species [12]. Changes in the habitat condition may affect the relative abundance of bird species composition [13]. However, studying of waterbirds in a wetland are excellent indicators of water quality and measures of biodiversity.
The study on behavior of birds (ethology) has assumed a great significance since each species has an impressive repertoire of innate behaviours and its learning ability is comparable to most mammals and many of the basic behaviors are applicable to human life. According to Pettingill OS, et al. [14] the behaviour is what a bird does and how it responds to its surroundings. Behaviour is also believed to be consisting of the expressions of a bird in response to the internal stimuli mainly related to the physiological needs. Development of effective conservation strategies for a species requires a thorough understanding of its behavior [15]. Physical displays or posture is one of the several means of animal communications and birds effectively use visual signals [16, 17]. In birds visual signals are communicated by the movements of the head, body, tail, wings and body feathers. Many signals may act as stimuli to which members of the same species or other species respond instinctively [18]. For correct interpretation of the gross behavior of a species one must first describe and categorise the individual units, thus establishing a behavioural repertoire [19].
The behaviour pattern is not always constant since the activities could change or modify according to the changes in the physiology of the birds and in the spaciotemporal conditions of the surroundings. However, the birds perform at the most opportune time [20]. The study of activity pattern enables us to understand how ecological factors influence the behaviour of the bird [21]. However, the allotment of time for an activity is determined by natural selection to maximize the chance of survival [22, 23]. The time budget and activity patterns of water birds have mainly been studied by Burton, et al. [24] on geese, Eguchi, et al. on waders and Baldesarre, et al. [25] on ducks. In India although several workers have documented behaviour and activity patterns of birds it needs further more to be done in view of the diverse avifauna available in the country. Some of the workers on related species are Ramachandran, et al, [26] on Pheasant tailed Jacana on Coot. Bhupathy, et al. [27]; Ali, et al. [28], Whistler, et al. [29] have reported their observations on behavior and activity patterns in Purple moorhen.
The Study Species
The Common Moorhen Gallinula chloropus is a sexually monomorphic waterbird of the rail family, which exploits a wide spectrum of habitats ranging from rivers and canals to lakes and ponds [30, 31]. The species was previously believed to have a worldwide distribution but is now thought to occupy the Palearctic region and to overlap with distinct species and/or geographical variations at the margins of its range [31, 32]. Studies on activity and time budget of this species are very less and there is no study was conducted in the study area. Hence the present study was conducted to know the activity and time budget of Common Moorhen with objectives of study various activities of the species, assess the time of various activities and recommend the management strategies for the conservation of the species.
Material and Methods
The present study was conducted during the period between November 2019 and February 2020 to know the activity and time budget of Common moorhen using 8 x 10 Nikon Binocular. Food and feeding and other behaviors were investigated using the focal animal sampling observational method of Altman, et al. [32]. The activity patterns of each bird were recorded by using focal and scan sampling methods (record each animal’s behavior at predetermined time for certain period) with 5 minutes intervals [33, 34, 35, 36, 37, 38]. The activities of all the visible individuals were recorded in each scan. The behavior of one individual during the scan was recorded as one observation. Other important behaviors (event or instantaneous behavior e.g. courtship and copulation, mating etc.) were also noted. Activities were recorded as foraging and feeding, moving, resting, calling, preening, chasing, hiding and breeding [37, 38]. When the bird under the observation was moved out of sight another bird was focused. To reduce disturbance to the focal bird close observations were made hiding behind the trees in the periphery of the pond.
Results
Time Budget and Activity Patterns
The analysis shows that out of 12 activities studied, the bird allotted the highest percentage of daily hours for feeding. There were two peaks of feeding in the morning hours; one between 08.00 and 09.00 (80.7%) and another between 09.00 and 10.00 (72.4%). The feeding activity was low in the noon hours and the least was observed between 12.00 and 13.00 (32.8%). Thereafter the feeding gradually increased till 17.00 to 18.00 when the feeding rate reached at its peak (76.8%) in the evening (Figure 1).
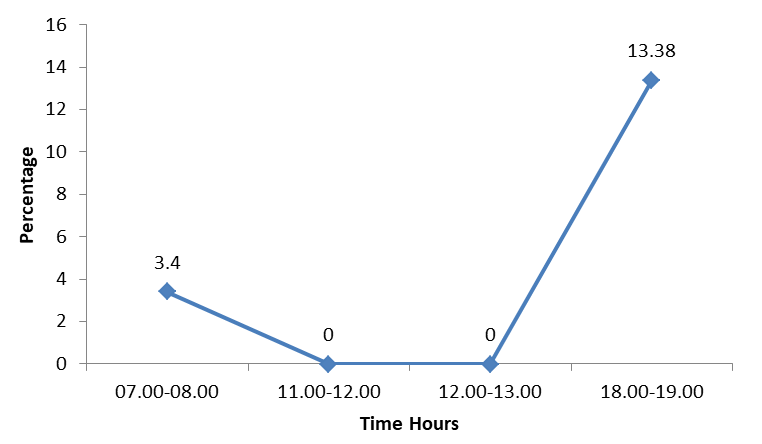
The bird mainly indulged in fighting in the evening hours with a peak of 13.38% between 18.00 and 19.00 (very late evening). Considering the morning hours the bird spent more time (3.4 %) for fighting in the very early morning. Fighting was not observed between 11.00-12.00 and 12.00- 13.00 (Figure 2).
The time budget and pattern of preening was almost similar to bathing. The bird allotted considerable time for preening in the noon hours; 14.3% between 11.00 and 12.00 and 15.87% between 12.00 and 13.00. The bird did not take rest in the very early morning (07.00-08.00). The bird allotted significant time for taking rest in the noon hours and the peak (38.4%) was between 12.00 and 13.00. The bird also spent considerable time for resting (21.9%) between
18.00 and 19.00. The chasing was not observed between 13.00 and 14.00.
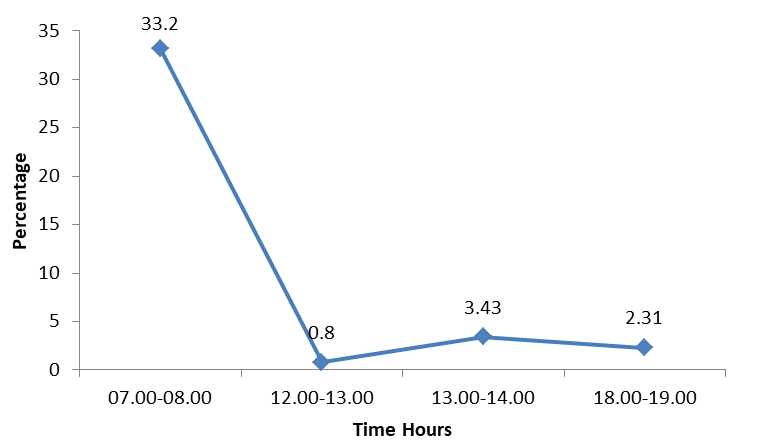
In the early morning (0700-0800) the bird made frequent calls allotting 16.38% time for calling which was the highest daily rate of calling. The bird spent the highest percentage of time (33.2%) for moving in the early morning (07.00-08.00). The bird moved slowly in the noon hours as it spent only 0.8% between 12.00 and 13.00 and 3.43% between 13.00 and 14.00. However the mobility was low in the late evening spending only 2.31% time between 18.00 and 19.00 (Figure 3).
No sexual activity was observed between 07.00 and 09.00, 12.00 and 14.00, 16.00 and 17.00 and 18.00 and 19.00. There was a significant sexual behavior (2.1%) exhibited by the bird between 15.00 and 16.00. It spent highest percentage of time (12.9%) for chasing between 16.00 and 17.00. Bathing was not at all observed or very insignificant in the early morning and evening hours. The bird mainly took bath between 13.00 and 14.00 (13.43%) and between 14.00 and 15.00 (12.21%).
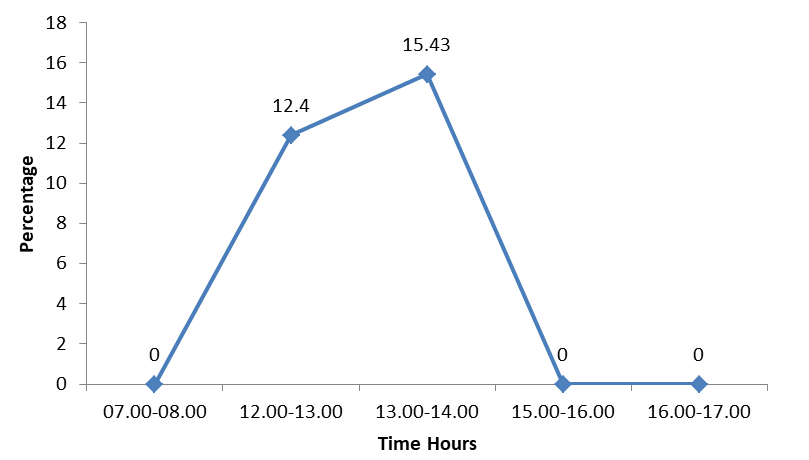
The pattern of swimming almost coincided with that of preening and bathing. The bird was not swimming in the early morning hours between 07.00 and 08.00 and in the evening hours (15.00 - 16.00 and 16.00 -17.00). The bird swam considerably in the noon hours with 12.4% between 12.00 and 13.00 hours and with 15.43% between 13.00 and 14.00 which was the peak rate of swimming (Figure 4).
The hopping was not observed between 07.00 and 08.00, 14.00 and 15.00, 15.00 and 16.00, 16.00 and 17.00 and 18.00 and 19.00 hours. The bird spent the highest time (2.60%) for hopping between 13.00 and 14.00.
Overall Activity Pattern
The overall activities of Common Moorhen shows that the bird spend most of the time 48.96% on Feeding activities followed by 24.54% on Movement, 13.45% on maintenance activities, 5.9% on postures and display, 4.28% on Agonistic and 2.87% on call activities (Figure 5).
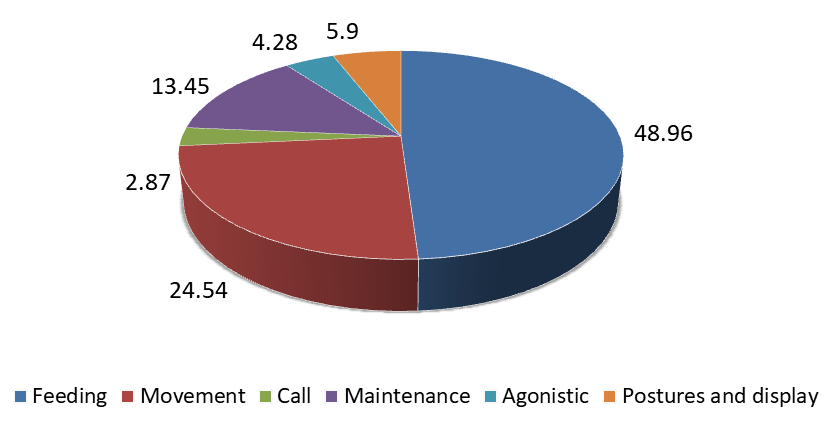
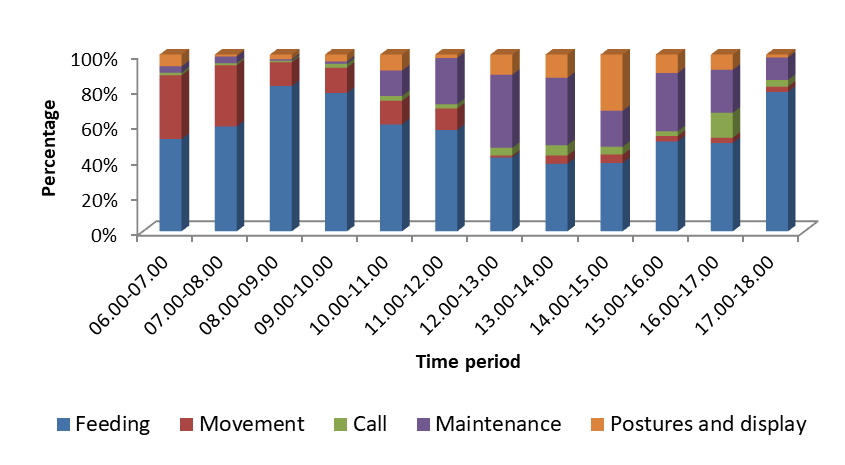
Overall time spent for different activities by the common Moorhen shows majority of the time they spent feeding activities. In the morning hours they engage movement activities whereas in the noon hours they spend Agonistic and maintenance activities (Figure 6).
Discussion
Apparently awakening and roosting of the bird was found coinciding with the time of sunrise and sunset respectively. However in a wider sense these two activities could be tuned with the circadian rhythm of the bird being influenced by various endogenous and exogenous factors like intensity of the light, temperature of the surrounding and the level of the hormones in the blood. In many birds roosting time is strongly influenced by abiotic factors and follows the rules of endogenous light entrained rhythmicity [39, 40]. Temperature, light intensity and external disturbances like human interferences influence the time of awakening and roosting in birds [41]. Swingland, et al. [41] have reported that in the low intensity of light and temperature the birds roost earlier. The birds roost communally helping them to reduce thermoregulation demands and predation risk [43]. Usually a bird selects a roosting site ensuring the safety and protection from predators. This is true in the case of study species also as it selected the middle of the pond habitat for roosting. The middle of the pond has abundant vegetation and high water surrounded by making it inaccessible to the predators. In summer as the most of the habitat dried up the bird roosted on surrounding trees are migrated from the study area which was safer.
As soon as the birds woke up they hurriedly moved to the foraging zone. Therefore in the early morning they were found scattered well throughout the buffer of the pond for feeding actively. However, human and non-human interference increasing in the habitat soon after the late morning forces the birds to move to the center. In the noon most of the birds in the center became less active with feeding becoming slow. In the afternoon again being undisturbed the birds reached the peripheral zone for active feeding but in the evening also interference less from people the birds were forced to stay over to the pheriphery. The human intruders mainly include the students and the non-human intruders were cattle and dogs.
The bird makes varieties of calls [29, 44]. The present study also documents various calls uttered by common moorhen and the functions of some calls are not clear. However there was no report of such anatomical features in common moorhen. Ali, et al. [28] has recognized that the soft calls are meant for maintaining party and rough calls for assembling. The alarm is very loud and contact calls are soft [44].
The male has low sonorous calls and the female has shriller soft calls. Sexual difference of some calls could be seen but most of the calls are uttered similarly by male and female of Porphyrio porphyrio [45]. This supports the observations with the present study that the male and female Gallinula chloropus made certain calls differently. Sexual difference in calls has been reported in some other rails including the Purple moorhen [46], American coot [47]. Similar roles of calls might be attributed to the common Moorhen also since it is a social or communal species. The reduced calls at night would be to ensure better safety in the roost from predators.
Like most of the rails the study species was also a fast runner facilitated by the long spidery spread toes and long legs. The bird although swims well is generally averse to swimming unless often forced to [22]. This agrees with the observation made in the present study. The common moorhen swims little while feeding. Taylor, et al. [48] also reports that the bird swims infrequently and the head moved back and forth while swimming. In the study area the leaves of Nymphaea stellata and Salvinia mat provided suitable substratum for walking, running and skulking. It could move at ease over floating vegetation without getting sunk in water for the body weight was evenly shared by long toes.
Displays, songs and calls are basic mechanism of communication in birds. Common moorhen being a social species the communication through displays are important for the interaction among individuals. Craig, et al. [19] has observed wide ranges of vocal and signal displays in P. melanotus. The roles of various kinds of displays and postures observed in the moorhen were not clearly understood. Head up display is used in fighting in many passerine birds. Head up displays and ruffling of neck feathers were observed in the study species also. The main weapons of aggression in many birds are bill and wings [49]. However the study birds heavy bill and clawed toes were effectively used to attack the opponent. Head up posture is also used by Magpie Robin for defense and fighting [50]. In the head forward posture of common Moorhen showed its red beak and shield towards the opponent. This put the opponent in a great difficulty of pecking until the other bird turned away or faced away. In splattering the bird ran fast towards its opponent. It might serve to show the increasingly aggressive tendency to the opponent. Similar posture was exhibited by common moorhen also.
The Common moorhen was not an exception to the general rule that the herbivorous birds feed actively in the morning and evening. In the early morning prior to leaving for foraging the birds is highly vocal spending considerable time for calling. As soon as they woke up they competed to reach the feeding areas. On the other hand the bird returned rather slowly to the center in the evening. During the study period the bird was very active in the morning. Like many other species the Common moorhen was very inactive in the noon hours. It might be a technique to save energy and to involve in maintenance behavior in the noon hours especially in summer. This could also be a thermoregulatory strategy adopted by the bird. The frequent agonistic behaviors common among the birds in the early morning and evening could be due to the closer assemblage of the birds in the habitat. The frequency of presence of intruders was high in the morning and evening that the birds spent considerable time on vigil during that time. The birds indulged in frequent agonistic encounters in monsoon probably because they became more territorial in the breeding season and also they provided protection to the eggs and chicks from predators. The breeding birds were very active in the monsoon during November as they were busy in courtship displays, mounting, nest construction, brooding and parental caring. Therefore they hardly took rest in the season. However the reason for low time allotment for vigil in monsoon could not be explained satisfactorily because more alertness was expected in breeding season. Probable reason might be that the flooded habitat reduced the access of land predators to the vicinity of the nest. In every daily hour most of the time was set apart for feeding by Jacanas irrespective of the seasons [26]. He has also observed that the time allocated for feeding is inversely proportional to that of maintenance. These findings agree with the behaviour patterns of Common moorhen obtained in the current study. Jayaraman, et al. [50] has also obtained similar pattern in Coot.
Conclusion
Since the study area (the college pond) is within the college campus and also the main entrance, students regularly visiting from 09.00 hrs and stays till 16.00 hrs. Disturbances during this period of time is higher makes the bird always in the center. It is also seen that the food remain from the college canteen is left out in the pond also makes the birds tempt to come in the periphery i.e. in the peak period. It is recommended that steps to be taken to erect a concrete wall along the pond makes less disturbance and prevent from the cattle’s and predators especially stray dogs. Proper waste disposable system may be arranged in the college canteen not to leave any kind of solid wastes especially the wasted food items makes the birds to come near to the canteen site. The Department of Zoology is doing the fresh water pisciculture activities on their routine research work; it is advised to fill the pond with water at least by July so that it attracts frequent visits of water birds early in the seasons.
References
-
Grimmett R, Inskipp T (2005) Birds of Sourthern India. Om Books International, New Delhi, India.
-
Kumar A, Sati JP, Tak P, Alfred JRB (2005) Handbook on Indian Wetland Birds and their Conservations, Zoological Survey of India pp: 472.
-
Dhakate PM, Tejaswini A, Patil, Bhartari R (2008) Wetland Birds of Corbett Tiger Reserve Landscape. Sengupta M, Dalwani R, et al. (Eds.), Proceedings of Taal: The 12th World Lake Conference pp: 1974-1982.
-
Halse SA, Pearson GB, Patrick S (1993) Vegetation of depth-gauged wetland in nature reserves of south- west Western Australia, Department of Conservation and Land Management, Technical Report pp: 30.
-
Mistry J, Mukherjee S (2015) Status and threats of waterbirds in Ahiran lake, Murshidabad, West Bengal, India 5(2).
-
Weller MW (1999) Wetland bird habitat resources and conservation implications. Press syndicate of the University of Cambridge, United Kingdom pp: 316.
-
Stewart RE (2001) Technical Aspects of Wetlands: Wetlands as Bird Habitat. National Water Summary on Wetland Resources, United States Geological Survey pp: 86.
-
Manikannan R, Asokan S, Samsoor M, Ali A (2012) Abundance and Factors Affecting Population Characteristics of Waders (Charadriiformes) in Great Vedaranyam Swamp of Point Calimere Wildlife Sanctuary, South-east Coast of India International Journal of Ecosystem 2(1): 6-14.
-
Prasad SN, Sengupta T, Kumar A, Vijayan VS, Vijayan L, et al. (2015) Wetland of India. 43(1): 173-186.
-
Zhijun MA, Cai Y, Bo Li, Chen J (2010) Managing wetland shabitats for waterbirds: An international perspective, Wetlands 30: 15-27.
-
Ramachandra TV (2006) Soil and Groundwater Pollution from Agricultural Activities, Commonwealth of learning, Canada and Indian Institute of Science, Bangalore, Printed by Capital Publishing Company, New Delhi.
-
Prasad SN, Jaggi AK, Kaushik P, Vijayan L, Muralidharan S, et al. (2004) Inland wetlands of India, Conservation Atlas. Salim Ali Centre for Ornithology and Natural History. Coimbatore, India pp: 222.
-
Nazeema M, Nirmala T (2015) Wetland birds species composition in Tannery tank, Dindigul, Tamilnadu, India. International Research Journal of Environment Sciences 4(5): 34-41.
-
Pettingill OS (1970) Ornithology in laboratory and field. Burgess Publishing Co., Minneapolis, Minnesota, USA.
-
Sutherland WJ (1998) The importance of behavioral studies in conservation biology. Anim Behav 56: 801- 809.
-
Collias NE (1943) Statistical analysis of factors which make for success in initial encounters between hens. American Naturalist 77: 519-538.
-
Butcher GS, Rohwer S (1989) The evolution of conspicuous and distinctive coloration for communication in birds. Curr Ornithol 6: 51-108.
-
Krebs JR, Devies NB (1987) An Introduction to Behavioural Ecology. Blackwell Scientific Publications. London pp: 456.
-
Craig JL (1977) The behaviour of the pukeko Porphyrio porphyrio melanotus. New Zealand journal of zoology 4: 413-433.
-
Smith JM (1976) Evolution and theory of games. Amer Sci 64: 41-45.
-
Boetcher RSM, Haag WC, Bridges JR (1994) Behavioral patterns and nearest neighbor distances among nonbreeding avocets. Condor 96: 973-986.
-
Fagen RM (1974) Selective and evolutionary aspects of animal play. Am Nat 108: 850-588.
-
Macfarland DJ (1977) Decision making in animals. Nature 269: 15-21.
-
Burton BA, Hudson, JR (1978) Activity budgets of Lesser Snow Geese wintering on the Faster River Estuary, British Columbia, Wildfowl 29: 111-l 17.
-
Baldassarre GA, Bolen EG (1994) Waterfowl Ecology and Management. John Wiley and Sons, Inc, New York 49: 342.
-
Ramachandran NK (1998) Activity patterns and time budgets of the Pheasant-tailed (Hydrophaslanus chirurgus) and Bronze winged (Metopidius indicus) Jacanas. Journal of the Bombay Natural History Society pp: 302.
-
Bhupthy S (1985) Ecology of Purple Moorhen (Porphyrio porphyrio) from January to March 1985 in Keoladeo National Park, Bharatpur. M.Sc. Dissertation, Bharathidasan University, Thiruchirapalli, Tamil Nadu, India.
-
Ali S, Ripley S D (1983) Handbook of the birds of India and Pakistan. Oxford University Press, London.
-
Whistler H (1938) Ornithological survey of Jodhpur state. J Bombay Nat Hist Soc 40: 213-235.
-
Cramp S, Simmons KEL (1980) The birds of the western Palearctic. Volume 2. Oxford: Oxford University Press.
-
Chesser RT, Banks RC, Barker FK, Cicero C, Dunn JL, et al. (2011) Fifty-second supplement to the American Ornithologists’ Union Check-list of North American birds. Auk 128: 600-613.
-
Altmann J (1974) Observational study of behavior: Sampling methods. Behaviour 49: 227-267.
-
Feeroz MM, Islam MA (1992) Ecology and behaviour of hoolock gibbons of Bangladesh. MARC (Multidisciplinary Action Research centre), Dhaka, Bangladesh pp: 13.
-
Hasan MK, Feeroz MM, Islam MA, Kabir MM, Begum S, et al. (2005) Food and feeding behaviour of hoolock gibbon (Bunopithecus hoolock hoolock) in a semievergreen forest of Bangladesh. Bangladesh J Life Sci 17(1): 43-49.
-
Akhtar S, Kabir MM, Hasan MK, Begum S (2007) Activity pattern of Pied kingfisher (Ceryle rudis) at Jahangirnagar University Campus. Bangladesh. J Life Sci 19(1): 49-58.
-
Akhtar S, Kabir MM, Hasan MK, Begum S (2009) Activity pattern of Bronze-winged Jacana (Metopidius indicus) at Jahangirnagar University Campus. Bangladesh. J Life Sci 21(2): 111-120.
-
Martin P, Bateson P (1993) Measuring Behaviour: An Introductory guide. In: 2nd (Edn.), Cambridge University Press.
-
Eiserer LA (1984) Communal roosting in birds. Bird Behav 5: 61-80.
-
Gwinner E (1975) Circadian and circannual rhythms in birds. Avian biology. In: Farmer DS, King J. R, Parkes KC, et al. (Eds.), Volume 5, Academic Press, New York, pp: 221-285.
-
Schmitt A (1994) Influence of Abiotic factors on preroosting behavior of Greylag Geese (Anser anser). The Auk 111(3): 759-764.
-
Swingland IR (1976) The influence of light intensity on the roosting time of the rook (Corvus frugilegus). Anim Behav 24: 154-158.
-
Beauchamp G (1999) A comparative study of breeding traits in colonial bird. Evol Ecol Res 1: 251-260.
-
Grimmett R, Inskipp C, Inskipp T (1999) A pocket guide to the birds of the Indian Subcontinent. 1st (Edn.), Delhi: Oxford University Press.
-
Tunnicliffee GA (1965) The biology of the pukeko. Unpubl. M.Sc. thesis, University of Canterbury library.
-
Howard E (1940) A Waterhen’s World. Cambridge University Press, Cambridge.
-
Gullion GW (1950) Voice differences between sexes in the American Coot. Condor 52: 272-272.
-
Tailor B (1998) Rails: A guide to the Rails, Crakes, Gallinules and Coots of the world. United Kingdom, Yale Univ Press, USA.
-
Andrew J (1961) The displays given by passerines in courtship and reproductive fighting: a review Ibis 103: 315-348.
-
Kumar A, Bhatt D (2001) Characteristics and Significance of Calls in Oriental Magpie robin. Current Science 80: 77- 82.
-
Jayaraman S (1985) Wintering ecology of Coots Fulica atra in Keoladeo National Park. Bharatpur, Rajasthan. M.Sc. dissertation, Bharathidasan University, Thiruchirapalli, Tamil Nadu, India. 51. Marler P, Hamilton J (1966) Mechanisms of animal behaviour.John Wiley and Sons, New York, USA, pp: 783.
- Natural Decline to Extinction of A New Zealand Rabbit Population
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System